
Effects of Juvenile Hormone Analog and Days after Emergence on the Reproduction of Oriental Armyworm, Mythimna separata (Lepidoptera: Noctuidae) Populations


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. JHA Treatment
2.3. Adult Age Treatment
2.4. Reproductive Parameter Determination
2.5. Data Analysis
3. Results
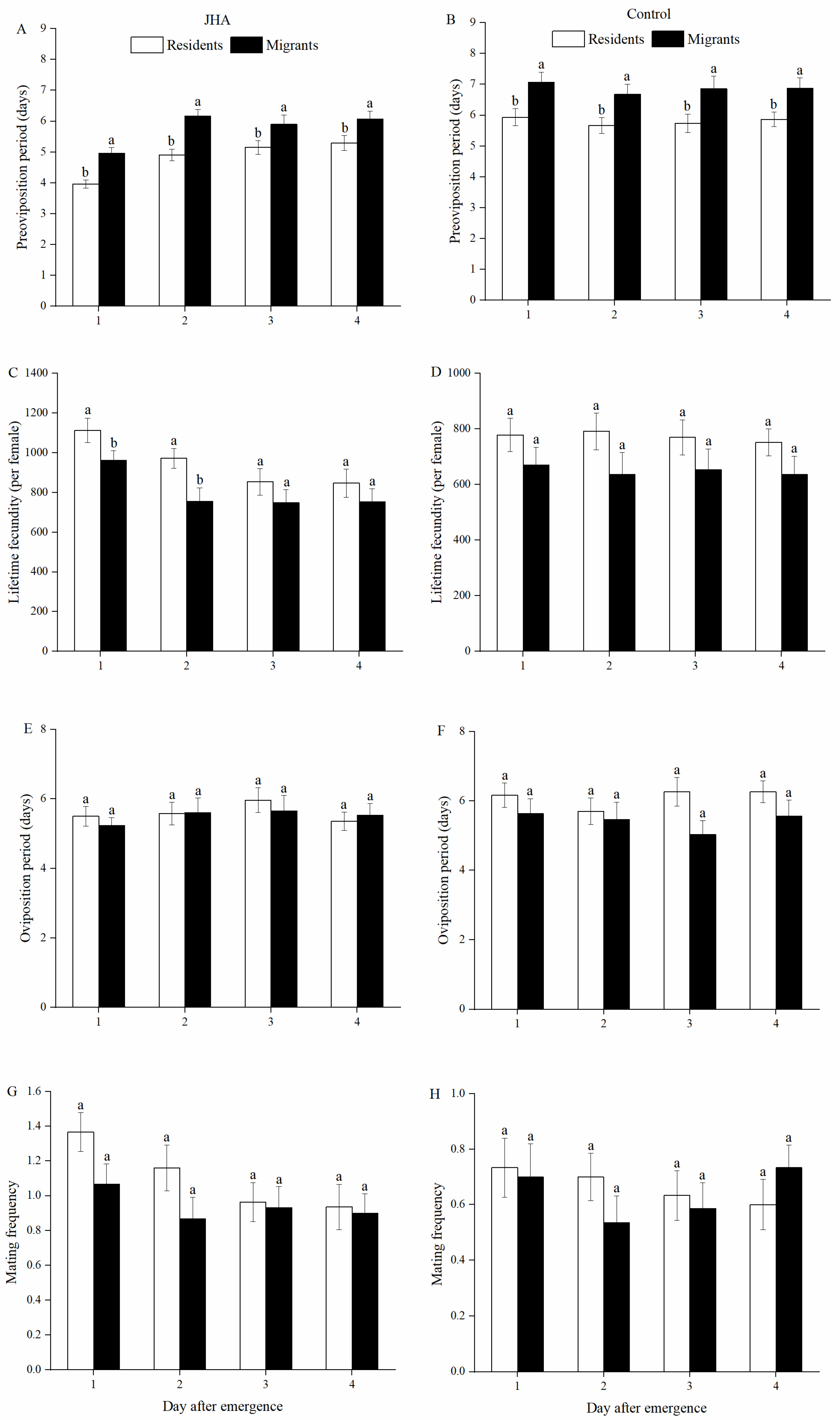
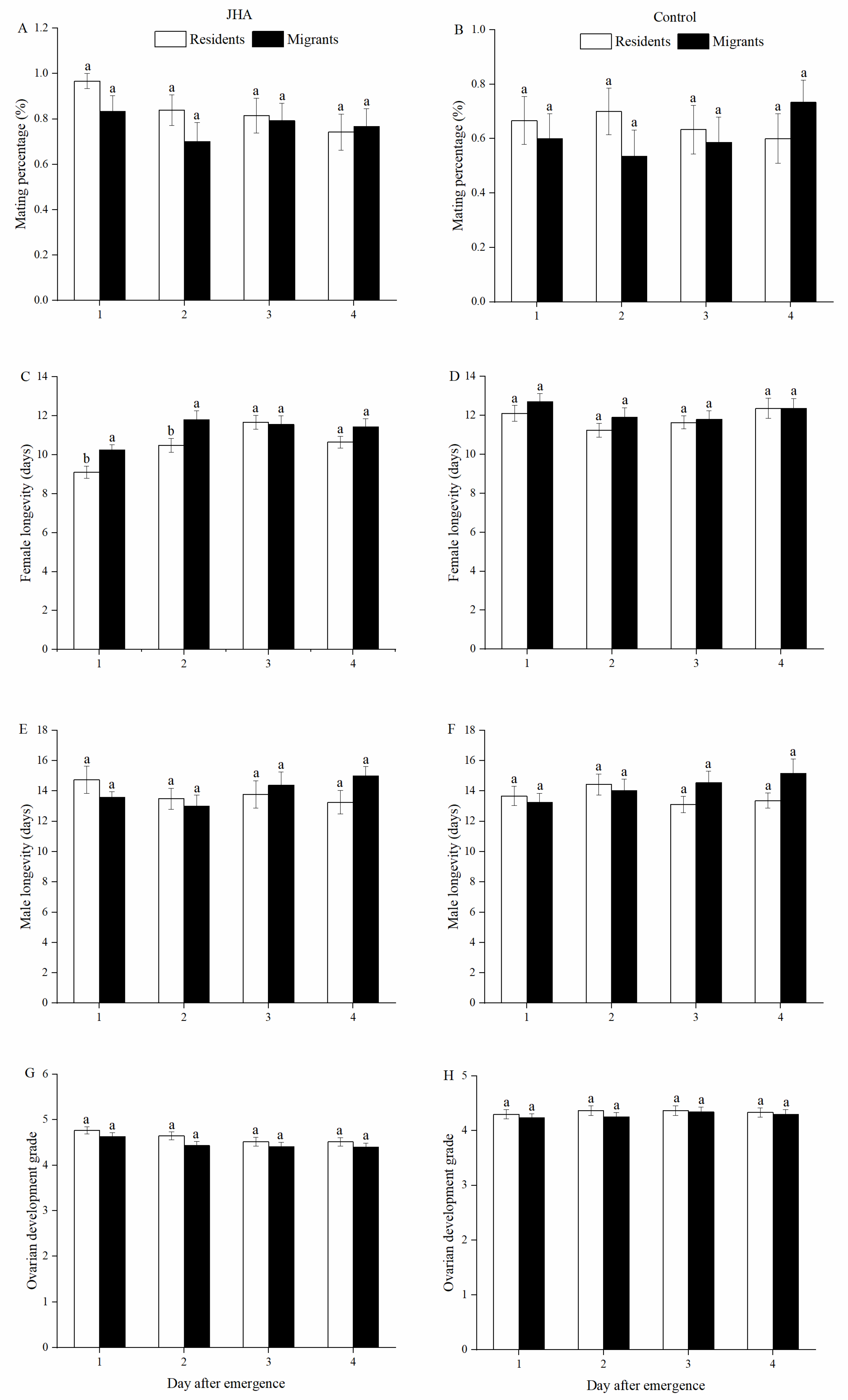
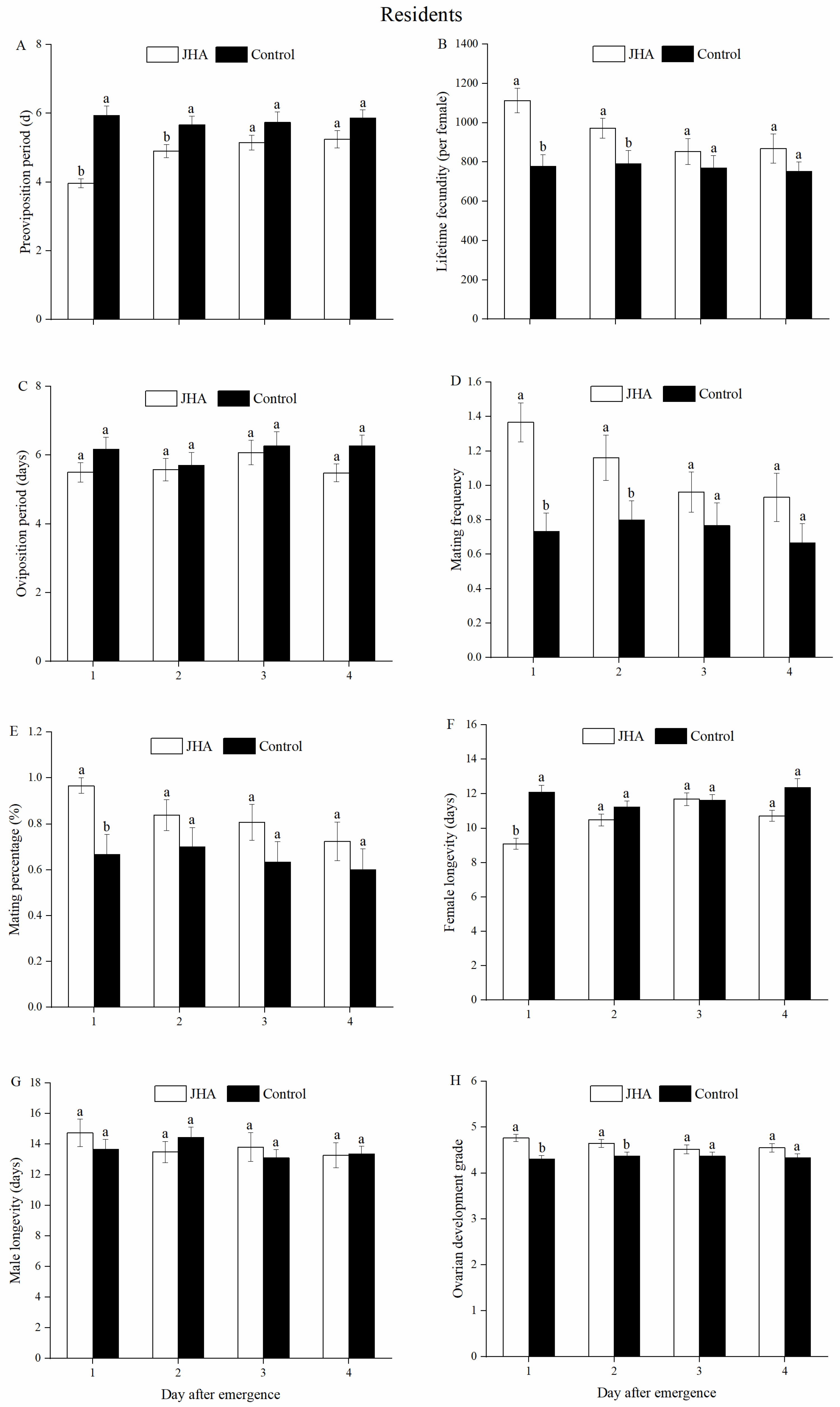
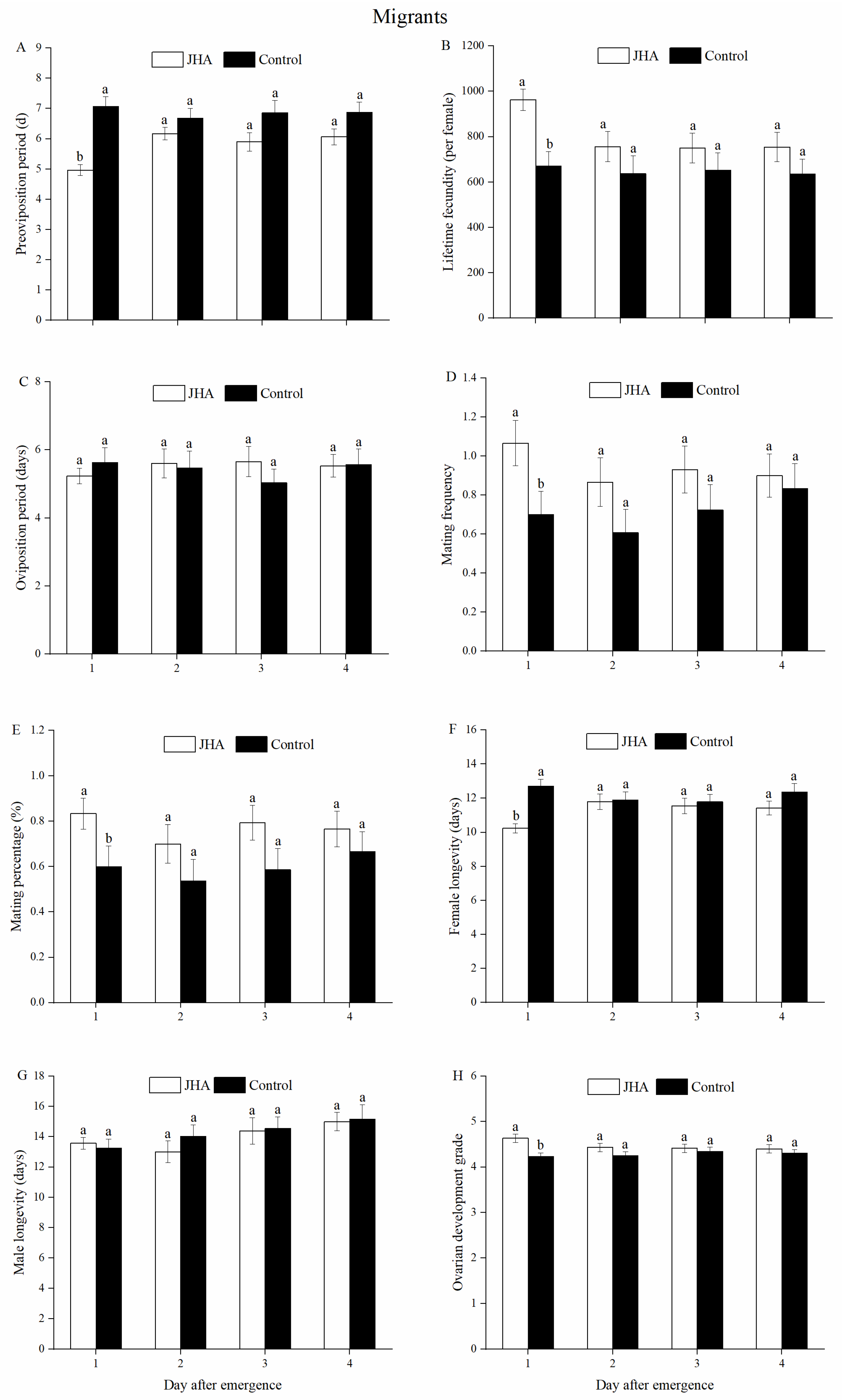
3.1. Reproductive Performance of M. separata between Residents and Migrants Exposed to JHA at Various Ages
3.2. Effect of JHA on Reproductive Performance of M. separata Populations at Various Ages
3.3. Effect of Age on Reproductive Performances of M. separata Populations Treated by JHA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baumann, A.; Fujiwara, Y.; Wilson, T.G. Evolutionary divergence of the paralogs methoprene tolerant (Met) and germ cell expressed (gce) within the genus Drosophila. J. Insect Physiol. 2010, 56, 1445–1455. [Google Scholar] [CrossRef]
- Bilen, J.; Atallah, J.; Azanchi, R.; Levine, J.D.; Riddiford, L.M. Regulation of onset of female mating and sex pheromone production by juvenile hormone in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2013, 110, 18321–18326. [Google Scholar] [CrossRef]
- Jindra, M. Something “hairy” in juvenile hormone signaling for mosquito reproduction. Proc. Natl. Acad. Sci. USA 2016, 113, 1474–1476. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.G. Migration and Dispersal of Insects by Flight; Methuen: London, UK, 1969. [Google Scholar]
- Dingie, H. Migration: The Biology of Life on the Move; Oxford University Press: New York, NY, USA, 1996. [Google Scholar]
