
Silencing of Chitin-Binding Protein with PYPV-Rich Domain Impairs Cuticle and Wing Development in the Asian Citrus Psyllid, Diaphorina citri


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. D. citri Rearing and Sample Collection
2.2. RNA Extraction, cDNA Synthesis and RT-qPCR Analysis
2.3. cDNA Library Preparation and Illumina Sequencing
2.4. Read Mapping, DEGs Identification and Functional Annotation
2.5. D. citri Total Proteins Isolation
2.6. Chitin and Cellulose Binding Properties Analysis
2.7. In-Gel Digestion and LC-MS/MS Analysis
2.8. Protein Identification and Bioinformatics Analysis
2.9. RNAi-Mediadted DcCP64 Silencing
2.10. Eosin Y Staining and Scanning Election Microscopy (SEM) Analysis
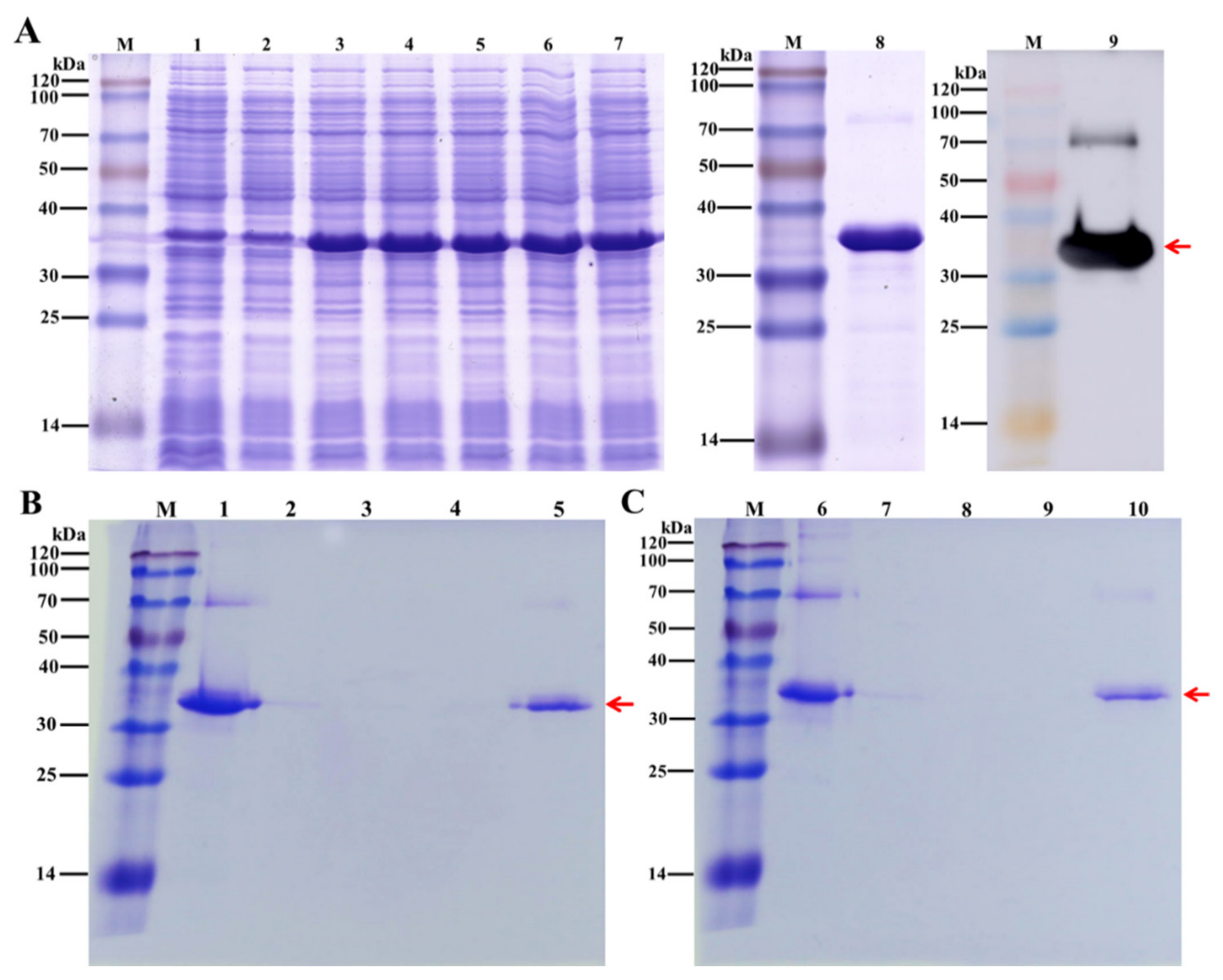
2.11. Protein Expression and Purification
3. Results
3.1. Identification of the DcCP64 Gene and Bioinformatics Analysis
3.2. Expression Patterns of DcCP64 in Different Tissues and Developmental Stages
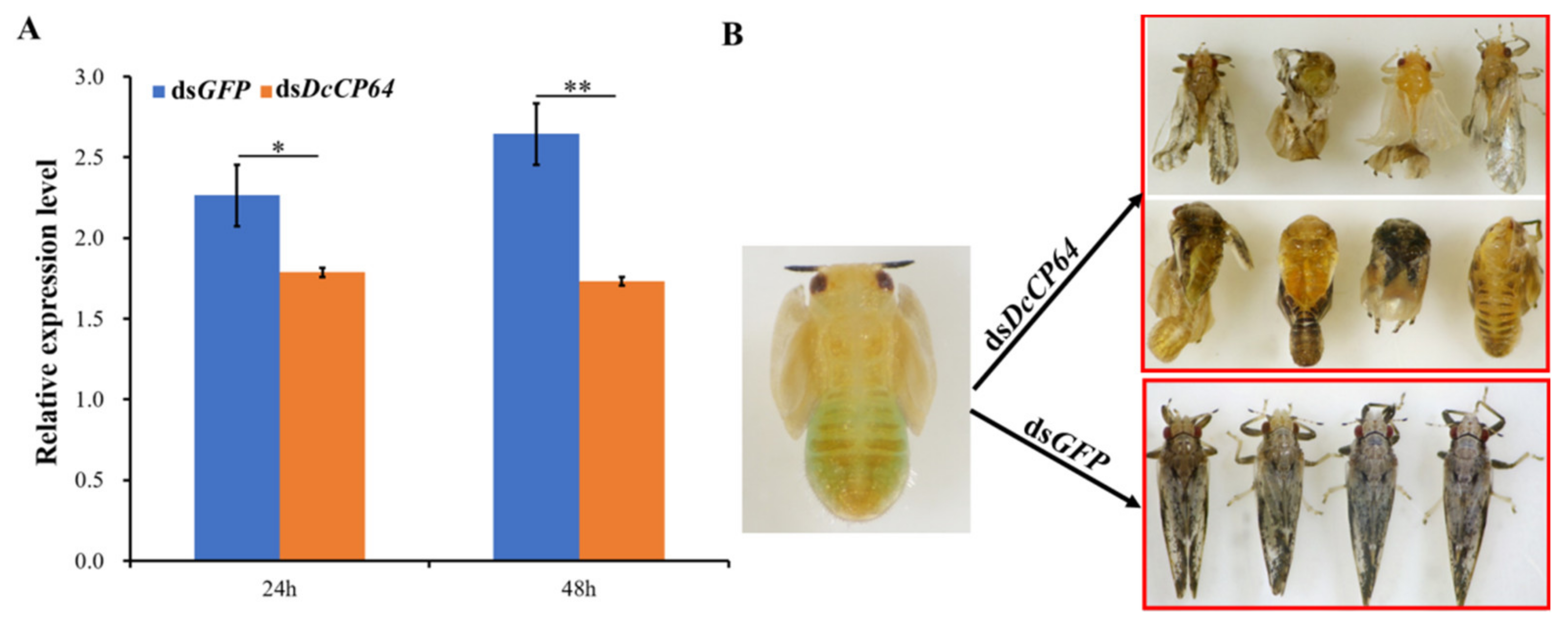
3.3. RNAi-Based Silencing of DcCP64 and Phenotypic Analysis
3.4. Permeability and Microstructure Analysis after Silencing of DcCP64
3.5. Verify the Chitin-Binding Using Recombinant DcCP64 Protein
3.6. Transcriptome Sequence and Assembly
3.7. Identification of DEGs and Functional Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleites, L.A.; Johnson, R.; Kruse, A.R.; Nachman, R.J.; Hall, D.G.; MacCoss, M.; Heck, M.L. Peptidomics approaches for the identification of bioactive molecules from Diaphorina citri. J. Proteome Res. 2020, 19, 1392–1408. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.Z.; Li, J.Y.; Rangel, L.T. The Candidatus liberibacter-host interface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, T.F.S.; Schneider, V.K.; Kishi, L.T.; Carmona, A.K.; Alves, M.F.M.; Belasque-Junior, J.; Rosa, J.C.; Hunter, W.B.; Henrique-Silva, F.; Soares-Costa, A. Characterization of a recombinant cathepsin B-like cysteine peptidase from Diaphorina citri. PLoS ONE 2015, 10, e0145132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, S.; Clayson, P.J.; Kuhns, E.H.; Stelinski, L.L. Effects of buprofezin and diflubenzuron on various developmental stages of Asian citrus psyllid, Diaphorina citri. Pest. Manag. Sci. 2012, 68, 1405–1412. [Google Scholar] [CrossRef]
