1. Introduction
Competition for food has often been observed in ants [
1], and many worker ants are killed in competition for food resources [
2,
3]. Mortality due to food competition is an important threat to the survival of early ant colonies [
4]. Competition is affected by various factors, including natural or anthropogenic disturbance and temperature [
5,
6]. Among them, temperature is expected to significantly affect competition by affecting the foraging and metabolic rate of species [
5], and the intensity of competition varies by species. Ant activity levels and trophic preferences shift with temperature changes [
7]. Seifert [
8] was the first who showed that competitive exclusion in multi-species communities in nature exists and that interspecific competition increases with relatedness. Frizzi, et al. [
9] revealed the role of environmental temperature in affecting the survival ability of an invasive garden ant,
Lasius neglectus, in competition with other species. Cerdá, et al. [
10] studied the role of competition with dominant species and temperature in the foraging of subordinate ant species and suggested that the foraging of subordinate species was influenced more by temperature than by competition with dominant species. Kwon [
11] revealed that competition between ants for food varied with temperature in South Korea. Food competition was greatest at intermediate temperatures, decreasing at both low and high temperatures. The reason for this was that the species involved in intense competition for food were usually found at intermediate temperatures, and the prevalence of such species decreased when the temperature was lowered or higher. Therefore, mortality due to food competition is likely to be affected by temperature. However, few studies have examined the effect of temperature on mortality due to competition for food in ants in the field. Therefore, we aimed to test the hypothesis that mortality due to food competition would increase at intermediate temperatures in forest ant species.
The co-occurrent ants, even closely related species, show different food preferences [
12,
13], and body size, aggression, recruitment behavior, and activity timing vary depending on the species [
14,
15]. Owing to differences in behavioral characteristics and body size between ant species, different ant colonies can consist of aggressive dominant species or submissive subordinate species [
8,
16]. A competitive hierarchy is created among ants gathered at large-sized food [
17,
18]. In competition for food, subordinate species can have a higher mortality rate than dominant species. In addition, it is expected that large-bodied individuals can kill small-bodied individuals more than vice versa. Food competition between ants is fierce not only between different species but also within the same species [
19,
20]. The intensity of inter- and intraspecific competition for food can vary between species. In particular, in the case of subordinate species, if the mortality rate due to interspecific competition decreases, then the mortality rate due to intraspecific competition is expected to increase. However, in dominant species that are less affected by other species, a significant relationship between interspecific competition mortality and intraspecific competition mortality would not be expected.