- Lorenz, M.W. Oogenesis-flight syndrome in crickets: Age-dependent egg production, flight performance, and biochemical composition of the flight muscles in adult female Gryllus bimaculatus. J. Insect Physiol. 2007, 53, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Zera, A.J. Endocrine analysis in evolutionary-developmental studies of insect polymorphism: Hormone manipulation versus direct measurement of hormonal regulators. Evol. Dev. 2007, 9, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wu, Z.; Wang, Z.; Deng, S.; Zhou, S. Krüppel-homolog 1 mediates juvenile hormone action to promote vitellogenesis and oocyte maturation in the migratory locust. Insect Biochem. Mol. Biol. 2014, 52, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Harano, K.I.; Sasaki, K.; Nagao, T.; Sasaki, M. Influence of age and juvenile hormone on brain dopamine level in male honeybee (Apis mellifera): Association with reproductive maturation. J. Insect Physiol. 2008, 54, 848–853. [Google Scholar] [CrossRef]
- Chapman, J.W.; Reynolds, D.R.; Wilson, K. Long-range seasonal migration in insects: Mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015, 18, 287–302. [Google Scholar] [CrossRef]
- Jiang, X.F.; Luo, L.Z.; Sappington, T.W. Relationship of flight and reproduction in beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae), a migrant lacking the oogenesis-flight syndrome. J. Insect Physiol. 2010, 56, 1631–1637. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Luo, L.Z.; Jiang, X.F.; Sappington, T.W. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm. PLoS ONE 2012, 7, e31562. [Google Scholar] [CrossRef]
- Luo, L.Z.; Jiang, X.F.; Li, K.B.; Hu, Y. Influences of flight on reproduction and longevity of the oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 1999, 42, 150–158. [Google Scholar]