- Huang, J.K.; Hu, R.F.; Pray, C.; Qiao, F.B.; Rozelle, S. Biotechnology as an alternative to chemical pesticides: A case study of Bt cotton in China. Agr. Econ. 2003, 29, 55–67. [Google Scholar] [CrossRef]
- Klepal, W.; Barnes, H. An ultrastructural study of the formation of the exoskeleton of stage I and II cirripede nauplii during embryonic development. J. Exp. Mar. Biol. Ecol. 1978, 32, 241–257. [Google Scholar] [CrossRef]
- Moussian, B. Recent advances in understanding mechanisms of insect cuticle differentiation. Insect Biochem. Mol. Biol. 2010, 40, 363–375. [Google Scholar] [CrossRef]
- Cornman, R.S.; Willis, J.H. Annotation and analysis of low-complexity protein families of Anopheles gambiae that are associated with cuticle. Insect Mol. Biol. 2009, 18, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fan, H.W.; Huang, H.J.; Xue, J.; Wu, W.J.; Bao, Y.Y.; Xu, H.J.; Zhu, Z.R.; Cheng, J.A.; Zhang, C.X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvata lugens and Laodelphax striatellus. Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Li, Z.L.; Tian, S.; Yang, H.; Zhou, X.; Xu, S.P.; Zhang, Z.Y.; Gong, J.; Hou, Y.; Xia, Q.Y. Genome-wide identification of chi tin-binding proteins and characterization of BmCBP1 in the silkworm, Bombyx mori. Insect Sci. 2019, 26, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Park, M.J.; Kim, B.Y.; Jin, B.R. Molecular characterization of a chitin-binding protein with the peritrophin-A domain from the Asiatic honeybee Apis cerana. J. Asia Pac. Entomol. 2016, 19, 963–968. [Google Scholar] [CrossRef]
- Deng, H.M.; Li, Y.; Zhang, J.L.; Liu, L.; Feng, Q.L. Analysis of expression and chitin-binding activity of the wing disc cuticle protein BmWCP4 in the silkworm, Bombyx mori. Insect Sci. 2016, 23, 782–790. [Google Scholar] [CrossRef]
- Futahashi, R.; Okamoto, S.; Kawasaki, H.; Zhong, Y.S.; Iwanaga, M.; Mita, K.; Fujiwara, H. Genome-wide identification of cuticular protein genes in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1138–1146. [Google Scholar] [CrossRef]
- Liu, J.Q.; Li, S.L.; Li, W.S.; Peng, L.; Chen, Z.W.; Xiao, Y.D.; Guo, H.Z.; Zhang, J.W.; Cheng, T.C.; Goldsmith, M.R.; et al. Genome-wide annotation and comparative analysis of cuticular protein genes in the noctuid pest Spodoptera litura. Insect Biochem. Mol. Biol. 2019, 110, 90–97. [Google Scholar] [CrossRef]
