
Installation Season May Significantly Impact Time Required for Subterranean Termites to Find and Feed on In-Ground Baits


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
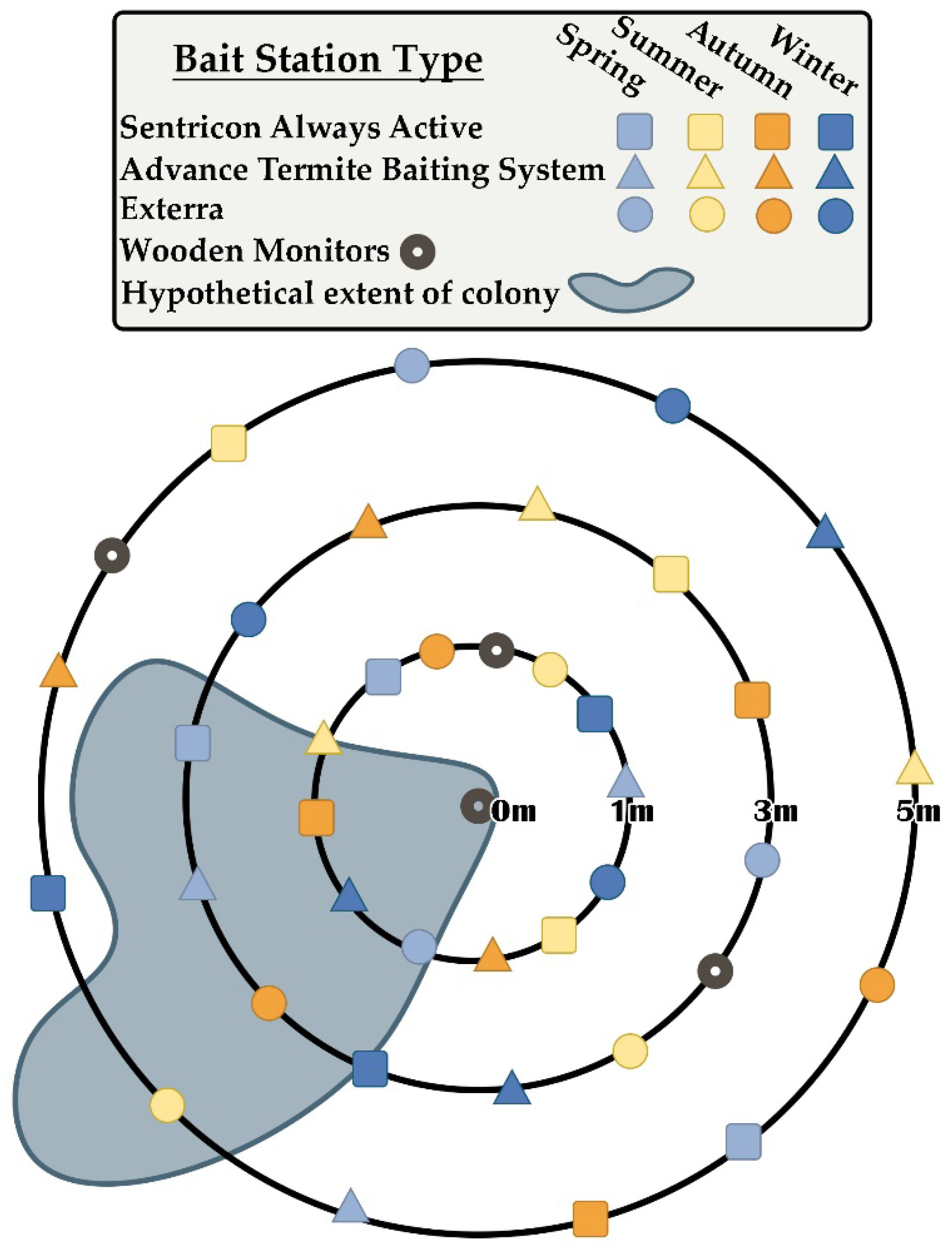
2. Materials and Methods
3. Results
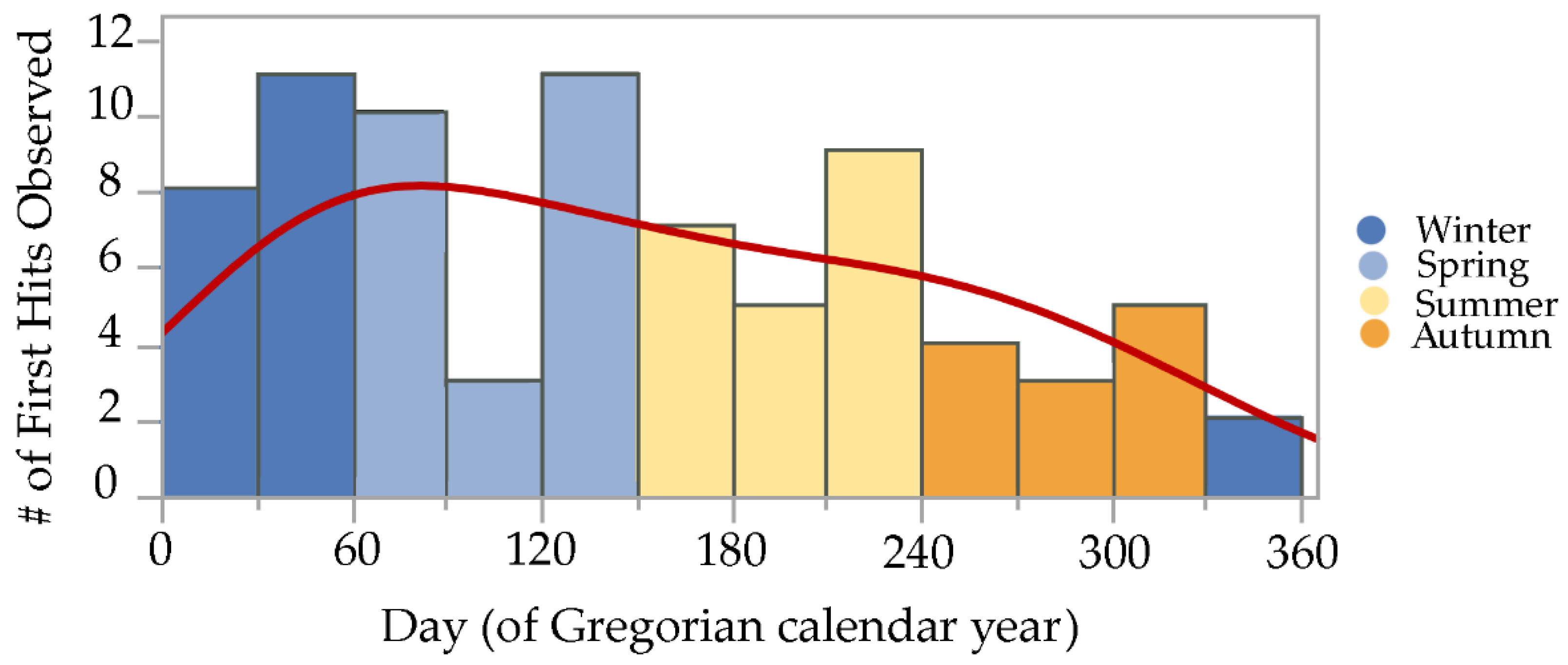
3.1. Overall Seasonal Activity
3.2. Observed Bait Interception Time
3.2.1. Bivariate Relationships: Effects on Bait Interception Time
3.2.2. Effect of Installation Season on Bait Interception Time
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kakkar, G.; Su, N.Y. Molting drives mortality in foraging populations of Formosan subterranean termites (Isoptera: Rhinotermitidae) baited with a chitin synthesis inhibitor, noviflumuron. Pest Manag. Sci. 2018, 74, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Su, N.-Y. Managing Social Insects Urban Importance. Annu. Rev. Entomol. 2012, 57, 535–575. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.T.; Borges, P.A.; Nunes, L.; Myles, T.G.; Guerreiro, O.; Scheffrahn, R.H. Termites (Isoptera) in the Azores: An overview of the four invasive species currently present in the archipelago. Arquipélago Life Mar. Sci. 2013, 30, 39–55. [Google Scholar]
- Lewis, V.R.; Haverty, M.I.; Carver, D.S.; Fouche, C. Field comparison of sand or insecticide barriers for control of Reticulitermes spp. (Isoptera: Rhinotermitidae) infestations in homes in Northern California. Sociobiology 1996, 28, 327–335. [Google Scholar]
- Haverty, M.I.; Getty, G.M.; Copren, K.A.; Lewis, V.R. Seasonal foraging and feeding behavior of Reticulitermes spp. (Isoptera: Rhinotermitidae) in a wildland and a residential location in northern California. Environ. Entomol. 1999, 28, 1077–1084. [Google Scholar] [CrossRef]
- Chouvenc, T. Comparative impact of chitin synthesis inhibitor baits and non-repellent liquid termiticides on subterranean termite colonies over foraging distances: Colony elimination versus localized termite exclusion. J. Econ. Entomol. 2018, 111, 2317–2328. [Google Scholar] [CrossRef]
- Quarles, W. IPM for Termites—Termite Baits. IPM Pract. 2003, 25, 1–9. [Google Scholar]
- Kakkar, G.; Chouvenc, T.; Osbrink, W.; Su, N.Y. Temporal assessment of molting in workers of Formosan subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2016, 109, 2175–2181. [Google Scholar] [CrossRef]
- Chouvenc, T.; Lee, S.B. Queen Egg Laying and Egg Hatching Abilities are Hindered in Subterranean Termite Colonies When Exposed to a Chitin Synthesis Inhibitor Bait Formulation. J. Econ. Entomol. 2021, 114, 2466–2472. [Google Scholar] [CrossRef]
- Eger, J.E., Jr.; Lees, M.D.; Neese, P.A.; Atkinson, T.H.; Thoms, E.M.; Messenger, M.T.; Demark, J.J.; Lee, L.C.; Vargo, E.L.; Tolley, M.P. Elimination of subterranean termite (Isoptera: Rhinotermitidae) colonies using a refined cellulose bait matrix containing noviflumuron when monitored and replenished quarterly. J. Econ. Entomol. 2012, 105, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Shults, P.; Richardson, S.; Eyer, P.A.; Chura, M.; Barreda, H.; Davis, R.W.; Vargo, E.L. Area-wide elimination of subterranean termite colonies using a novaluron bait. Insects 2021, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Iqbal, N. Termite (order Blattodea, infraorder Isoptera) baiting 20 years after commercial release. Pest Manag. Sci. 2015, 71, 897–906. [Google Scholar] [CrossRef]
- Chouvenc, T. Subterranean Termite (Coptotermes gestroi (Blattodea: Rhinotermitidae)) colony elimination through exposure to a novaluron CSI bait formulation in laboratory. J. Econ. Entomol. 2021, 114, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Su, N.Y. Development of baits for population management of subterranean termites. Annu. Rev. Entomol. 2019, 64, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Su, N.Y.; Monteagudo, E.J. Hyperecdysonism in the Formosan subterranean termite and eastern subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2017, 110, 1736–1739. [Google Scholar] [CrossRef]
- Getty, G.M.; Solek, C.W.; Sbragia, R.J.; Haverty, M.I.; Lewis, V.R. Large-scale Suppression of a Subterranean Termite Community Using the Sentricon® Termite Colony Elimination System: A Case Study in Chats-worth, California, USA. Sociobiology 2007, 50, 1041–1050. [Google Scholar]
- Shelton, T.G.; Wagner, T.L.; Gerard, P.D. Effects of monitor examination intervals on resource affinity by Reticulitermes spp. (Isoptera: Rhinotermitidae). J. Entomol. Sci. 2011, 46, 102–111. [Google Scholar] [CrossRef]
- Thoms, E.M.; Eger, J.E.; Messenger, M.T.; Vargo, E.; Cabrera, B.; Riegel, C.; Murphree, S.; Mauldin, J.; Scherer, P. Bugs, baits, and bureaucracy: Completing the first termite bait efficacy trials (Quarterly replenishment of noviflumuron) initiated after adoption of Florida rule. Am. Entomol. 2009, 55, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Keefer, T.C.; Puckett, R.T.; Brown, K.S.; Gold, R.E. Field trials with 0.5% novaluron insecticide applied as a bait to control subterranean termites (Reticulitermes sp. and Coptotermes formosanus [Isoptera: Rhinotermitidae]) on structures. J. Econ. Entomol. 2015, 108, 2407–2413. [Google Scholar] [CrossRef]
- Kauffman, E. Climate and topography. In Atlas of the Biodiversity of California; California Department of Fish and Game: Sacramento, CA, USA, 2003; pp. 12–15. [Google Scholar]
- Haagsma, K.A.; Rust, M.K. Colony size estimates, foraging trends, and physiological characteristics of the western subterranean termite (Isoptera: Rhinotermitidae). Environ. Entomol. 1995, 24, 1520–1528. [Google Scholar] [CrossRef]
- Haverty, M.I.; Getty, G.M.; Nelson, L.J.; Lewis, V.R. Flight phenology of sympatric populations of Reticulitermes (Isoptera: Rhinotermitidae) in northern California: Disparate flight intervals indicate reproductive isolation among cuticular hydrocarbon phenotypes. Ann. Entomol. Soc. 2003, 96, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Härdle, W.; Werwatz, A.; Müller, M.; Sperlich, S. Nonparametric Density Estimation. In Nonparametric and Semiparametric Models; Springer: Berlin, Germany, 2004. [Google Scholar]