- Cheng, Y.X.; Luo, L.Z.; Sappington, T.W.; Jiang, X.F.; Zhang, L.; Frolov, A.N. Onset of oviposition triggers abrupt reduction in migratory flight behavior and flight muscle in the female beet webworm, Loxostege sticticalis. PLoS ONE 2016, 11, e0166859. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.W.; Li, P.; Zhang, J.; Liu, X.D.; Zhai, B.P.; Hu, G. Cnaphalocrocis medinalis moths decide to migrate when suffering nutrient shortage on the first day after emergence. Insects 2019, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Luo, L.Z.; Jiang, X.F. Starvation influences allatotropin gene expression and juvenile hormone titer in the female adult oriental armyworm, Mythimna separata. Arch. Insect Biochem. Physiol. 2008, 68, 63–70. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Sappington, T.W.; Luo, L.Z.; Zhang, L.; Jiang, X.F. Starvation on first or second day of adulthood reverses larval-stage decision to migrate in beet webworm (lepidoptera: Pyralidae). Environ. Entomol. 2021, 50, 523–531. [Google Scholar] [CrossRef]
- Lv, W.X.; Xie, X.C. Effect of fluctuating temperatures on development, reproduction and energy of oriental armyworm populations, Mythimna separata. J. Appl. Entomol. 2022, 146, 511–524. [Google Scholar] [CrossRef]
- Mittler, T.E. Juvenile Hormone and Aphid Polymorphism; Rutgers University Press: New Brunswick, NJ, USA, 1991. [Google Scholar]
- Zera, A.J.; Denno, R.F. Physiology and ecology of dispersal polymorphism in insects. Annu. Rev. Entomol. 1997, 42, 207–230. [Google Scholar] [CrossRef]
- Du, B.; Ding, D.; Ma, C.; Guo, W.; Kang, L. Locust density shapes energy metabolism and oxidative stress resulting in divergence of flight traits. Proc. Natl. Acad. Sci. USA 2022, 119, e2115753118. [Google Scholar] [CrossRef]
- Wang, G.P.; Zhang, Q.W.; Ye, Z.H.; Luo, L.Z. The role of nectar plants in the severe outbreaks of armyworm, Mythimna separata (Lepidoptera: Noctuidae) in China. Bull. Entomol. Res. 2006, 96, 445–455. [Google Scholar]