- Togawa, T.; Dunn, W.A.; Emmons, A.C.; Nagao, J.; Willis, J.H. Developmental expression patterns of cuticular protein genes with the R&R consensus from Anopheles gambiae. Insect Biochem. Mol. Biol. 2008, 38, 508–519. [Google Scholar]
- Pan, P.L.; Ye, Y.X.; Lou, Y.H.; Lu, J.B.; Chen, C.; Shen, Y.; Moussian, B.; Zhang, C.X. A comprehensive omics analysis and functional survery of cuticular proteins in the brown planthopper. Proc. Natl. Acad. USA 2018, 115, 5175–5180. [Google Scholar] [CrossRef] [Green Version]
- Song, T.Q.; Yang, M.L.; Wang, Y.L.; Liu, Q.; Wang, H.M.; Zhang, J.; Li, T. Cuticular protein LmTwd1 is involved in molt development of the migratory locust. Insect Sci. 2016, 23, 520–530. [Google Scholar] [CrossRef]
- Dittmer, N.T.; Tetreau, G.; Cao, X.L.; Jiang, H.B.; Wang, P.; Kanost, M.R. Annotation and expression analysis of cuticular proteins from the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2015, 62, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef]
- Copper, A.M.; Silver, K.; Zhang, J.Z.; Park, Y.; Zhu, K.Y. Molecular mechanisms influencing efficiency of RNA interference in insects. Pest Manag. Sci. 2019, 75, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.Y.; Jing, T.X.; Li, W.; Liu, X.Q.; Liu, T.Y.; Liu, Y.; Chen, M.L.; Jiang, R.X.; Yuan, G.R.; Dou, W.; et al. NADPH-cytochrome P450 reductase mediates the susceptibility of Asian citrus psyllid Diaphorina citri to imidacloprid and thiamethoxam. Pest Manag. Sci. 2021, 77, 677–685. [Google Scholar] [CrossRef]
- Yu, X.D.; Killiny, N. RNA interference of two glutathione S-transferase genes, Diaphorina citri DcGSTe2 and DcGSTd1, increases the susceptibility of Asian citrus psyllid (Hemiptera: Liviidae) to the pesticides, fenpropathrin and thiamethoxam. Pest Manag. Sci. 2018, 74, 638–647. [Google Scholar] [CrossRef]
- Yu, H.Z.; Huang, Y.L.; Lu, Z.J.; Zhang, Q.; Su, H.N.; Du, Y.M.; Yi, L.; Zhong, B.L.; Chen, C.X. Inhibition of trehalase affects the trehalose and chitin metabolism pathways in Diaphorina citri (Hemiptera: Psyllidae). Insect Sci. 2021, 28, 718–734. [Google Scholar] [CrossRef]
- Lu, Z.J.; Zhou, C.H.; Yu, H.Z.; Huang, Y.L.; Liu, Y.X.; Xie, Y.X.; Wang, J.; Hu, W.; Su, H.N.; Yang, C. Potential roles of insect Tropomyosin1-X1 isoform in the process of Candidatus Liberibacter asiaticus infection of Diaphorina citri. J. Insect Physiol. 2019, 114, 125–135. [Google Scholar] [CrossRef]
