Simple Summary
Tree of heaven, Ailanthus altissima, is a deciduous tree indigenous to China and introduced for ornamental purposes into North America and Europe. It shows a highly invasive profile in field, urban, and suburban areas, posing a serious threat to ecosystems in the introduced ranges. The current management of this noxious infesting plant by mechanical and chemical measures remains ephemeral and incomplete. A multi-tactic approach emphasizing classical biological control appears highly relevant. Eriophyid mites are well known for their high specificity and for the impact caused to the associated hosts, resulting in relevant potential biological control agents of infesting plants. The partially unresolved taxonomy of the eriophyid mite species reported on the tree of heaven is currently seen as an impediment to their further concern as biocontrol agents. This paper investigated morphological and molecular characters of Aculus mosoniensis in order to better clarify its taxonomic status. A paratype specimen of Aculops taihangensis was also studied, and this mite species was reassigned to the genus Aculus. The strong congruence between morphological and molecular analyses for all mites collected on tree of heaven in Europe led to the conclusion that A. mosoniensis is a junior synonym of Ac. taihangensis.
Abstract
The taxonomy of Aculus mosoniensis appears to be an unresolved question and its clarification is required, owing to the potential relevance of this mite species as a biological control agent of the tree of heaven. This paper is aimed at giving accurate details on a previously and shortly announced synonymization with Aculops taihangensis, using a morphological and molecular approach. A fusiform morph of A. mosoniensis was distinguished from a vermiform morph and this latter was recognized as deutogyne, which was herein documented. Phylogenetic relationships between Chinese Ac. taihangensis and all A. mosoniensis mites collected in twenty localities in Europe were examined through the analysis of the mitochondrial cytochrome c subunit I (CO1) protein and the nuclear ribosomal internal transcribed spacer 1 region (ITS1). CO1 sequences of Ac. taihangensis from the Shandong province in China and those from mites collected in Austria and Slovenia were 100% identical; the ITS1 sequence of an Ac. taihangensis paratype matched for 99.8% with those obtained from protogynes and deutogynes of A. mosoniensis collected in Italy. All these data supported the announced synonymization of A. mosoniensis with Ac. taihangensis. Aculus mosoniensis was found genetically variable, with five CO1 haplotypes in Europe (becoming eight along with those of Ac. taihangensis) clustering in two highly supported maternal lineages and eight ITS1 haplotypes (becoming nine along with those of Ac. taihangensis) distributed in four supported clades. No overlap between intra- and interspecies distances was observed for both markers and all studied A. mosoniensis populations clustered in one monophyletic mitochondrial clade, suggesting that only one single species might occur in Europe. However, more mite clades may be related to more tree of heaven biotypes with potential ecological differences, which might have potential effects on the biological control and should be investigated.
1. Introduction
The tree of heaven (ToH), Ailanthus altissima (Mill.) Swingle, belongs to Simaroubaceae (Sapindales), which is a family of Eastern Asian origin [1]. Ailanthus altissima is a deciduous tree species, alien in Europe and North America, and considered as one of the most invasive and noxious infesting plants worldwide favored by its bioecological characteristics [2,3,4,5]. Eriophyid mites (Acari: Eriophyoidea) are highly specialized plant feeders, and most of them are well known for their high specificity and for the impact they have on the associated hosts, despite their extremely small size [6]. Aculops ailanthi Lin, Jin & Kuang, Aculops taihangensis Hong & Xue (the new combination of the genus name was assigned only at the end of this paper), Aculus altissimae Xue & Hong, and Aculus mosoniensis (Ripka) have been found associated with A. altissima [7,8]. All of them have been recorded in China, except for A. mosoniensis, which has been reported only in European countries [6], and Ac. Ailanthi, also found in the USA [9,10]. Aculus mosoniensis and Ac. ailanthi are considered very promising biological control agents based on the thus-far records of impact noticed to provoke on their host [6,9,10]. Host-range testing under open-field conditions recently carried out in a European context provided a good indication of the specificity of A. mosoniensis for ToH [6]. As reported in an earlier paper [7], the taxonomic and the bioecological characterization of A. mosoniensis has been under evaluation, and the synonymy between Ac. taihangensis and A. mosoniensis was announced in a short review paper with a very brief and non-exhaustive explanation.
Different procedures of specimen slide mounting can stress the morphology and hide some fundamental details, especially in such tiny eriophyid mites, which can lead to poor descriptions of species and need for their revision [11]. Integrative taxonomy, as coined by Dayrat [12] by combining molecular phylogenetic and phylogeographical information, was demonstrated to be a valid approach to support the taxonomy of eriophyid mites [13,14,15]. A further interference in the correct definition of the taxonomic status of an eriophyid species can come from its life strategy. Overwintering deutogyne morphs, which are characterized by an innate resistance to the environmental stresses, can have a distinctive morphology in respect of spring–summer protogyne morphs [16]. The differences can be noticeable but very pale for some species and require careful morphometric examination [17]. Therefore, in the definition of the taxonomic status of an eriophyid mite, it is worthy of interest to examine the biological cycle in order to verify the existence of these morphs, which can make the identification less easy and lead to the constitution of species which might need, then, to be reassigned and synonymized. This is the case of the eriophyoid species currently known to be associated with ToH: some species have a poor description and no biological data are available. This means that synonymy of A. mosoniensis with other species might be possible.
The taxonomic status of A. mosoniensis, as well as its ecological relationship with the host plant, appear to be unresolved questions and their clarification is required considering the potential relevance of this mite species as a biological control agent for ToH. Therefore, this paper is aimed at elucidating the taxonomic position of A. mosoniensis using morphology and molecular characters, verifying and explaining accurately the announced synonymization along with Ac. taihangensis.
2. Materials and Methods
2.1. Morphological Study by Light Microscopy
Occasional samplings of ToH were carried out from 2016 to 2017 in Apulia (Bari, Italy, 41°06′38″ N, 16°52′55″ E; 19 m a.s.l.) and Lazio (Saxa Rubra, Rome, Italy, 41°58′52″ N, 12°29′38″ E; 18 m a.s.l.). Samples were transported to the laboratory and eriophyid mites were examined under a light microscope. Specimens of A. mosoniensis were slide-mounted according to the protocol described in Baker et al. [18] and Keifer [19]. Only one paratype specimen of Ac. taihangensis preserved in absolute ethanol (tube n. 24, 22 August 2003, Taihang mountain, Hebei province, China, legit Xue Xiao-Feng) was mounted on a slide following the procedure described by Keifer [19].
The terminology and setal notation used in the morphological description of the mites follow mainly those proposed by Lindquist [20]. All morphological measurements were taken using a phase contrast microscope Olympus BX50 according to Amrine and Manson [21], as modified by de Lillo et al. [11], and are given in micrometers (μm). Measurements are rounded off to the nearest integer, referring to the length of the morphological traits unless specified otherwise, and express the range of values of the studied population. Line drawings were handmade through a camera lucida according to de Lillo et al. [11], and figure abbreviations mainly follow Amrine et al. [22].