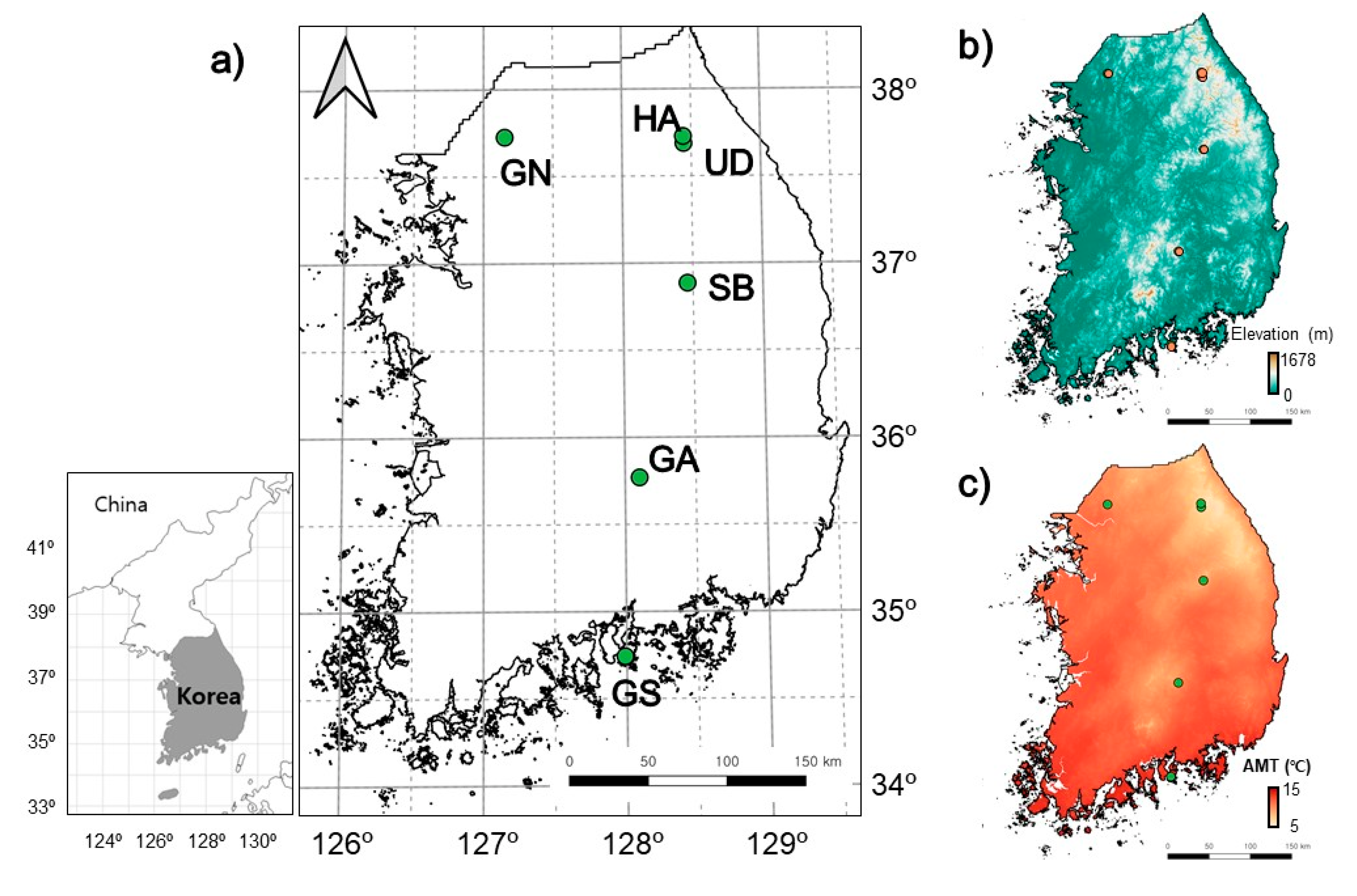
To test these expectations, we investigated the mortality rate due to food competition among ants in six regions under different temperature conditions from the southernmost area to the northernmost area of South Korea.
3. Results
In bait experiments, 16 species with 134,030 ant individual observations were recorded (
Table 1).
Myrmica spp. and
Camponotus atrox Emery, 1925 were mainly observed at low-temperature sites, whereas
A. japonica,
Lasius japonicus Santschi, 1941, and
P. fervida were observed at most sites, showing a hump-shaped distribution with a larger number of individuals at intermediate temperatures. The most abundant species was
N. flavipes, whose occurrence increased as a function of temperature.
In the experiment, 173 individuals (0.13% of the total number of visitors) were killed in the context of intra- and inter-species competition for food. Small-bodied and abundant species, such as
N. flavipes and
P. fervida, had a high number of victims (i.e., killed ants) (
Table 2 and
Table 3).
N. flavipes, the most abundant species, showed the highest number of victims with 76 individuals (44% of the total victims; 0.12% of its abundance), followed by
P. fervida, the second most abundant species, with 61 individuals (35% of total victims; 0.16% of its abundance) (
Table 2 and
Table 4). Although the number of individuals killed in the context of interspecific competition was similar between the two species, the number of individuals killed in the context of intraspecific competition was higher in the subordinate species,
N. flavipes (19 individuals), than in the dominant species,
P. fervida (three individuals) (
Table 2 and
Table 4). Victims of
N. flavipes due to intraspecific competition occurred mainly at the highest temperature site (89%), where the mortality due to interspecific competition was very low. Meanwhile,
Myrmica spp. showed the highest victim rate at 0.53% (16 out of 2996 observed individuals). As the dead ants were transported to the nests of their killers, it is likely that they were used for food by inter- or intraspecific individuals (
Table 5).
P. fervida was the only species with soldier ants (5.4% of observed individuals). Among the dead ants, five soldiers were killed by
Formica japonica and
A. japonica, which were larger than
P. fervida (
Table 3).
A. japonica, a dominant species with a relatively large body, was also frequently killed. The number of
A. japonica individuals killed by intraspecific competition (12 individuals) was higher than that killed by interspecific competition (seven individuals) (
Table 4). In the case of mortality due to interspecific competition, there were 131 cases in which large species killed small species, but only five cases in which smaller species killed larger species (χ
2 = 116.7,
p < 0.001;
Table 2 and
Table 3). There were also 131 cases in which dominant species killed subordinate species, but only four cases in which subordinate species killed dominant species (χ
2 = 119.5,
p < 0.001;
Table 2 and
Table 3).
The mortality rate due to intraspecific competition was in the range of 0.01–0.1%, whereas that due to interspecific competition was in the range of 0.12–0.53%, indicating a higher mortality rate from interspecific competition (
Table 4).
Myrmica spp. had the highest mortality rate in general due to intra- and interspecific competition. In the subordinate species,
N. flavipes and
Myrmica spp., the interspecific competition mortality rate was 3 and 4.3 times higher than that of intraspecific competition, respectively.
P. fervida, a small dominant species, showed a considerably higher interspecific mortality rate compared with that of intraspecific competition (19.3 times), whereas the intraspecific competitive mortality rate for
A. japonica, a medium-sized dominant species, was twice that for interspecific competition. The mortality rate might be underestimated due to the repeated counts of same individuals.
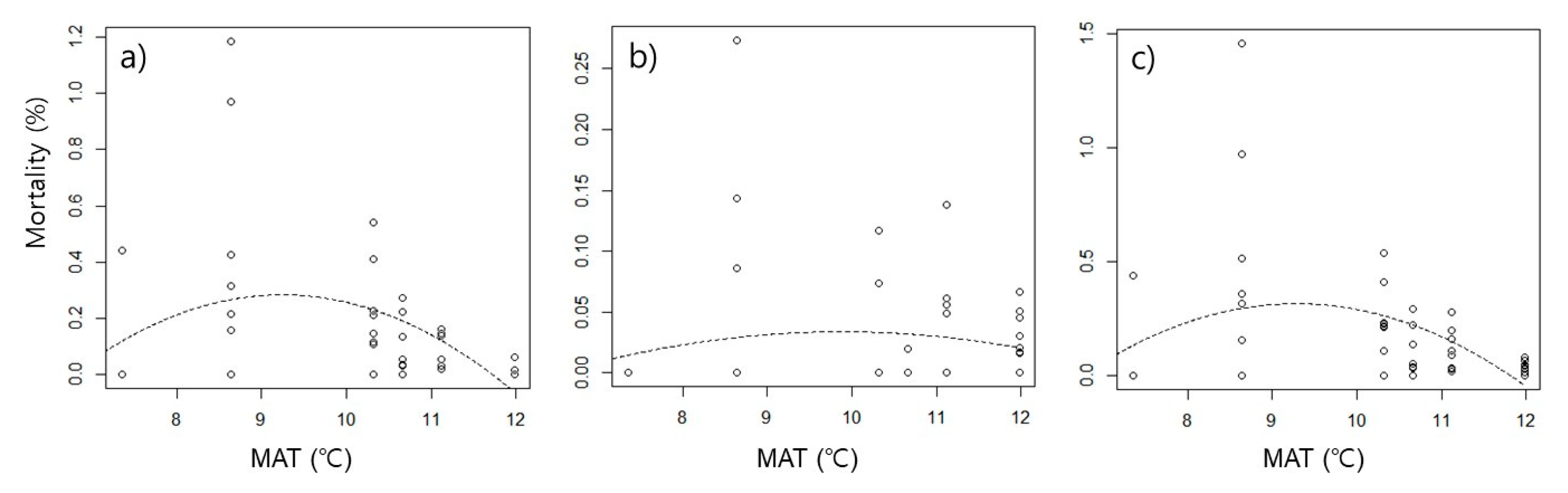
The mortality rate caused by food competition was higher at intermediate temperatures than at low and high temperatures, showing hump-shaped curves (
Table 2,
Figure 2). The peaks of mortality were observed around 9.3 ℃ in interspecific competition, 9.8 ℃ in intraspecific competition, and 9.3 ℃ in total competition. The significance of the hump-shaped trend was determined by the significant effect of MAT
2 (
p < 0.05) (
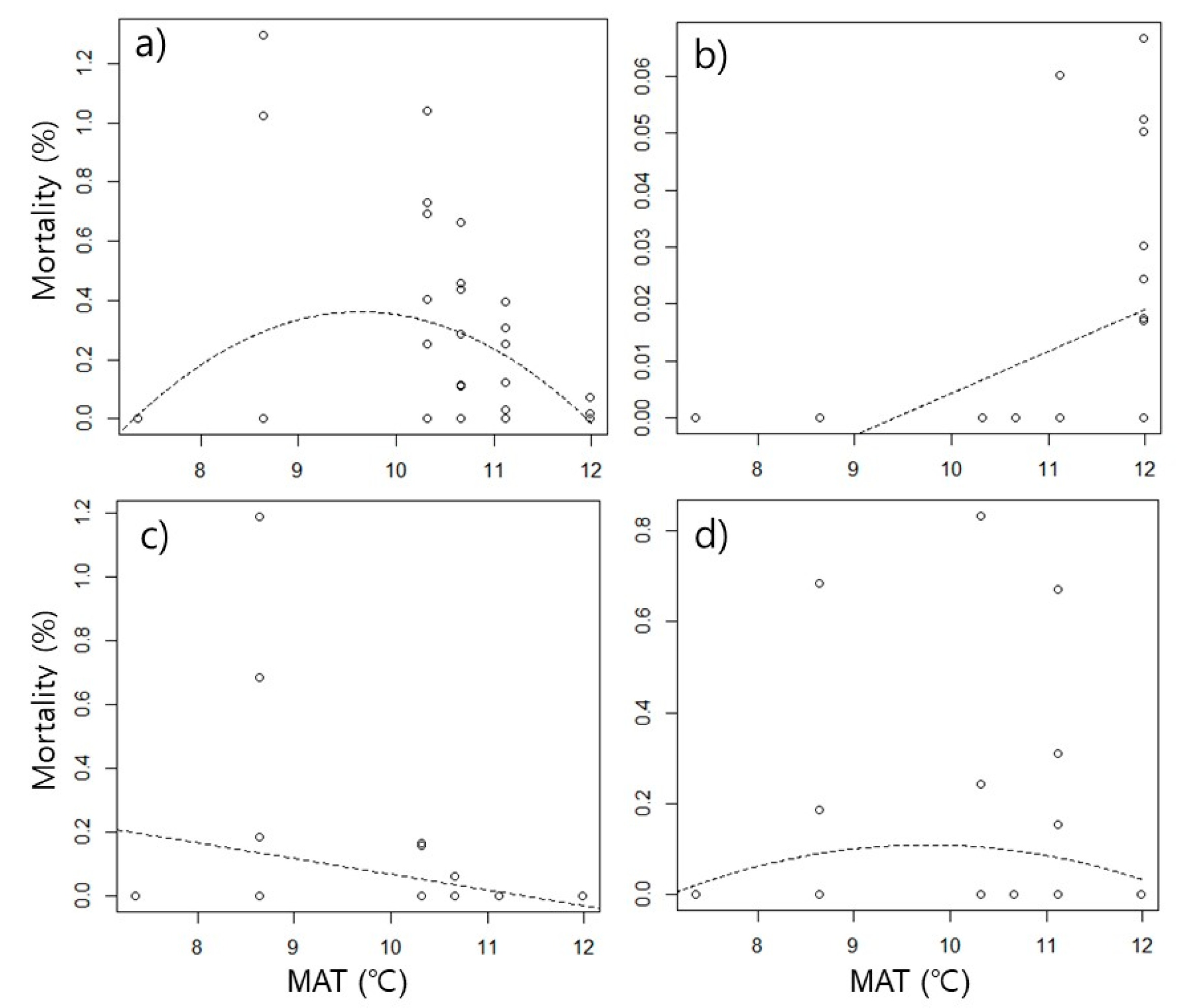
Table 6). However, the trend differed depending on the species. In
N. flavipes, interspecific competitive mortality showed a hump-shaped trend, but intraspecific competitive mortality showed a linear trend that increased as temperature increased (
Figure 3a,b). Mortality due to interspecific competition in
P. fervida displayed a hump-shaped trend (
Table 6), but intraspecific competition was not analyzed because of the low frequency (n = 3).
A. japonica showed a contrasting trend to
N. flavipes. In interspecific competition, a linear trend that increased at low temperatures was observed (
Figure 3c,
Table 6), and in intraspecific competition, a hump-shaped trend was observed (
Figure 3d,
Table 6). The peaks of mortality were observed around 9.7 ℃ in interspecific competition of
N. flavipes, and 9.8 ℃ in intraspecific competition of
A. japonica.
5. Conclusions
The mortality rate of ants due to food competition was between 0% and 0.53%. However, this was significantly underestimated due to the repeated counts of same individuals. Contrary to expectations, mortality due to interspecific competition was significantly higher than that due to intraspecific competition. However, the dominant species, which were less affected by other species, had a higher mortality rate due to intraspecific competition. In the case of the most subordinate species, N. flavipes, the mortality rate due to intraspecific competition increased sharply when mortality by other species decreased. The authors hypothesized that mortality due to food competition would decrease at intermediate and high temperatures but increase at low temperatures as climate warms, which was evidenced by the results. Given the effect of temperature on worker mortality due to food competition, it is expected that changes in competition due to climate change can affect the fitness of ant species in the future. The results of this study showed that the mortality of ants due to food competition is determined not only by physical environmental factors (temperature) but also by biological factors, such as the abundance and characteristics of species (body size, dominance, and aggressiveness).








{kind=link}
{kind=link}
{kind=link}