- Tang, L.; Liang, J.B.; Zhan, Z.G.; Xiang, Z.H.; He, N.J. Identification of the chitin-binding proteins from the larval proteins of silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 228–234. [Google Scholar] [CrossRef]
- Yu, H.Z.; Xie, Y.X.; Wang, J.; Wang, Y.; Du, Y.M.; Wang, H.G.; Zhong, B.L.; Zhu, B.; Yu, X.D.; Lu, Z.J. Integrated transcriptome sequencing and RNA interference reveals molecular changes in Diaphorina citri after exposure to validamycin. Insect Sci. 2021, 28, 1690–1707. [Google Scholar] [CrossRef]
- Dong, W.; Gao, Y.H.; Zhang, X.B.; Moussian, B.; Zhang, J.Z. Chitinase 10 controls chitin amounts and organization in the wing cuticle of Drosophila. Insect Sci. 2020, 27, 1198–1207. [Google Scholar] [CrossRef]
- Li, L.; Jiang, Y.P.; Liu, Z.Y.; You, L.L.; Wu, Y.; Xu, B.; Ge, L.Q.; Stanley, D.; Song, Q.S.; Wu, J.C. Jinggangmycin increases fecundity of the brown planthopper, Nilaparvata lugens (Stål) via fatty acid synthase gene expression. J. Proteomics 2016, 130, 140–149. [Google Scholar] [CrossRef]
- Dong, Z.M.; Zhang, W.W.; Zhang, Y.; Zhang, X.L.; Zhao, P.; Xia, Q.Y. Identification and characterization of novel chi tin-binding proteins from the larval cuticle of silkworm, Bombyx mori. J. Proteome Res. 2016, 15, 1435–1445. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juarez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puinean, A.M.; Foster, S.P.; Oliphantm, L.; Denholm, I.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Chris, B. Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebers, J.E.; Wills, J.H. A conserved domain in arthropod cuticular proteins binds chitin. Insect Biochem. Mol. Biol. 2001, 31, 1083–1093. [Google Scholar] [CrossRef]
- Lu, J.B.; Luo, X.M.; Zhang, X.Y.; Pan, P.L.; Zhang, C.X. An ungrouped cuticular protein is essential for normal endocuticle formation in the brown planthopper. Insect Biochem. Mol. Biol. 2018, 100, 1–9. [Google Scholar] [CrossRef]
- Zhao, X.M.; Gou, X.; Liu, W.M.; Ma, E.B.; Moussian, B.; Li, S.; Zhu, K.Y.; Zhang, J.Z. The wing-specific cuticular protein LmACP7 is essential for normal wing morphogenesis in the migratory locust. Insect Biochem. Mol. Biol. 2019, 112, 103206. [Google Scholar] [CrossRef]
- Zhang, W.W.; Dong, Z.M.; Zhang, Y.; Zhang, X.L.; Zhang, S.Y.; Zhao, P. Expression pattern and chitin-binding mode analyses of cuticle protein BmCPAP3-G in the silkworm (Bombyx mori). Sci. Agri. Sin. 2017, 50, 1723–1733. [Google Scholar]