- JMP® Statistical Software. Manual for Version Pro 16; SAS Institute Inc.: Cary, NC, USA, 2021. [Google Scholar]
- Woolson, R.F. Wilcoxon Signed-Rank Test. In Wiley Encyclopedia of Clinical Trials; D’Agostino, R.B., Sullivan, L., Massaro, J., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Hox, J.J. Multilevel Analysis: Techniques and Applications; Routledge: Oxfordshire, UK, 2010. [Google Scholar]
- Hu, X.P.; Appel, A.G.; Traniello, J.F. Behavioral response of two subterranean termites (Isoptera: Rhinotermitidae) to vibrational stimuli. J. Insect Behav. 2003, 16, 703–715. [Google Scholar] [CrossRef]
- Green, F., III; Arango, R.; Esenther, G. Community-wide suppression of R. flavipes from Endeavor, Wisconsin-search for the holy grail. Int. Res. Group Wood Prot. 2008, IRG/WP, 08-10674. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutherland, A.M.; Hubble, C.; Barber, M. Installation Season May Significantly Impact Time Required for Subterranean Termites to Find and Feed on In-Ground Baits. Insects 2022, 13, 445. https://doi.org/10.3390/insects13050445
Sutherland AM, Hubble C, Barber M. Installation Season May Significantly Impact Time Required for Subterranean Termites to Find and Feed on In-Ground Baits. Insects. 2022; 13(5):445. https://doi.org/10.3390/insects13050445
Chicago/Turabian StyleSutherland, Andrew M., Casey Hubble, and Molly Barber. 2022. "Installation Season May Significantly Impact Time Required for Subterranean Termites to Find and Feed on In-Ground Baits" Insects 13, no. 5: 445. https://doi.org/10.3390/insects13050445
APA StyleSutherland, A. M., Hubble, C., & Barber, M. (2022). Installation Season May Significantly Impact Time Required for Subterranean Termites to Find and Feed on In-Ground Baits. Insects, 13(5), 445. https://doi.org/10.3390/insects13050445