- Chen, R.L.; Sun, Y.J.; Wang, S.Y.; Zhai, B.P.; Bao, X.Z. Migration of the Oriental Armyworm, Mythimna separata in East Asia in Relation to Weather and Climate. I. Northeastern China; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- He, Y.Q.; Feng, B.; Guo, Q.S.; Du, Y.J. Age influences the olfactory profiles of the migratory oriental armyworm Mythimna separata at the molecular level. BMC Genom. 2017, 18, 32. [Google Scholar] [CrossRef][Green Version]
- Wang, C.L.; Zhang, L.; Lv, W.X. Flying males mediate oviposition and migration in female Mythimna separata (Lepidoptera: Noctuidae). Bull. Entomolog. Res. 2022, 112, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.F.; Luo, Z.L.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of migration in the oriental armyworm, Mythimna separata (Walker) in China: A review integrating environmental, physiological, hormonal, genetic, and molecular factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cheng, L.L.; Chapman, J.W.; Sappington, T.W.; Liu, J.J.; Cheng, Y.X.; Jiang, X.F. Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 2020, 10, 11626. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.F.; Luo, L.Z. Comparison of behavioral and physiological characteristics between the emigrant and immigrant populations of the oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 2005, 48, 61–67. [Google Scholar]
- Luo, L.Z.; Li, K.B.; Jiang, X.F.; Hu, Y. Regulation of flight capacity and contents of energy substances by methoprene in the moths of oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 2001, 8, 63–72. [Google Scholar]
- Zhang, L.; Jiang, X.F.; Luo, L.Z. Determination of sensitive stage for switching migrant oriental armyworms into residents. Environ. Entomol. 2008, 37, 1389–1395. [Google Scholar] [CrossRef]