- Boina, D.R.; Rogers, M.E.; Wang, N.; Stelinski, L.L. Effect of pyriproxyfen, a juvenile hormone mimic, on egg hatch, nymph development, adult emergence and reproduction of the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2010, 66, 349–357. [Google Scholar] [CrossRef]
- Charles, J.P. The regulation of expression of insect cuticle protein genes. Insect Biochem. Mol. Biol. 2010, 40, 205–213. [Google Scholar] [CrossRef]
- Shang, F.; Ding, B.Y.; Ye, C.; Yang, L.; Chang, T.Y.; Xie, J.Q.; Tang, L.D.; Niu, J.Z.; Wang, J.J. Evaluation of a cuticle protein gene as a potential RNAi target in aphids. Pest Manag. Sci. 2020, 76, 134–140. [Google Scholar] [CrossRef]
- Lu, Z.J.; Huang, Y.L.; Yu, H.Z.; Li, N.Y.; Xie, Y.X.; Zhang, Q.; Zeng, X.D.; Hu, H.; Huang, A.J.; Yi, L.; et al. Silencing of the chitin synthase gene is lethal to the Asian citrus psyllid, Diaphorina citri. Int. J. Mol. Sci. 2019, 20, 3734. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.Q.; Zhou, Z.; Sun, K.D.; Huang, B.; Stanley, D.; Song, Q.S. The antibiotic jinggangmycin increases brown planthopper (BPH) fecundity by enhancing rice plant sugar concentrations and BPH insulin-like signaling. Chemosphere 2020, 249, 126463. [Google Scholar] [CrossRef]
- Zhao, X.M.; Jia, P.; Zhang, J.; Yang, Y.; Liu, W.M.; Zhang, J.Z. Structural glycoprotein LmAbd-9 is required for the formation of the endocuticle during locust molting. Int. J. Biol. Macromol. 2019, 125, 588–595. [Google Scholar] [CrossRef]
- Hettenhausen, C.; Schuman, M.C.; Wu, J.Q. MAPK signaling: A key element in plant defense response to insects. Insect Sci. 2015, 22, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Toufeeq, S.; Yu, H.Z.; Wang, J.; Zhang, S.Z.; Li, B.; Li, Z.; Yang, L.A.; Hu, P.; Ma, Y.; et al. Molecular characterization of two mitogen-activated protein kinases: p38 MAP kinase and ribosomal S6 kinase from Bombyx mori (Lepidoptera: Bombycidae), and insight into their roles in response to BmNPV infection. J. Insect Sci. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Jin, K.; Hang, L.R.; Xia, Y.X. MaMk1, a FUS3/KSS1-type mitogen-activated protein kinase gene, is required for appressorium formation, and insect cuticle penetration of the entomopathogenic fungus Metarhizium acridum. J. Invertebr. Pathol. 2014, 115, 68–75. [Google Scholar] [CrossRef]