2.2. Morphological Study by Scanning Electron Microscopy
For scanning electron microscopy observations, leaf and bud samples of ToH were collected in Belgrade, Serbia (44°50′33″ N, 20°24′19″ E, 82 m a.s.l.), on 2 May 2017 (for A. mosoniensis protogynes) and 27 February 2018 (for A. mosoniensis deutogynes), respectively; further bud samples were collected in Bari, Italy (41°06′38″ N, 16°52′55″ E; 19 m a.s.l.), on 16 December 2019 (for A. mosoniensis deutogynes). Live mites were collected with a fine entomological needle from fresh plant parts under a stereomicroscope and transferred on aluminum holders. For Serbian specimens, prior to the observation, holder-mounted mites were gold-coated (BAL-TEC SC-RD005) by a sputter coater (BAL-TEC AG, Balzers, Liechtenstein) for 100 s under 30 mA ion current. Micrographs were recorded by the scanning electron microscope JSM-6390 LV (JEOL GmbH, Munich, Germany) at the Laboratory of Electron Microscopy, Faculty of Agriculture, University of Belgrade (UBS), Serbia. For Italian specimens, holder-mounted mites were treated under a critical point dryer (Leica EM CPF300, Wetzlar, Germany) and gold-coated by a sputter coater (Edwards S150A, Irvine, CA, USA) for 60 s under 30 mA ion current. Micrographs were recorded by scanning electron microscopy (TM3000; Tabletop Microscope, Hitachi, Chiyoda, Japan) at SELGE Laboratory, Dipartimento di Scienze del Suolo, della Pianta e degli Alimenti, University of Bari Aldo Moro (UBI), Italy.
2.3. Sampling for the Molecular Study and DNA Extraction
Live mites were taken directly from ToH leaves in twenty localities distributed across ten European countries, as well as one paratype specimen of Ac. taihangensis (Table 1). All mite specimens were preserved in 96% ethanol in collecting tubes and stored at 4 °C until DNA analysis. The DNA extraction, PCR amplification, and sequencing of all specimens were performed by three different laboratories, i.e., Department of Plant Pests, Institute for Plant Protection and Environment (IPPE, Belgrade, Serbia), the USDA-ARS European Biological Control Laboratory (EBCL, Montpellier, France), and UBI.

Table 1.
Collection information of the Aculus mosoniensis populations and Aculops taihangensis specimens analyzed, and Genbank accession numbers (id = unique numbers for DNA extraction vouchers).
At IPPE, genomic DNA was extracted from a pool of 5–10 specimens using DNeasy Blood & Tissue Kit (Qiagen GmbH, Hilden, Germany), according to the manufacturer’s instructions, with modifications based on Dabert et al. [23].
At EBCL, genomic DNA was extracted from a pool of 5–7 specimens using a different method, as described by Kashefi et al. [24].
At UBI, total genomic DNA was extracted from single specimens by adding 100 µL of Chelex suspension (10% in water) (InstaGene matrixTM—BioRad, Hercules, CA, USA) and 5 µL of proteinase-K (20 mg/μL) (Qiagen). More specifically, the UBI extracted DNA from live mites collected directly from leaves (six protogynes and/or males with fusiform aspect) and from overwintering buds (nine deutogynes with vermiform aspect), as well as from a second paratype specimen of Ac. taihangensis. These mites were individualized in tubes, spun at 10,000 rpm for 10 min at 4 °C, and stored at −20 °C until DNA extraction.
All residual DNA are archived at the place of the DNA extraction, except at UBI where there was no DNA left post-amplification of ITS1, hence preventing the amplification of the barcode region.
2.4. Markers for the Molecular Study
The barcode region of the cytochrome oxidase subunit I gene (CO1) was amplified at IPPE and EBCL as described in Marini et al. [6] and in Kashefi et al. [24], respectively. The amplified fragments were sequenced in both directions with the same primer pairs as in the initial PCR procedure, using ABI technology by Macrogen (Seoul, Korea) for IPPE and Genoscreen (Lille, France) for EBCL. Genbank accession numbers of COI sequences are given in Table 1.
The internal transcribed spacer 1 region of ribosomal DNA (ITS1) primarily targeted by UBI was obtained using the primer pair 18S/5.8S reported in Navajas et al. [25]. PCRs were performed in 20 μL final volume containing 10 μL of Go Taq® Green Master Mix 2X (Promega), 1 μL for each primer 10 μM, 3 μL of water, and 5 μL of DNA template. Touch-down PCR reactions were conducted following Carew et al. [26]. Five μL of amplicons were purified using Illustra™ ExoProStar™ (GE Healthcare Life Sciences) according to the manufacturer’s instructions and sequenced in both senses using ABI technology by Applied Biosystems™ DNA Analyzer 3730XL (Genomic Facility at Foundation Edmund Mach San Michele all’Adige, Trento, Italy).
At IPPE and EBCL, we followed the same procedure as described above except that (i) the DNA template was always diluted, (ii) the PCR composition, apart from the primers, was similar to that used for the barcode, and (iii) sequencing was performed by different companies. The ITS1 products obtained in Colombes3 (France) and TOHA2 (Italy) were further purified using Qiaquick gel extraction kit (Qiagen, Valencia, CA) and sequenced after cloning into the pGEM-T easy vector (Promega, Madison, WI, USA). Sequencing of 10 clones for each ITS1 product was carried out using the universal M13 primers. Genbank accession numbers of ITS1 sequences are given in Table 1.
2.5. Alignments, Phylogenetic Analyses, and Genetic Distance for the Molecular Study
Both strands of each amplicon were assembled into contigs using either FinchTV v.1.4.0 (www.geospiza.com (accessed on 1 April 2022) at IPPE or Bioedit version 7.2 [27] at EBCL and UBI. The CO1 sequences were translated into amino acids to check for the absence of stop codons and frame shifts in this protein coding gene. Direct sequencing of ITS1 resulted in few sites that were heterozygous for single nucleotide polymorphisms in four specimens. The haplotypic phase of these heterozygous genotypes was inferred using PHASE as implemented in DnaSP v6 [28]. With respect to ITS1 clones, only clones represented at least twice were selected, to avoid inclusion of spurious mutation raised from PCR and cloning error which can produce a false spectrum of genomic diversity. Our dataset of 24 CO1 sequences was then merged with twelve sequences of Ac. taihangensis retrieved from the Barcode of Life system (BOLD) and reported by Yin et al. [29]. A total of 36 CO1 and 45 length-variable ITS1 sequences (combining clones and phased genotypes) were aligned by CLUSTAL W, as implemented in MEGA 11 [30]. We obtained a final CO1 ingroup alignment of 658 characters, 79 of which were parsimony informative, and a final ITS1 ingroup alignment of 440 characters (after excluding gaps), 29 of which were parsimony informative. Identical sequences of CO1 and ITS1 were merged to unique haplotypes with DnaSP v6. For both markers, we calculated pairwise genetic distances within A. mosoniensis and between A. mosoniensis, Ac. Taihangensis, and congeners using MEGA 11. We calculated both the K2P distance [31] (since it has often been used in the literature) and the uncorrected p-distance (since K2P could be inappropriate when employed for closely related taxa) [32]. Phylogenetic relationships were inferred in a maximum likelihood framework and 1000 bootstrap replicates in MEGA 11, and in a Bayesian framework in MrBayes 3.2 [33], with two independent analyses of four Markov chains calculated for 1 million generations, and a 25% “burn-in”. The most appropriate model of evolution selected based on the Bayesian information criterion (BIC [34]), as implemented in MEGA 11, was HKY + G [35] and JC [36] for CO1 and ITS1, respectively. As a priori outgroups for the phylogenetic analyses were chosen: Aculus ichnocarpae (Ghosh & Chakrabarti) (Genbank accession number KM111094.1) and Aculus amygdali Xue & Hong (KM111095.1) for CO1, and Aculus cercidis (Hall) (KJ209712.1) for ITS1.
3. Results
3.1. Morphological Study
Winter and spring–summer populations of A. mosoniensis were observed and studied. The descriptions of males and spring–summer females were previously given [7,8]. A mixed population of fusiform and vermiform mites was observed on leaves collected during autumn (Saxa Rubra, 13 November 2015) in a preliminary collection. The first morph was composed of few females and males, whereas the second one was composed only of females. A further check was made on the mite population recovered into the buds (Saxa Rubra, 21 February 2017) and only vermiform females were found in them. During spring and summer, vermiform females were not found anymore. Therefore, the vermiform was retained to be the deutogynes of A. mosoniensis. Deutogynes were also collected in Bari (Italy) and Serbia. The morphology of the Italian deutogyne is reported below.
3.2. Description
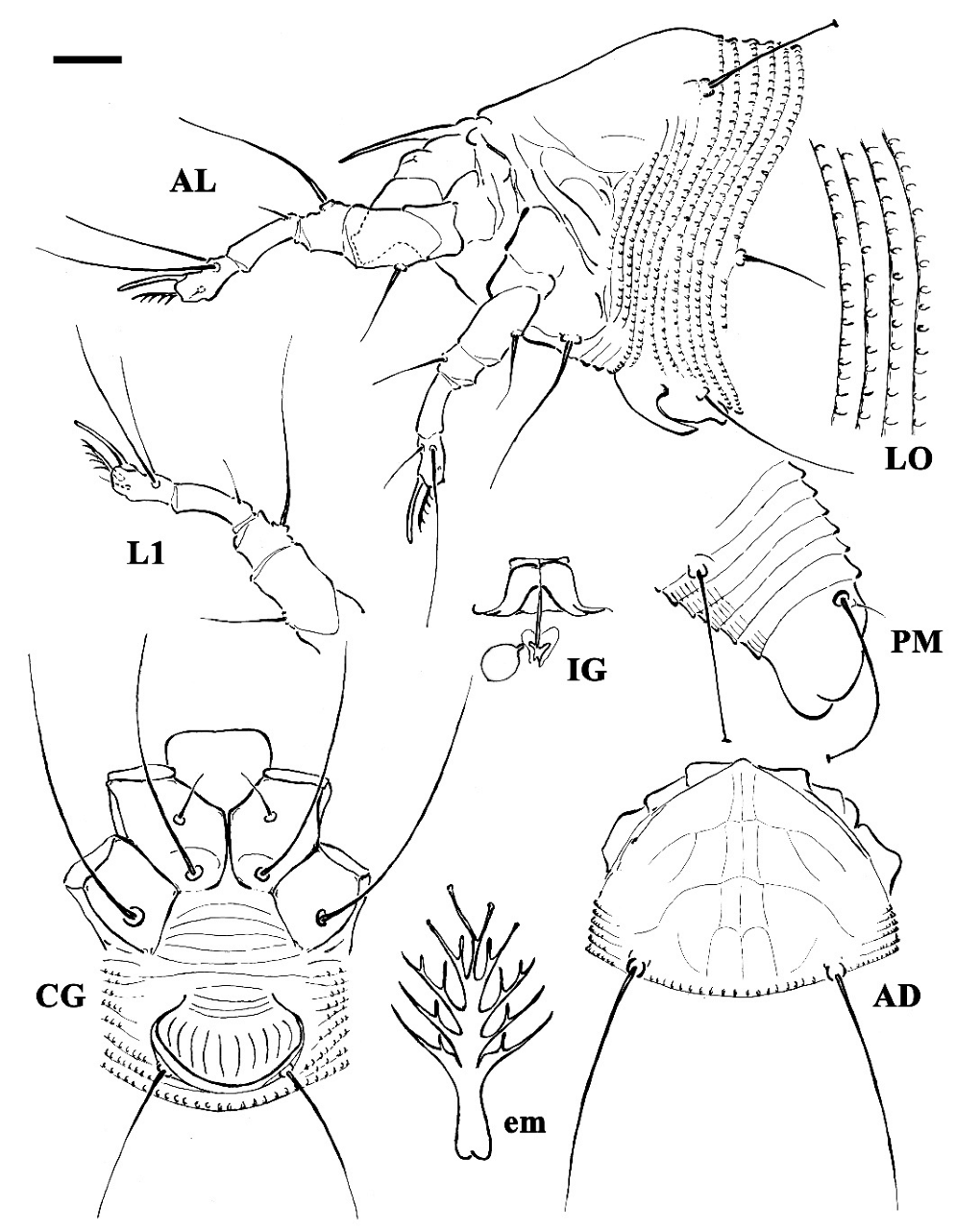
DEUTOGYNE: (n = 18 measured specimens) (Figure 1, Figure 2 and Figure 3). Body vermiform, 194–292 (including gnathosoma), 42–56 wide, 42–60 thick. Gnathosoma 21–25 projecting downwards, pedipalp coxal setae ep 2–3, dorsal pedipalp genual setae d 2–3, unbranched, pedipalp tarsus setae v undetectable, cheliceral stylets 16–19. Prodorsal shield 35–40, including the frontal lobe, 35–45 wide; frontal lobe 2 over gnathosomal base, anteriorly rounded in dorsal view. Network shield pattern often obscure and pale, composed of lines weaker and thinner than that of the protogyne: complete median line and complete pair of admedian lines connected by three transversal lines forming four pairs of cells; two pairs of transverse lines extending outermost from the admedian lines on the half and on the anterior 1/3 of the shield. Tubercles sc quite rounded, on the rear shield margin, 22–26 apart, scapular setae sc 36–43, directed backward. Leg I 41–46, femur 13–15, genu 6–7, tibia 13–13, tarsus 9–10, solenidion ω 9–10, quite straight, distally rounded, empodium 6–7, simple, 5-rayed; femoral setae bv 11–14, genual setae l′′ 23–28, tibial setae l′ 4–7, tarsal setae ft′ 21–23, setae ft′′ 24–29. Leg II 36–42, femur 12–14, genu 6–7, tibia 10–11, tarsus 9–10, solenidion ω 9–10, quite straight, distally rounded, empodium 6–7, simple 5-rayed; femoral setae bv 11–13, genual setae l′′ 6–7, tarsal setae ft′ 5–8, setae ft′′ 24–32. Coxae I and II smooth; setae 1b 10–14, tubercles 1b 9–11 apart, setae 1a 32–45, tubercles 1a 6–7 apart, setae 2a 51–63, tubercles 2a 20–24 apart, prosternal apodeme 9–11. Opisthosoma dorsally arched with 66–76 dorsal semiannuli mainly smooth or with very weak and not-well-delimited microtubercles; 67–80 ventral semiannuli, with rounded microtubercles close to rear margin; 5–6 smooth semiannuli between coxae and genital coverflap; last 5 ventral and dorsal semiannuli with elongated microtubercles. Setae c2 17–26, on ventral semiannulus 8–10; setae d 52–83, on ventral semiannulus 23–27; setae e 22–27, on ventral semiannulus 42–48; setae f 36–47, on ventral semiannulus 63–75, 5 annuli after setae f. Setae h2 62–150 (the variability depends on the age of the mites—the oldest are those on the leaves in early spring and showed longer h2 setae in respect to those collected from the buds in the previous winter), setae h1 4–5. Genital coverflap 10–14, 21–25 wide, coverflap with 8–10 longitudinal striae, setae 3a 20–28, 14–17 apart; 3 close lines at genital coverflap base.
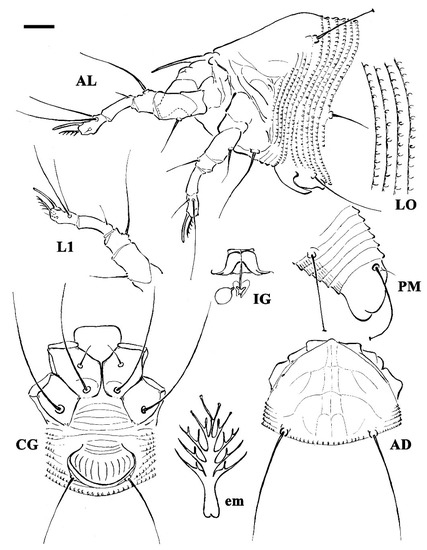
Figure 1.
Line drawings of a deutogyne of Aculus mosoniensis. AD: prodorsal shield; AL: lateral view of anterior body region; CG: female coxigenital region; em: empodium; IG: internal female genitalia; LO: lateral view of annuli; L1: leg I; PM: lateral view of posterior opisthosoma. Scale bar: 10 μm for AD, AL, CG, IG, and PM; 5 μm for LO and L1; 2.5 μm for em.

Figure 2.
Scanning electron micrographs of deutogynes of Aculus mosoniensis: (A) dorsal view; (B) leg tarsal empodium and solenidion; (C) coxigenital region; (D) dorsal view of gnathosoma and frontal lobe. Scale bars: 10 μm for (A,B,D); 2 μm for (B).

Figure 3.
Scanning electron micrographs of a protogyne (A) and a deutogyne (B) of Aculus mosoniensis. Prodorsal shield and part of dorsal opisthosoma. Scale bars: 10 μm.
Basic morphological differences between protogyne and deutogyne are reflecting in more pronounced ornamentation of prodorsal shield in protogyne and in appearance of dorsal opisthosoma that is supplied with elliptical elongate microtubercles in protogynes and is mainly smooth or with very weak and not-well-delimited microtubercles in deutogynes. This is in accordance with the description of most of the deutogynes, and the explanation is in connection with the reduction of the body surface due to the desiccation during unfavorable winter conditions. Number of empodial rays and the genital striae are the same in both female morphs (Figure 2 and Figure 3).
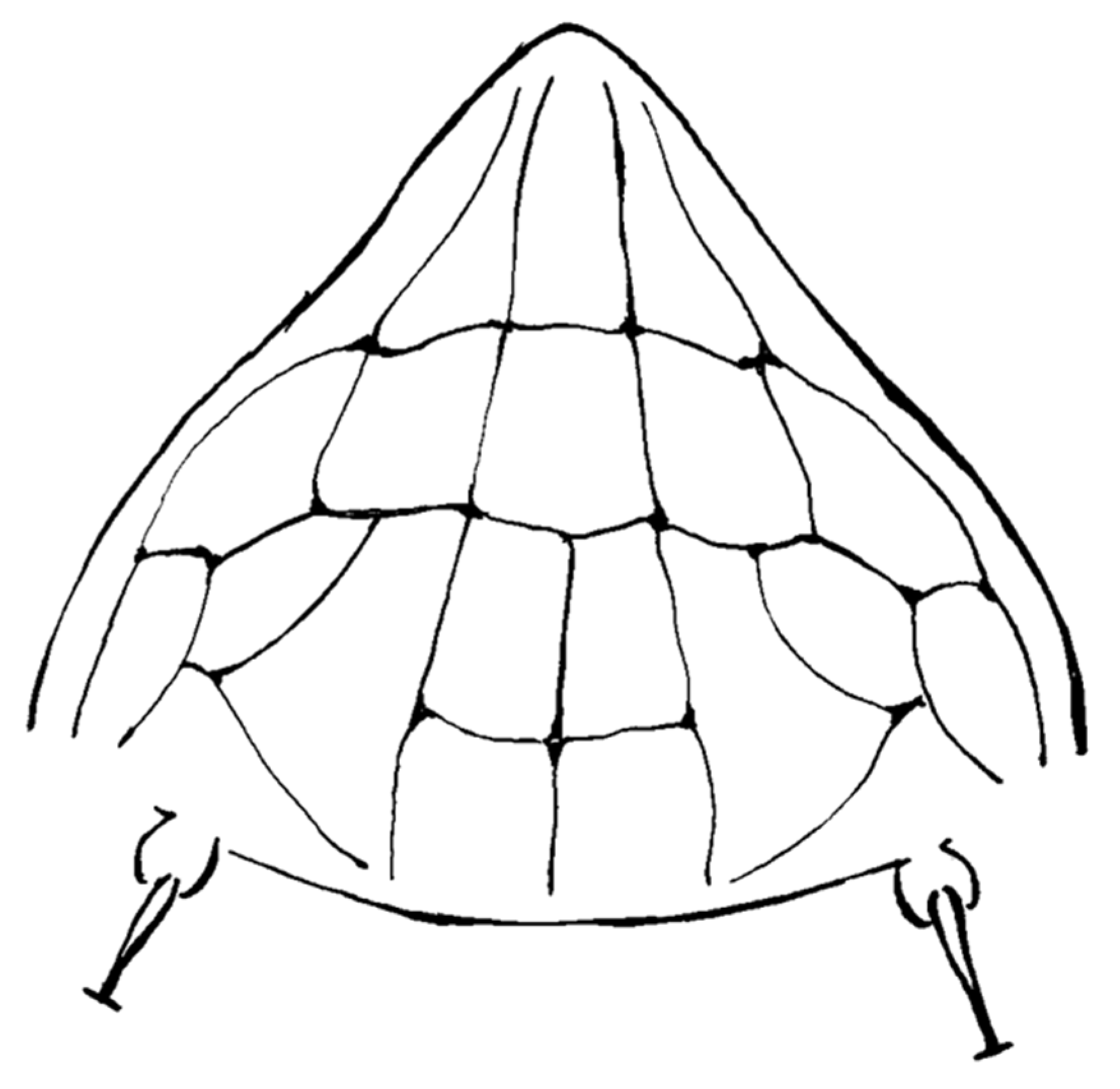
A detailed morphological study of Ac. taihangensis was not possible owing to the few specimens comprising the ethanol’s preserved paratypes, as well as the strong tissue fixation induced by ethanol, which were unsuitable for a tissue digestion. However, it was possible to check the morphology of the prodorsal shield of a single female paratype, and the anterior lobe resulted to be rounded (Figure 4). The pattern of the prodorsal shield is characterized by a short median line on the rear half of the shield, a complete pair of admedian lines, two transverse lines joining the median with the admedian lines on the rear half of the shield forming four cells in the middle field of the shield, and a transverse line joining the admedian lines on the anterior one third of the shield forming two median cells; this transverse line continues in a pair of outer and arched submedian lines contributing to the delimitation of further cells between the submedian lines and the anterior-lateral edge of the shield (Figure 4). The original description of Ac. taihangensis indicates the presence of a complete median line [37], but a weak or obscure median line has also been observed in the anterior half of the shield in some specimens of the currently studied population of A. mosoniensis.
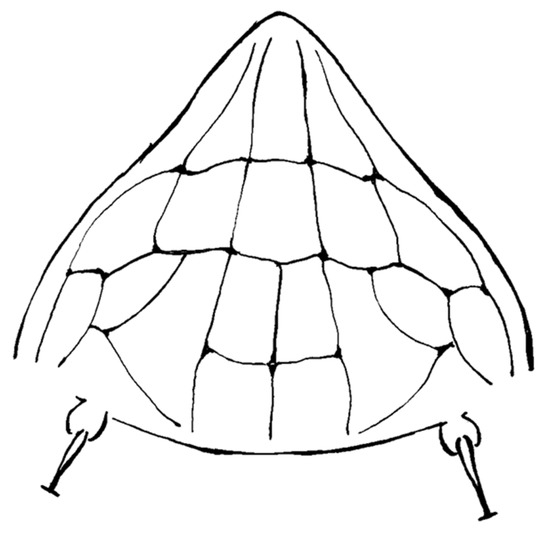
Figure 4.
Line drawing of the prodorsal shield of Aculops taihangensis paratype.
3.3. Molecular and Phylogenetic Analyses
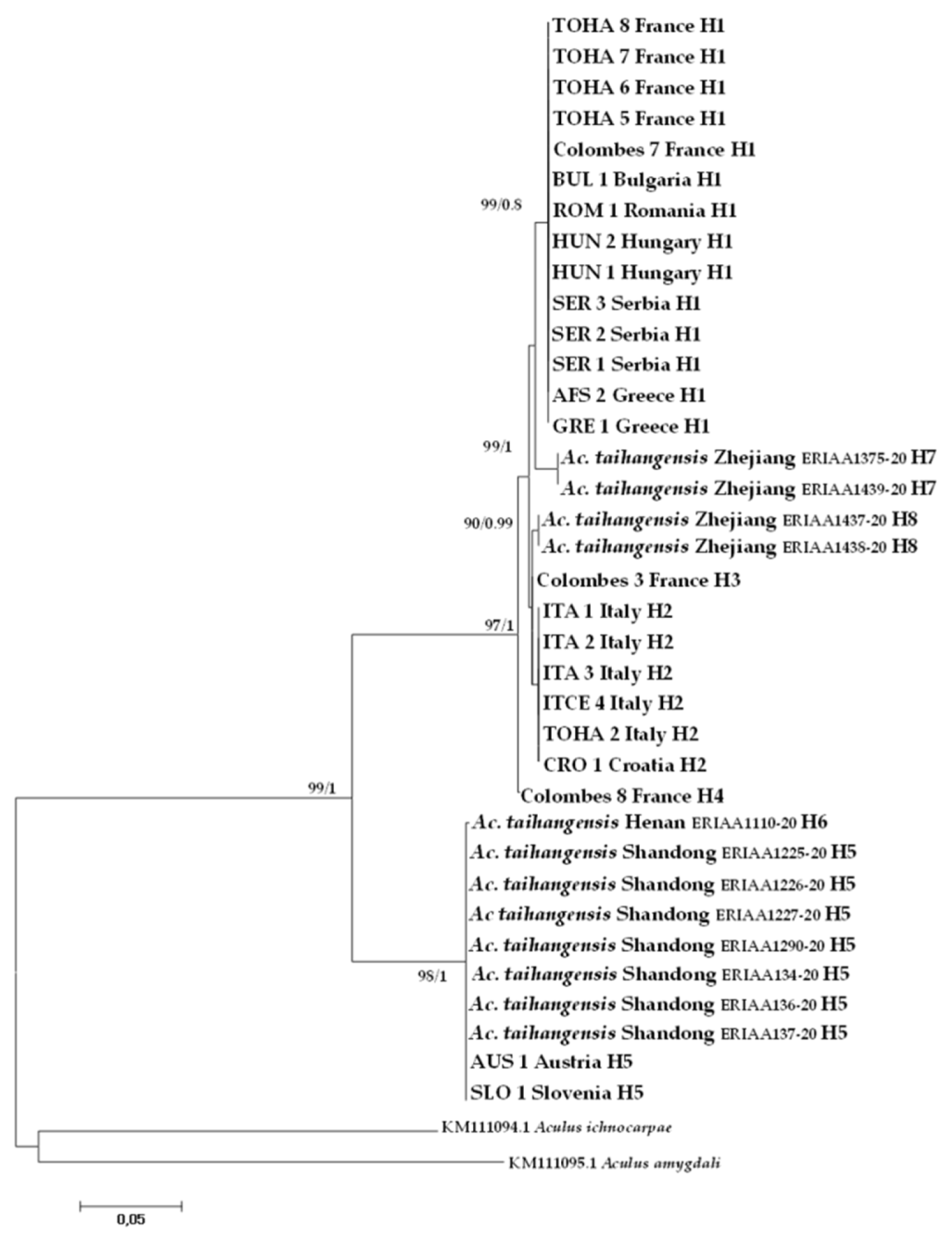
Barcode-compliant sequences of 658 nt in length were generated from all studied specimens (Table 1). A total of eight haplotypes were recovered from the CO1 dataset combining A. mosoniensis and Ac. taihangensis sequences and the most common haplotype H1 was shared between six European countries (Figure 5). The haplotype H5 was harbored both by Ac. taihangensis from the Shandong province and A. mosoniensis mites from Austria and Slovenia. The pairwise genetic p-distance between A. mosoniensis and Ac. taihangensis averaged 7.68%, which is less than half the pairwise distances (23.03% and 24.78%) estimated between A. mosoniensis, A. ichnocarpae, and A. amygdali, respectively (Table 2). The phylogenetic reconstruction obtained under maximum likelihood and in a Bayesian framework (tree not shown) with A. ichnocarpae and A. amygdali as outgroups (Figure 5) demonstrated that all A. mosoniensis grouped with Ac. taihangensis in one fully supported clade with 99% support. It also demonstrated that this clade split into two major maternal lineages with high bootstrap support (97 to 98%), one including Ac. taihangensis from Shandong and Henan provinces and A. mosoniensis from Austria and Slovenia, the other clustering all the other European A. mosoniensis with Ac. taihangensis from the Zhejiang Province.
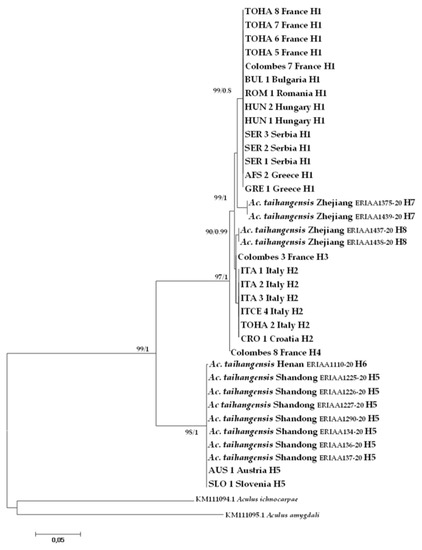
Figure 5.