- Wang, F.; Lv., W.X. Low temperature triggers physiological and behavioral shifts in adult oriental armyworm, Mythimna separata. Bull. Entomolog. Res. 2022, in press. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Y.D.; Qi, X.H.; Xu, Y.W.; Hou, Y.H.; Fan, Z.Y.; Shen, H.L.; Liu, D.; Shi, X.K.; Li, S.M.; et al. The effects of climate warming on the migratory status of early summer populations of Mythimna separata (Walker) moths: A case–study of enhanced corn damage in central-northern China, 1980–2016. Ecol. Evol. 2019, 9, 12332–12338. [Google Scholar] [CrossRef]
- Marshall, K.E.; Sinclair, B.J. Repeated stress exposure results in a survival reproduction trade-off in Drosophila melanogaster. Proc. Biol. Sci. 2010, 277, 963–969. [Google Scholar]
- Lv, W.X.; Jiang, X.F.; Chen, X.J.; Cheng, Y.X.; Xia, J.X.; Zhang, L. Flight and reproduction variations of rice leaf roller, Cnaphalocrocis medinalis in response to different rearing temperatures. Insects 2021, 12, 1083. [Google Scholar] [CrossRef]
- Zhao, X.C.; Feng, H.Q.; Wu, B.; Wu, X.F.; Liu, Z.F.; Wu, K.M.; McNeil, J.N. Does the onset of sexual maturation terminate the expression of migratory behaviour in moths? A study of the oriental armyworm, Mythimna separata. J. Insect Physiol. 2009, 55, 1039–1043. [Google Scholar] [CrossRef]
- Guo, J.W.; Cui, Y.; Lin, P.J.; Zhai, B.P.; Lu, Z.X.; Chapman, J.W.; Hu, G. Male nutritional status does not impact the reproductive potential of female Cnaphalocrocis medinalis moths under conditions of nutrient shortage. Insect Sci. 2022, 29, 467–477. [Google Scholar] [CrossRef]
- Hill, J.K.; Gatehouse, A.G. Genetic control of the pre-reproductive period in Autographa gamma (L.) (Silver Y moth) (Lepidoptera: Noctuidae). Heredity 1992, 69, 458–464. [Google Scholar] [CrossRef]