- Meng, Y.M.; Zhang, X.; Guo, N.; Fang, W.G. MrSt12 implicated in the regulation of transcription factor AFTF1 by Fus3-MAPK during cuticle penetration by the entomopathogenic fungus Metarhizium robertsii. Fungal Genet. Biol. 2019, 131, 103244. [Google Scholar] [CrossRef]
- Gao, X.L.; Fu, Y.T.; Ajayi, O.E.; Guo, D.Y.; Zhang, L.Q.; Wu, Q.F. Identification of genes underlying phenotypic plasticity of wing size via insulin signaling pathway by network-based analysis in Sogatella furcifera. BMC Genom. 2019, 20, 396. [Google Scholar] [CrossRef]
- Lin, X.D.; Yao, Y.; Wang, B.; Lavine, M.D.; Lavine, L.C. FOXO links wing form polyphenism and wound healing in the brown planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2016, 70, 24–31. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | Application |
|---|---|---|
| DcCP64 | F1: GCATTCAATCATCAAACCGAG | RT-qPCR analysis |
| R1: AGACCAAACAGACCACGCTTC | ||
| GAPDH | F2: CATGGCAAGTTCAACGGTGA | |
| R2: CGATGCCTTCTCAATGGTGG | ||
| DcCP64 | F3: TAATACGACTCACTATAGGGGAAGCGTGGTCTGTTTG | dsRNA synthesis |
| R3: TAATACGACTCACTATAGGGTGGGTAGGGTTTTGGTA | ||
| GFP | F4: TAATACGACTCACTATAGGGCAGTGCTTCAGCCGCTACCC | |
| R4: TAATACGACTCACTATAGGGACTCCAGCAGGACCATGTGAT | ||
| DcCP64 | F5: CCGGAATTCGAGGACAAGAAAGTCGCA | Protein expression |
| R5: CCGCTCGAGGTGGTGGTAGTTGTAACTGTAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Yi, L.; Lu, Z. Silencing of Chitin-Binding Protein with PYPV-Rich Domain Impairs Cuticle and Wing Development in the Asian Citrus Psyllid, Diaphorina citri. Insects 2022, 13, 353. https://doi.org/10.3390/insects13040353
Yu H, Yi L, Lu Z. Silencing of Chitin-Binding Protein with PYPV-Rich Domain Impairs Cuticle and Wing Development in the Asian Citrus Psyllid, Diaphorina citri. Insects. 2022; 13(4):353. https://doi.org/10.3390/insects13040353
Chicago/Turabian StyleYu, Haizhong, Long Yi, and Zhanjun Lu. 2022. "Silencing of Chitin-Binding Protein with PYPV-Rich Domain Impairs Cuticle and Wing Development in the Asian Citrus Psyllid, Diaphorina citri" Insects 13, no. 4: 353. https://doi.org/10.3390/insects13040353
APA StyleYu, H., Yi, L., & Lu, Z. (2022). Silencing of Chitin-Binding Protein with PYPV-Rich Domain Impairs Cuticle and Wing Development in the Asian Citrus Psyllid, Diaphorina citri. Insects, 13(4), 353. https://doi.org/10.3390/insects13040353