Maximum likelihood phylogenetic tree of CO1 sequences of Aculus mosoniensis, Aculops taihangensis, and two congeners, Aculus ichnocarpae and Aculus amygdali, used as outgroups. Only significant values of bootstrap support (>90%) and of Bayesian posterior probabilities obtained by MrBayes (>0.8) are shown at the nodes. Branch lengths represent expected substitutions per site. The scale bar indicates the expected number of substitutions per site. Tip labels refer to specimen IDs and haplotypes listed in Table 1.
Table 2.
CO1 pairwise distance matrix with uncorrected p distances shown as percentages (above the diagonal) and K2P distances (under the diagonal) between Aculus mosoniensis, Aculops taihangensis, and two congeners (Aculus ichnocarpae and Aculus amygdali), and within Aculus mosoniensis, and Aculops taihangensis (in the diagonal). Data are expressed as mean in % ± S.E.
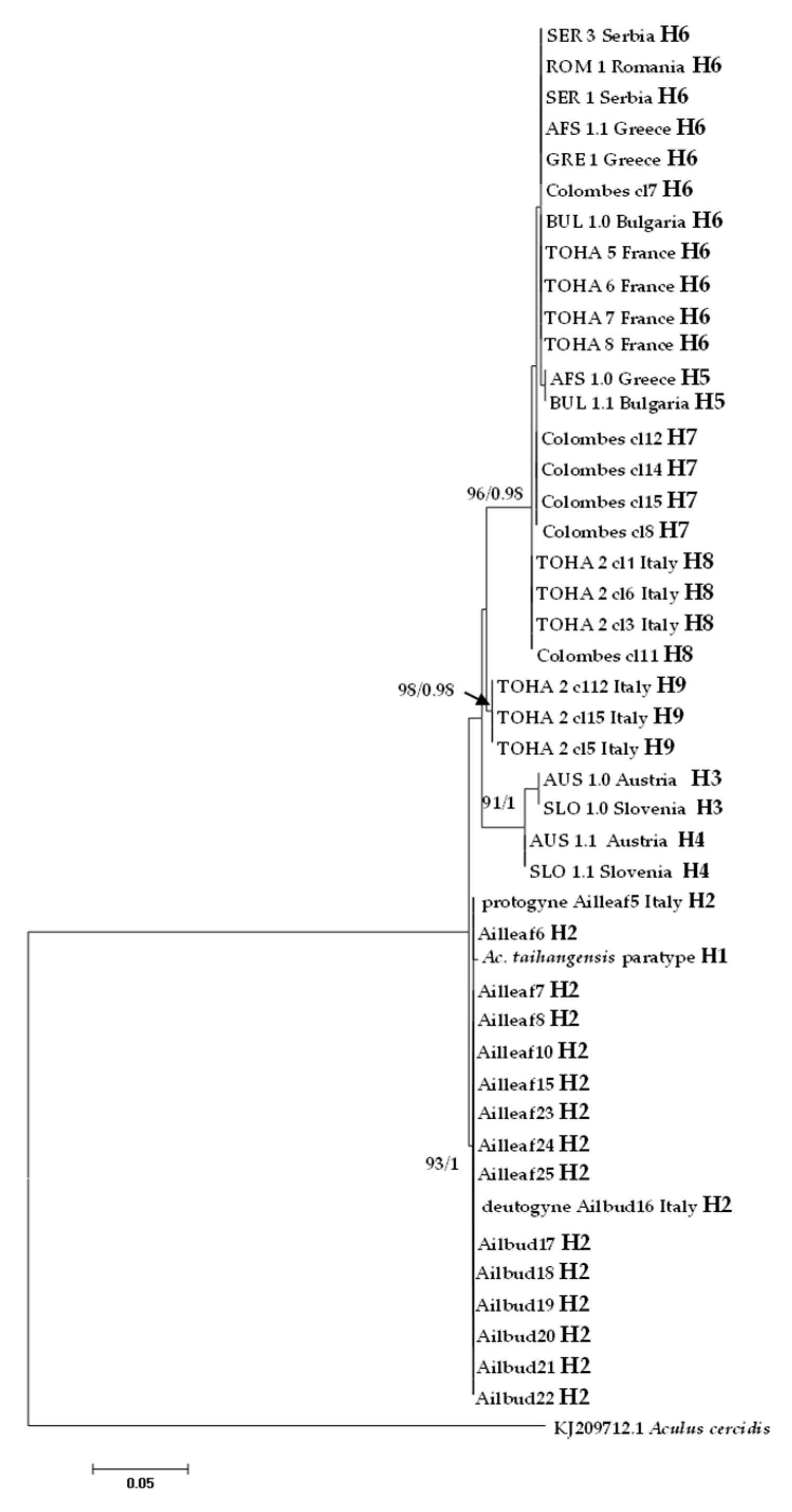
In the ITS1 alignment generated from all analyzed specimens, due to the presence of indels, sequences varied in length from 440 bp (Colombes in France) to 449 bp (Austria and Slovenia), while ITS1 was 447 bp long in Ac. taihangensis paratype (Table 1). A total of nine haplotypes were recovered and the most common haplotype H6 was shared between five European countries (Figure 6).
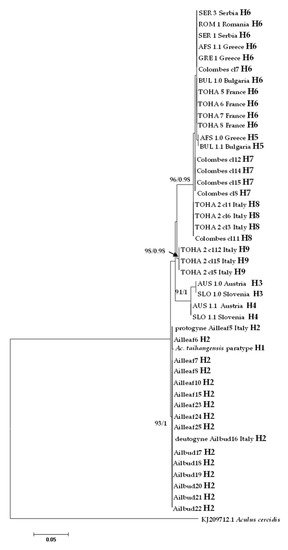
Figure 6.
Maximum likelihood phylogenetic tree of Aculus mosoniensis ITS1 sequences, Aculops taihangensis, and one congener, Aculus cercidis, used as outgroup. Only significant values of bootstrap support (>90%) and of Bayesian posterior probabilities obtained by MrBayes (>0.9) are shown at the nodes. The scale bar indicates the expected number of substitutions per site. Tips labels refer to specimen IDs and haplotypes listed in Table 1.
All mites collected from leaves and buds at Bari (Italy) harbored the same ITS1 haplotype H2 which was shown to be the most closely related to H1 found in the Chinese Ac. taihangensis specimens, with a pairwise p-distance of 0.24%. The pairwise p-distances between all A. mosoniensis and Ac. taihangensis averaged 2.26%, which is almost sixteen times less than the pairwise distance (37.10%) estimated between A. mosoniensis and A. cercidis (Table 3). The phylogenetic reconstruction obtained in a maximum likelihood framework as well as in a Bayesian framework (tree not shown) with A. cercidis as an outgroup (Figure 6) demonstrated that most leaf nodes of the tree were highly supported, contrary to internal nodes. All A. mosoniensis collected in Bari (Italy) grouped with Ac. taihangensis in one fully supported clade with 93% support. Three other clades with high bootstrap support (91 to 98%) were evidenced: one gathering H3 and H4 from Austria and Slovenia, one with haplotype H9 found in Rome (Italy), and the last ones clustering four haplotypes (H5, H6, H7, and H8) widely distributed in Europe.
Table 3.
ITS1 pairwise distance matrix with uncorrected p distances shown as percentages (above the diagonal) and K2P distances (under the diagonal) within Aculus mosoniensis; between Aculus mosoniensis, Aculops taihangensis, and one congener (Aculus cercidis); and within Aculus mosoniensis (in the diagonal). Data are expressed as mean ± S.E.
4. Discussion and Conclusions
The examination of the anterior prodorsal shield lobe of an Ac. taihangensis paratype allowed assigning this species to the genus Aculus, as previously suspected based on the original description [7]. A different procedure applied in digesting and mounting mites allowed us to revise the description of its prodorsal shield pattern which appears to be consistent with that of A. mosoniensis. All the other traits were already stated to be very similar between the two species [7]. The description of a deutogyne morph, as found in the current study, has also been added to the original description.
The two molecular markers applied in the present study resulted as congruent with the morphological examination and supported the similarity between the two species names. The CO1 sequences of Ac. taihangensis from the Shandong province in China were identical to that of A. mosoniensis specimens collected in Austria and Slovenia, and the pairwise genetic p-distance between Ac. taihangensis and A. mosoniensis averaged 7.68%, which is much higher than the average intraspecific divergence (1.15%) in the Eriophyidae [29] but twofold less than estimates of intraspecific variation in other eriophyid mites such as Aceria tosichella Keifer (14.5%, e.g., [15]). Moreover, this estimate was also less than half the genetic distances between Aculus species (23–24.78%) estimated in the present study, in the Eriophyidae (19.86% [29]), and between congeneric species in other genera, such as Aculodes (20.2–21.5% [13]) or Aceria (15.6–22.0% [15]). In addition, the ITS1 sequence of Ac. taihangensis was found nearly identical (99.8%) to the ITS1 of A. mosoniensis specimens collected in Bari, Italy, and the pairwise genetic p-distance between Ac. taihangensis and A. mosoniensis averaged 2.6%, which was one order of magnitude less than the divergence between A. mosoniensis and the outgroup A. cercidis (37.10%). This latter estimate was surprisingly higher than the interspecies divergence reported for nuclear markers in other eriophyids, which ranged from 2% in Aceria [15] to 17% in Abacarus [38], but rather comparable to between Abacarus and Aceria [38]. A more accurate estimation of the range of divergence between Aculus species would require further investigation by increasing the ITS1 dataset of this genus as there is only one NCBI publicly available ITS1 sequence. Interestingly, in our analysis, A. mosoniensis was shown to exhibit some genetic polymorphism by uncovering five CO1 in Europe (becoming eight along with those of Ac. taihangensis) and clustered in two major maternal lineages that were consistent but not entirely identical to the four clades/subclades described by the ITS1. The maternal lineage (Austria and Slovenia) and the one grouping most of the European populations were also evidenced in the ITS1 phylogenetic reconstruction. The large distribution of the latter clade suggested that most European countries have been colonized by mostly one CO1 haplotype (H1) and one ITS1 haplotype (H6). Results also evidenced that the two European A. mosoniensis lineages were closely related to different Ac. taihangensis haplotypes found in two geographically distant provinces in China, i.e., Zhejiang and Shandon–Henan, suggesting two different origins in China of the European A. mosoniensis.
Based on the new morphological and molecular data, A. mosoniensis has to be considered a junior synonym of Aculus taihangensis new combination, which also means that the distribution range of A. taihangensis is not only restricted to China but also includes Europe.
Nevertheless, the eriophyid fauna associated with ToH still require further investigations. In fact, the prodorsal shield of A. taihangensis new. comb. appears to match that of Ac. ailanthi, as recently documented by Skrvarla et al. [10]. Unfortunately, the original morphological description of the latter species [39] is not consistent with the usual standard and shows some differences with A. taihangensis new. comb. [11]. Hence, it requires a better morphological characterization of the types in order to provide more careful details of the prodorsal shield pattern, genital, and coxal region, and morphometric data as well. Similarly, no molecular marker of this species has been currently known. Clarifying the taxonomic status of these two species is particularly important in the perspective of a biological control approach against ToH, especially because recent studies have shown a relevant impact of Ac. ailanthi on the ToH, as well as some similarities with A. taihangensis new. comb. concerning the host plant relationships [10,12]. In addition, deep investigations are needed to verify the real impact of the different haplotypes on the ToH.
Partial descriptions of new species, often out of standard, are confusing. The proper taxonomic identification of the natural enemies associated with a weed is a binding step of a weed biological control program. Due to some of their attributes, such as high host specificity and potentially high impact on their host, the general interest in the use of eriophyid mites in biological control of weeds is increasing [6]. It is precisely in this perspective that the current study represents an important step forward in the process of clarifying the eriophyid fauna associated with a highly invasive species such as ToH.
Author Contributions
Conceptualization, E.d.L., M.C. and R.P.; methodology, B.V., E.d.L. and M.C.; validation, M.-C.B. and T.C.; formal analysis, M.-C.B. and T.C.; investigation, B.V., D.V., E.d.L., F.M., M.C. and M.-C.B.; resources, M.-C.B. and T.C.; data curation, M.-C.B. and T.C.; writing—original draft preparation, E.d.L. and M.-C.B.; writing—review and editing, E.d.L., F.M. and M.C.; funding acquisition, B.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was in part supported by: Ministry of Education, Science and Technological Development of the Republic of Serbia, Grant No. 451-03-68/2022-14/200116; Serbian Academy of Sciences and Arts, Grant No. F-195.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article, whereas the new sequences have been deposited in the Genbank repository.
Acknowledgments
The authors are really grateful to Xiao-Yue Hong and Xiao-Feng Xue (Department of Entomology, Nanjing Agricultural University, Nanjing, Jiangsu, China) for providing type material and for the useful information regarding molecular analysis of Chinese specimens, to James W. Amrine (University of Morgantown, Morgantown, WV, USA) and Lincoln Smith (USDA-ARS Western Regional Research Center, Albany, CA, USA) for their careful review of a draft of this paper, to Fatiha Guermache (EBCL, France) for technical support in molecular biology, to Javid Kashefi (EBCL, France) for collecting mites in France, Greece, Bulgaria, to Pasquale Trotti (DiSSPA, University of Bari Aldo Moro, Italy) for technical support in scanning electron microscopy, and to Dragica Smiljanić (UB, Serbia) for collecting mites in Serbia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nooteboom, H.P. Ailanthus. In Flora Malesiana (Ser I, 6); Noordhoff International publishing: Leyden, The Netherlands, 1962; pp. 215–222. [Google Scholar]
- Burch, P.L.; Zedaker, S.M. Removing the Invasive Tree Ailanthus altissima and Restoring Natural Cover. J. Arboricult. 2003, 29, 18–24. [Google Scholar] [CrossRef]
- Heisey, R.M. Identification of an allelopathic compound from Ailanthus altissima (Simaroubaceae) and characterization of its herbicidal activity. Am. J. Bot. 1996, 83, 192–200. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Per. Pl. Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Krussmann, G.; Wennemuth, G.; Thon, H.E. Die baumschule, 5th ed.; Parey: Berlin, Germany, 1981; p. 565. [Google Scholar]
- Marini, F.; Profeta, E.; Vidović, B.; Petanović, R.; de Lillo, E.; Weyl, P.; Hinz, H.L.; Moffat, C.E.; Bon, M.C.; Cvrković, T.; et al. Field Assessment of the Host Range of Aculus mosoniensis (Acari: Eriophyidae), a Biological Control Agent of the Tree of Heaven (Ailanthus Altissima). Insects 2021, 12, 637. [Google Scholar] [CrossRef]
- de Lillo, E.; Panzarino, O.; Loverre, P.; Valenzano, D.; Mattia, C.; Marini, F.; Augé, M.; Cristofaro, M. New eriophyoid mites from Italy. IV. Mites associated with weed plants. Syst. Appl. Acarol. 2017, 22, 2256–2272. [Google Scholar] [CrossRef]
- Ripka, G.; Érsek, L. A new Aculops species (Acari: Prostigmata: Eriophyoidea) on Ailanthus altissima from Hungary. Acta Phytopathol. Entomol. Hung. 2014, 49, 49–56. [Google Scholar] [CrossRef]
- Gardener, R.T. Native and Indigenous Biocontrols for Ailanthus Altissima. Master’s Thesis, University of Maryland, College Park, MD, USA, 2008. [Google Scholar]
- Skvarla, M.J.; Ochoa, R.; Ulsamer, A.; Amrine, J. The eriophyid mite Aculops ailanthi Lin, Jin, & Kuang, 1997 (Acariformes: Prostigmata: Eriophyidae) from tree-of-heaven in the United States-new state records and morphological observations. Acarologia 2021, 61, 121–127. [Google Scholar]
- de Lillo, E.; Craemer, C.; Amrine, J.W., Jr.; Nuzzaci, G. Recommended procedures and techniques for morphological studies of Eriophyoidea (Acari: Prostigmata). Exp. Appl. Acarol. 2010, 51, 283–307. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- de Lillo, E.; Vidović, B.; Petanović, R.; Cristofaro, M.; Marini, F.; Augé, M.; Cvrković, T.; Babić, E.; Mattia, C.; Lotfollahi, P.; et al. A new Aculodes species (Prostigmata: Eriophyoidea: Eriophyidae) associated with medusahead, Taeniatherum caput-medusae (L.) Nevski (Poaceae). Syst. Appl. Acarol. 2018, 23, 1217. [Google Scholar] [CrossRef] [Green Version]
- de Lillo, E.; Fanelli, E.; Valenzano, D.; Monfreda, R.; Troccoli, A.; Vovlas, A.; De Luca, F. Characterization of Aceria massalongoi and a histopathology study of the leaf galls induced on chaste tree. Exp. Appl. Acarol. 2020, 82, 33–57. [Google Scholar] [CrossRef]
- Skoracka, A.; Kuczyński, L.; Szydło, W.; Rector, B. The wheat curl mite Aceria tosichella (Acari: Eriophyoidea) is a complex of cryptic lineages with divergent host ranges: Evidence from molecular and plant bioassay data. Biol. J. Linn. Soc. 2013, 109, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Manson, D.C.M.; Oldfield, G. Life forms, deutogyny, diapause and seasonal development. In Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests 6; Elsevier Science Publishers: Amsterdam, The Netherlands, 1996; pp. 173–183. [Google Scholar]
- Valenzano, D.; Tumminello, M.T.; Gualandri, V.; de Lillo, E. Morphological and molecular characterization of the Colomerus vitis erineum strain (Trombidiformes: Eriophyidae) from grapevine erinea and buds. Exp. Appl. Acarol. 2020, 80, 183–201. [Google Scholar] [CrossRef]
- Baker, E.W.; Kono, T.; Amrine, J.W., Jr.; Delfinado-Baker, M.; Stasny, T.A. Eriophyoid mites of the United States; Indira Publishing House: West Bloomfield, MI, USA, 1996. [Google Scholar]
- Keifer, H.H. Eriophyoidea Nalepa. Injurious eriophyoid mites. In Mites Injurious to Economic Plants; Jeppson, L.R., Keifer, H.H., Baker, E.W., Eds.; University California Press: Berkeley, CA, USA, 1975; pp. 327–533. [Google Scholar]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests 6; Elsevier Science Publishers: Amsterdam, The Netherlands, 1996; pp. 3–31. [Google Scholar]
- Amrine, J.W., Jr.; Manson, D.C.M. Preparation, mounting and descriptive study of Eriophyoid mites. In Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests 6; Elsevier Science Publishers: Amsterdam, The Netherlands, 1996; pp. 383–396. [Google Scholar]
- Amrine, J.W., Jr.; Stasny, T.A.; Flechtmann, C.H.W. Revised Keys to World Genera of Eriophyoidea (Acari: Prostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2003. [Google Scholar]
- Dabert, J.; Ehrnsberger, R.; Dabert, M. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): Compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa 2008, 1719, 41–52. [Google Scholar]
- Kashefi, J.; Vidović, B.; Guermache, F.; Cristofaro, M.; Bon, M.C. Occurrence of Aculus mosoniensis (Ripka 2014) (Acari: Prostigmata: Eriophyoidea) on tree of heaven (Ailanthus altissima Mill.) is expanding across Europe: First records in France confirmed by barcode. Phytoparasitica 2021, 49, 49–51. [Google Scholar] [CrossRef]
- Navajas, M.; Lagnel, J.; Fauvel, G.; De Moraes, G. Sequence variation of ribosomal internal transcribed spacers (ITS) in commercially important phytoseiidae mites. Exp. Appl. Acarol. 1999, 23, 851–859. [Google Scholar] [CrossRef]
- Carew, M.E.; Goodisman, M.A.D.; Hoffmann, A.A. Species status and population genetic structure of grapevine eriophyoid mites. Entomol. Exp. Appl. 2004, 111, 87–96. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Yin, Y.; Yao, L.-F.; Hu, Y.; Shao, Z.-K.; Hong, X.-Y.; Hebert, P.D.N.; Xue, X.-F. DNA barcoding uncovers cryptic diversity in minute herbivorous mites (Acari, Eriophyoidea). Mol. Ecol. Res. 2022, 00, 1–13. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. & Evol. 2021, 38, 3022–3027. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Srivathsan, A.; Meier, R. On the inappropriate use of Kimura-2-parameter (K2P) divergences in the DNA-barcoding literature. Cladistics 2012, 28, 190–194. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Hong, X.-Y.; Xue, X.-F. Four new species of Aculops Keifer (Acari: Eriophyidae) from China. Oriental Ins. 2005, 39, 203–211. [Google Scholar] [CrossRef]
- Duarte, M.E.; De Mendonça, R.S.; Skoracka, A.; Silva, E.S.; Navia, D. Integrative taxonomy of Abacarus mites (Eriophyidae) associated with hybrid sugarcane plants, including description of a new species. Exp. Appl. Acarol. 2019, 78, 373–401. [Google Scholar] [CrossRef]
- Lin, F.P.; Jin, C.; Kuang, H.Y. A new genus and six new species of Eriophyidae from China (Acari: Eriophyoidea). Acta Entomol. Sin. 1997, 40, 86–93. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).