- Jiang, X.F.; Luo, L.Z.; Hu, Y. Genetic characteristics of pre-oviposition period in the oriental armyworm Mythimna separata (Walker). Acta Ecol. Sin. 2005, 25, 68–72. [Google Scholar]
- Zhang, L.; Pan, P.; Sappington, T.W.; Lu, W.X.; Luo, L.Z.; Jiang, X.F. Accelerated and synchronized oviposition induced by flight of young females may intensify larval outbreaks of the rice leaf roller. PLoS ONE 2015, 10, e0121821. [Google Scholar] [CrossRef] [PubMed]
- Li, K.B.; Gao, X.W.; Luo, L.Z.; Yin, J.; Cao, Y.Z. Changes in the activities of four related enzymes during the flight of oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 2005, 48, 643–647. [Google Scholar]
- Jindra, M.; Palli, S.R.; Riddiford, L.M. The juvenile hormone signaling pathway in insect development. Annu. Rev. Entomol. 2013, 58, 181–204. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Kim, Y. Purification and characterization of vitellin and vitellogenin of the beet armyworm, Spodoptera exigua (Noctuidae: Lepidoptera). J. Asia Pac. Entomol. 2003, 6, 37–43. [Google Scholar] [CrossRef]
- Saha, T.T.; Shin, S.W.; Dou, W.; Roy, S.; Zhao, B.; Hou, Y.; Wang, X.L.; Zou, Z.; Girke, T.; Raikhel, A.S. Hairy and Groucho mediate the action of juvenile hormone receptor methoprene-tolerant in gene repression. Proc. Natl. Acad. Sci. USA 2016, 113, 735–743. [Google Scholar] [CrossRef]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reproductive Parameters | Source | Df | F | p |
|---|---|---|---|---|
| Preoviposition period (days) | Population | 1 | 53.974 | <0.001 * |
| JHA treatment | 1 | 53.619 | <0.001 * | |
| Age | 3 | 4.084 | <0.001 * | |
| Population × JHA treatment | 1 | 0.019 | 0.890 | |
| Population × Age | 3 | 0.638 | 0.591 | |
| JHA treatment × Age | 3 | 6.625 | <0.001 * | |
| Population × JHA treatment × Age | 3 | 0.179 | 0.836 | |
| Sum of Squares Error | 460 | |||
| Lifetime fecundity (per female) | Population | 1 | 15.483 | <0.001 * |
| JHA treatment | 1 | 28.635 | <0.001 * | |
| Age | 3 | 3.545 | 0.015 * | |
| Population × JHA treatment | 1 | 0.145 | 0.703 | |
| Population × Age | 3 | 0.647 | 0.585 | |
| JHA treatment × Age | 3 | 2.628 | 0.050 * | |
| Population × JHA treatment × Age | 3 | 0.156 | 0.856 | |
| Sum of Squares Error | 460 | |||
| Oviposition period (days) | Population | 1 | 3.680 | 0.056 |
| JHA treatment | 1 | 0.856 | 0.355 | |
| Age | 3 | 1.296 | 0.237 | |
| Population × JHA treatment | 1 | 1.091 | 0.297 | |
| Population × Age | 3 | 0.370 | 0.774 | |
| JHA treatment × Age | 3 | 0.545 | 0.652 | |
| Population × JHA treatment × Age | 3 | 0.196 | 0.822 | |
| Sum of Squares Error | 460 | |||
| Mating frequency | Population | 1 | 1.616 | 0.204 |
| JHA treatment | 1 | 18.532 | <0.001 * | |
| Age | 3 | 1.487 | 0.150 | |
| Population × JHA treatment | 1 | 1.755 | 0.186 | |
| Population × Age | 3 | 2.179 | 0.090 | |
| JHA treatment × Age | 3 | 1.945 | 0.121 | |
| Population × JHA treatment × Age | 3 | 0.204 | 0.815 | |
| Sum of Squares Error | 460 | |||
| Mating percentage (%) | Population | 1 | 1.365 | 0.243 |
| JHA treatment | 1 | 14.986 | <0.001 * | |
| Age | 3 | 1.406 | 0.183 | |
| Population × JHA treatment | 1 | 0.026 | 0.871 | |
| Population × Age | 3 | 1.817 | 0.143 | |
| JHA treatment × Age | 3 | 0.726 | 0.537 | |
| Population × JHA treatment × Age | 3 | 0.177 | 0.838 | |
| Sum of Squares Error | 460 | |||
| Female longevity (days) | Population | 1 | 10.454 | 0.001 * |
| JHA treatment | 1 | 24.492 | <0.001 * | |
| Age | 3 | 2.637 | 0.006 * | |
| Population × JHA treatment | 1 | 2.573 | 0.109 | |
| Population × Age | 3 | 0.618 | 0.603 | |
| JHA treatment × