
The Effect of Diet on Colony Recognition and Cuticular Hydrocarbon Profiles of the Invasive Argentine Ant, Linepithema humile

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dietary Manipulation in the Field and the Laboratory
2.1.1. Field Experiment
2.1.2. Laboratory Experiment
2.2. Behavioral Assays
2.3. Cuticular Hydrocarbon Analysis
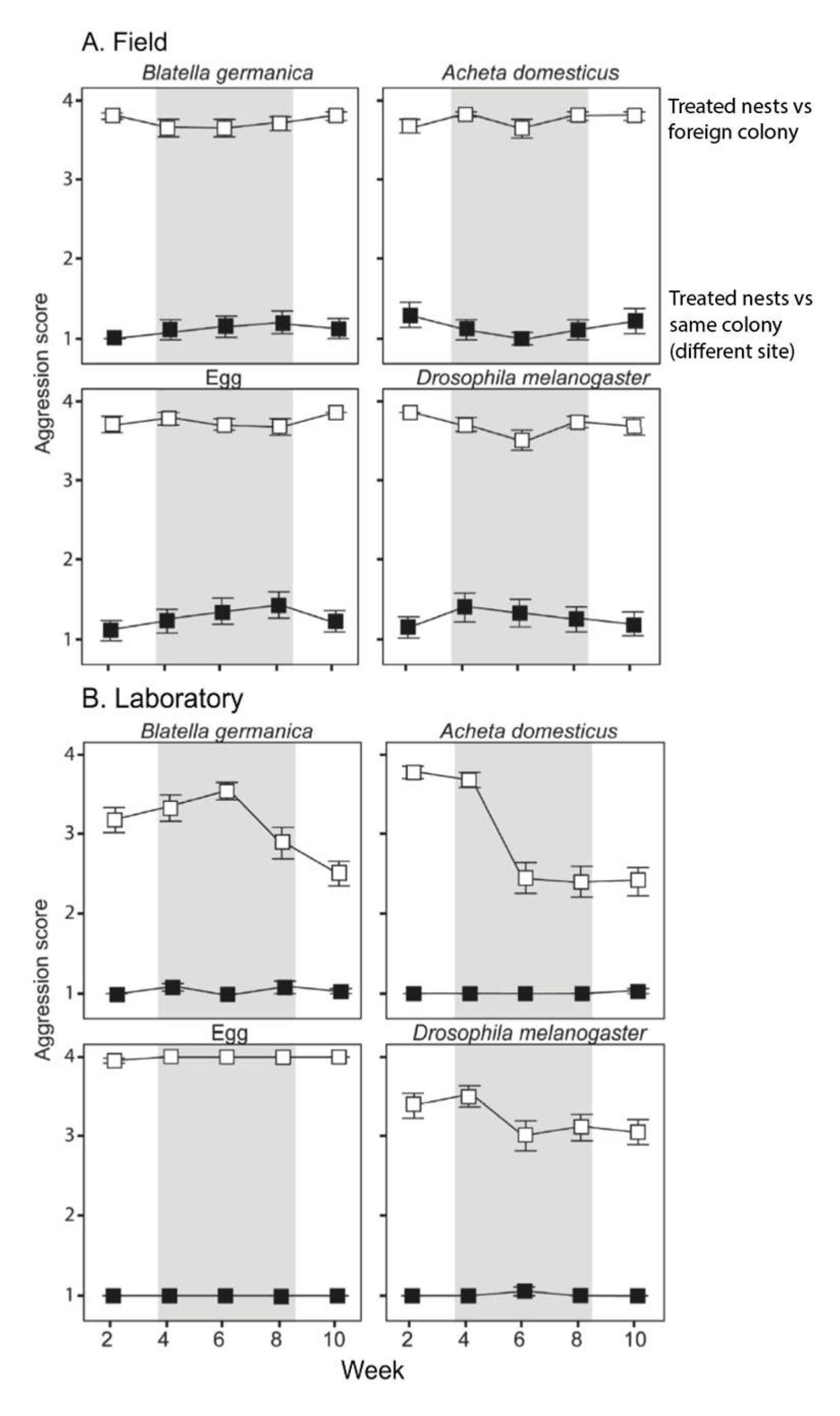
3. Results
Dietary Manipulation in the Field and Laboratory
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blancafort, X.; Gomez, C. Consequences of the Argentine ant, Linepithema humile (Mayr), invasion on pollination of Euphorbia characias (L.) (Euphorbiaceae). Acta Oecologica 2005, 28, 49–55. [Google Scholar] [CrossRef]
- Brandt, M.; van Wilgenburg, E.; Sulc, R.; Shea, K.J.; Tsutsui, N.D. The scent of supercolonies: The discovery, synthesis and behavioural verification of ant colony recognition cues. BMC Biol. 2009, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, M.; van Wilgenburg, E.; Tsutsui, N.D. Global-scale analyses of chemical ecology and population genetics in the invasive Argentine ant. Mol. Ecol. 2009, 18, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Breed, M.D.; Bennett, B. Kin recognition in highly eusocial insects. In Kin Recognition in Animals; Fletcher, D.J.C., Michener, C.D., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 1987; pp. 243–285. [Google Scholar]
- Buczkowski, G.; Kumar, R.; Suib, S.L.; Silverman, J. Diet-related modification of cuticular hydrocarbon profiles of the Argentine ant, Linepithema humile, diminishes intercolony aggression. J. Chem. Ecol. 2005, 31, 829–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, D.A.; Bernier, U.R.; Sutton, B.D. Elution patterns from capillary GC for methyl-branched alkanes. J. Chem. Ecol. 1998, 24, 1845–1865. [Google Scholar] [CrossRef]
- Carney, S.E.; Byerley, M.B.; Holway, D.A. Invasive Argentine ants (Linepithema humile) do not replace native ants as seed dispersers of Dendromecon rigida (Papaveraceae) in California, USA. Oecologia 2003, 135, 576–582. [Google Scholar] [CrossRef]
- Carpintero, S.; Reyes-Lopez, J.; DeReyna, L.A. Impact of Argentine ants (Linepithema humile) on an arboreal ant community in Donana National Park, Spain. Biodivers. Conserv. 2005, 14, 151–163. [Google Scholar] [CrossRef]
- Chapman, R.E.; Bourke, A.F.G. The influence of sociality on the conservation biology of social insects. Ecol. Lett. 2001, 4, 650–662. [Google Scholar] [CrossRef]
- Cole, F.R.; Medeiros, A.C.; Loope, L.L.; Zuehlke, W.W. Effects of the Argentine ant on arthropod fauna of Hawaiian high-elevation shrubland. Ecology 1992, 73, 1313–1322. [Google Scholar] [CrossRef]
- Crozier, R.H.; Dix, M.W. Analysis of two genetic models for the innate components of colony odor in social Hymenoptera. Behav. Ecol. Sociobiol. 1979, 4, 217–224. [Google Scholar] [CrossRef]
- Dittmann, M.; Buczkowski, G.; Scharf, M.; Bennett, G. Gene expression changes in response to field-to-lab transition in the Argentine ant, Linepithema humile. J. Insect Physiol. 2019, 117, 103901. [Google Scholar] [CrossRef] [PubMed]
- Florane, C.B.; Bland, J.M.; Husseneder, C.; Raina, A.K. Diet-mediated inter-colonial aggression in the Formosan subterranean termite Coptotermes formosanus. J. Chem. Ecol. 2004, 30, 2559–2574. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Pedersen, J.S.; Keller, L. Evolution of supercolonies: The Argentine ants of southern Europe. Proc. Natl. Acad. Sci. USA 2002, 99, 6075–6079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinze, J.; Foitzik, S.; Hippert, A.; Hölldobler, B. Apparent dear-enemy phenomenon and environment-based recognition cues in the ant Leptothorax nylanderi. Ethology 1996, 102, 510–522. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Holway, D.A. Effect of Argentine ant invasions on ground-dwelling arthropods in northern California riparian woodlands. Oecologia 1998, 116, 252–258. [Google Scholar] [CrossRef]
- Holway, D.A. Competitive mechanisms underlying the displacement of native ants by the invasive Argentine ant. Ecology 1999, 80, 238–251. [Google Scholar] [CrossRef]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef] [Green Version]
- Holway, D.A.; Suarez, A.V. Colony-structure variation and interspecific competitive ability in the invasive Argentine ant. Oecologia 2004, 138, 216–222. [Google Scholar] [CrossRef]
- Holway, D.A.; Suarez, A.V.; Case, T.J. Loss of intraspecific aggression in the success of a widespread invasive social insect. Science 1998, 283, 949–952. [Google Scholar]
- Howard, R.W. Cuticular hydrocarbons and chemical communication. In Insect Lipids: Chemistry, Biochemistry and Biology; Stanley-Samuelson, D.W., Nelson, D.R., Eds.; University of Nebraska Press: Lincoln, NE, USA, 1993; pp. 179–226. [Google Scholar]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Human, K.G.; Gordon, D.M. Exploitation and interference competition between the invasive Argentine ant, Linepithema humile, and native ant species. Oecologia 1996, 105, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Jaquiery, J.; Vogel, V.; Keller, L. Multilevel genetic analyses of two European supercolonies of the Argentine ant, Linepithema humile. Mol. Ecol. 2005, 14, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Jutsum, A.R. Interspecific aggression in leaf-cutting ants. Anim. Behav. 1979, 27, 833–838. [Google Scholar] [CrossRef]
- Lahav, S.; Soroker, V.; Hefetz, A.; Vander Meer, R.K. Direct behavioral evidence for hydrocarbons as ant recognition discriminators. Naturwissenschaften 1999, 86, 246–249. [Google Scholar] [CrossRef]
- Lenoir, A.; Fresneau, D.; Errard, C.; Hefetz, A. Individuality and colonial identity in ants: The emergence of the social representation concept. In Information Processing in Social Insects; Detrain, C., Deneubourg, J.-L., Pasteels, J.-M., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1999. [Google Scholar]
- Liang, D. Hydrocarbon-released nestmate aggression in the Argentine ant, Linepithema humile, following encounters with insect prey. Comp. Biochem. Physiol. Part B 2001, 129, 871–882. [Google Scholar] [CrossRef]
- Liang, D.; Silverman, J. “You are what you eat”: Diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepithema humile. Naturwissenschaften 2000, 87, 412–416. [Google Scholar] [CrossRef]
- Matsuura, K. Nestmate recognition mediated by intestinal bacteria in a termite, Reticulitermes speratus. Oikos 2001, 92, 20–26. [Google Scholar] [CrossRef]
- Moffett, M.W. The Human Swarm: How Our Societies Arise, Thrive, and Fall; Basic Books: New York, NY, USA, 2019. [Google Scholar]
- Mothapo, N.P.; Wossler, T.C. “You are not always what you eat”: Diet did not override intrinsic nestmate recognition cues in Argentine ants from two supercolonies in South Africa. Afr. Zool. 2016, 51, 161–171. [Google Scholar] [CrossRef]
- Naughton, I.; Boser, C.; Tsutsui, N.D.; Holway, D.A. Direct evidence of native ant displacement by the Argentine ant in island ecosystems. Biol. Invasions 2020, 22, 681–691. [Google Scholar] [CrossRef]
- Obin, M.S.; Vandermeer, R.K. Sources of nestmate recognition cues in the imported fire ant Solenopsis invicta Buren (Hymenoptera, Formicidae). Anim. Behav. 1988, 36, 1361–1370. [Google Scholar] [CrossRef]
- Pedersen, J.S.; Krieger, M.J.B.; Vogel, V.; Giraud, T.; Keller, L. Native supercolonies of unrelated individuals in the invasive Argentine ant. Evolution 2006, 60, 782–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, F.J.; Hefetz, A.; Christides, J.P.; Errard, C. Food influence on colonial recognition and chemical signature between nestmates in the fungus- growing ant Acromyrmex subterraneus subterraneus. Chemoecology 2004, 14, 9–16. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S., 3rd; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Sanders, N.J.; Gotelli, N.J.; Heller, N.E.; Gordon, D.M. Community disassembly by an invasive species. Proc. Natl. Acad. Sci. USA 2003, 100, 2474–2477. [Google Scholar] [CrossRef] [Green Version]
- Suarez, A.V.; Bolger, D.T.; Case, T.J. Effects of fragmentation and invasion on native ant communities in coastal southern California. Ecology 1998, 79, 2041–2056. [Google Scholar] [CrossRef]
- Suarez, A.V.; Case, T.J. Bottom-up effects on persistence of a specialist predator: Ant invasions and horned lizards. Ecol. Appl. 2002, 12, 291–298. [Google Scholar] [CrossRef]
- Suarez, A.V.; Holway, D.A.; Liang, D.; Tsutsui, N.D.; Case, T.J. Spatiotemporal patterns of intraspecific aggression in the invasive Argentine ant. Anim. Behav. 2002, 64, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Suarez, A.V.; Tsutsui, N.D.; Holway, D.A.; Case, T.J. Behavioral and genetic differentiation between native and introduced populations of the Argentine ant. Biol. Invasions 1999, 1, 43–53. [Google Scholar] [CrossRef]
- Thomas, M.L.; Payne-Makrisa, C.M.; Suarez, A.V.; Tsutsui, N.D.; Holway, D.A. When supercolonies collide: Territorial aggression in an invasive and unicolonial social insect. Mol. Ecol. 2006, 15, 4303–4315. [Google Scholar] [CrossRef]
- Torres, C.W.; Brandt, M.; Tsutsui, N.D. The role of cuticular hydrocarbons as chemical cues for nestmate recognition in the invasive Argentine ant (Linepithema humile). Insectes Sociaux 2007, 54, 363–373. [Google Scholar] [CrossRef]
- Tsutsui, N.D.; Case, T.J. Population genetics and colony structure of the Argentine ant (Linepithema humile) in its native and introduced ranges. Evolution 2001, 55, 976–985. [Google Scholar] [CrossRef]
- Tsutsui, N.D.; Suarez, A.V. The colony structure and population biology of invasive ants. Conserv. Biol. 2003, 17, 48–58. [Google Scholar] [CrossRef]
- Tsutsui, N.D.; Suarez, A.V.; Grosberg, R.K. Genetic diversity, asymmetrical aggression, and recognition in a widespread invasive species. Proc. Natl. Acad. Sci. USA 2003, 100, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsutsui, N.D.; Suarez, A.V.; Holway, D.A.; Case, T.J. Reduced genetic variation and the success of an invasive species. Proc. Natl. Acad. Sci. USA 2000, 97, 5948–5953. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, N.D.; Suarez, A.V.; Holway, D.A.; Case, T.J. Relationships among native and introduced populations of the Argentine ant (Linepithema humile) and the source of introduced populations. Mol. Ecol. 2001, 10, 2151–2161. [Google Scholar] [CrossRef]
- van Wilgenburg, E.; Clemencet, J.; Tsutsui, N.D. Experience influences aggressive behaviour in the Argentine ant. Biol. Lett. 2010, 6, 152–155. [Google Scholar] [CrossRef] [Green Version]
- van Wilgenburg, E.; Sulc, R.; Shea, K.J.; Tsutsui, N.D. Deciphering the chemical basis of nestmate recognition. J. Chem. Ecol. 2010, 36, 751–758. [Google Scholar] [CrossRef] [Green Version]
- van Wilgenburg, E.; Torres, C.W.; Tsutsui, N.D. The global expansion of a single ant supercolony. Evol. Appl. 2010, 3, 136–143. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Dantonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Walters, A.C. Invasion of Argentine ants (Hymenoptera:Formicidae) in South Australia: Impacts on community composition and abundance of invertebrates in urban parklands. Aust. Ecol. 2006, 31, 567–576. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Before | During | After | Before vs. During | During vs. After | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Opponent | Treatment | Mean ± SD | n | n | n | Z | p | Z | p- | ||
| Same supercolony | A. domesticus | 1.43 ± 0.53 | 8 | 1.23 ± 0.45 | 7 | 1.34 ± 0.53 | 7 | −0.853 | 0.393 | −0.597 | 0.551 |
| egg | 1.22 ± 0.31 | 8 | 1.55 ± 0.63 | 8 | 1.35 ± 0.40 | 8 | −1.069 | 0.285 | −0.541 | 0.588 | |
| D. melanogaster | 1.28 ± 0.44 | 8 | 1.37 ± 0.53 | 7 | 1.12 ± 0.30 | 7 | −0.393 | 0.694 | −0.07 | 0.944 | |
| B. germanica | 1.15 ± 0.23 | 8 | 1.33 ± 0.44 | 8 | 1.25 ± 0.30 | 8 | −0.756 | 0.45 | −0.171 | 0.864 | |
| Different supercolony | A. domesticus | 3.83 ± 0.20 | 8 | 3.94 ± 0.15 | 7 | 3.94 ± 0.15 | 7 | −1.326 | 0.185 | 0 | 1 |
| egg | 3.85 ± 0.28 | 8 | 3.83 ± 0.27 | 8 | 4.00 ± 0.00 | 8 | −0.387 | 0.669 | −1.852 | 0.064 | |
| D. melanogaster | 4.00 ± 0.00 | 8 | 3.89 ± 0.23 | 8 | 3.83 ± 0.29 | 7 | 1.565 | 0.118 | −0.161 | 0.876 | |
| B. germanica | 3.95 ± 0.09 | 8 | 3.85 ± 0.23 | 8 | 3.95 ± 0.14 | 8 | −0.769 | 0.442 | −1.105 | 0.269 | |
| Before | During | After | Before versus During | During versus After | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Opponent | Treatment | Mean ± SD | n | n | n | Z | p- | Z | p | ||
| Same supercolony | A. domesticus | 1.00 ± 0.00 | 8 | 1.00 ± 0.00 | 8 | 1.25 ± 0.07 | 8 | −1 | 0.317 | −1 | 0.317 |
| egg | 1.00 ± 0.00 | 8 | 1.00 ± 0.00 | 8 | 1.00 ± 0.00 | 8 | 0 | 1 | 0 | 1 | |
| D. melanogaster | 1.00 ± 0.00 | 8 | 1.00 ± 0.00 | 8 | 1.00 ± 0.00 | 8 | 0 | 1 | 0 | 1 | |
| B. germanica | 1.00 ± 0.00 | 8 | 1.08 ± 0.21 | 8 | 1.03 ± 0.07 | 8 | 0 | 1 | −0.091 | 0.927 | |
| Different supercolony | A. domesticus | 3.38 ± 0.36 | 8 | 3.10 ± 0.19 | 8 | 3.05 ± 0.26 | 8 | −3.401 | 0.001 | 0 | 1 |
| egg | 3.78 ± 0.20 | 8 | 2.40 ± 0.44 | 8 | 2.40 ± 0.37 | 8 | 1.464 | 0.143 | 0 | 1 | |
| D. melanogaster | 3.95 ± 0.09 | 8 | 4.00 ± 0.00 | 8 | 4.00 ± 0.00 | 8 | −1.846 | 0.083 | −0.113 | 0.91 | |
| B. germanica | 3.18 ± 0.40 | 8 | 2.88 ± 0.54 | 8 | 2.50 ± 0.32 | 8 | −1.124 | 0.261 | −1.807 | 0.071 | |
| Laboratory | Field | |||||||
|---|---|---|---|---|---|---|---|---|
| Cuticular | Before | During | Before | During | ||||
| Hydrocarbon | Mean ± SD | Mean ± SD | Z-Value | p-Value | Mean ± SD | Mean ± SD | Z-Value | p-Value |
| 1 | 0.000 ± 0.000 | 0.005 ± 0.005 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 2 | 0.000 ± 0.000 | 0.003 ± 0.002 | −3.59 | <0.001 | 0.002 ± 0.002 | 0.002 ± 0.001 | 0 | 1 |
| 3 | 0.005 ± 0.001 | 0.009 ± 0.005 | −2.406 | 0.016 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 4 | 0.000 ± 0.000 | 0.007 ± 0.005 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 5 | 0.000 ± 0.000 | 0.006 ± 0.004 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 6 | 0.106 ± 0.023 | 0.103 ± 0.048 | −0.21 | 0.834 | 0.055 ± 0.024 | 0.053 ± 0.026 | −0.105 | 0.916 |
| 7 | 0.000 ± 0.000 | 0.046 ± 0.029 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 9 | 0.000 ± 0.000 | 0.018 ± 0.018 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | −0.316 | 0.752 |
| 9 | 0.000 ± 0.000 | 0.037 ± 0.021 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 10 | 0.010 ± 0.004 | 0.058 ± 0.044 | −3.361 | 0.001 | 0.001 ± 0.002 | 0.001 ± 0.002 | −0.231 | 0.817 |
| 11 | 0.000 ± 0.000 | 0.041 ± 0.020 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 12. | 0.000 ± 0.000 | 0.106 ± 0.061 | −3.59 | <0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| Laboratory | Field | |||||||
|---|---|---|---|---|---|---|---|---|
| Cuticular | Before | During | Before | During | ||||
| Hydrocarbon | Mean ± SD | Mean ± SD | Z-Value | p-Value | Mean ± SD | Mean ± SD | Z-Value | p-Value |
| 1 | 0.273 ± 0.148 | 0.192 ± 0.087 | −0.926 | 0.355 | 0.130 ± 0.175 | 0.114 ± 0.087 | −0.575 | 0.565 |
| 2 | 0.030 ± 0.016 | 0.019 ± 0.010 | −1.389 | 0.165 | 0.016 ± 0.028 | 0.014 ± 0.007 | −1.023 | 0.306 |
| 3 | 0.015 ± 0.041 | 0.000 ± 0.000 | −0.935 | 0.35 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 4 | 0.178 ± 0.151 | 0.120 ± 0.041 | −0.694 | 0.487 | 0.016 ± 0.034 | 0.000 ± 0.000 | −1.468 | 0.142 |
| 5 | 0.000 ± 0.000 | 0.007 ± 0.013 | −1.565 | 0.118 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 6 | 0.000 ± 0.000 | 0.004 ± 0.008 | −1.985 | 0.047 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 7 | 0.000 ± 0.000 | 0.001 ± 0.002 | −1.985 | 0.047 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 8 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
| 9 | 0.001 ± 0.002 | 0.008 ± 0.004 | −3.227 | 0.001 | 0.000 ± 0.001 | 0.000 ± 0.000 | −1 | 0.317 |
| 10 | 0.000 ± 0.000 | 0.032 ± 0.018 | −3.133 | 0.002 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Wilgenburg, E.; Mariotta, M.; Tsutsui, N.D. The Effect of Diet on Colony Recognition and Cuticular Hydrocarbon Profiles of the Invasive Argentine Ant, Linepithema humile. Insects 2022, 13, 335. https://doi.org/10.3390/insects13040335
van Wilgenburg E, Mariotta M, Tsutsui ND. The Effect of Diet on Colony Recognition and Cuticular Hydrocarbon Profiles of the Invasive Argentine Ant, Linepithema humile. Insects. 2022; 13(4):335. https://doi.org/10.3390/insects13040335
Chicago/Turabian Stylevan Wilgenburg, Ellen, Mario Mariotta, and Neil Durie Tsutsui. 2022. "The Effect of Diet on Colony Recognition and Cuticular Hydrocarbon Profiles of the Invasive Argentine Ant, Linepithema humile" Insects 13, no. 4: 335. https://doi.org/10.3390/insects13040335
APA Stylevan Wilgenburg, E., Mariotta, M., & Tsutsui, N. D. (2022). The Effect of Diet on Colony Recognition and Cuticular Hydrocarbon Profiles of the Invasive Argentine Ant, Linepithema humile. Insects, 13(4), 335. https://doi.org/10.3390/insects13040335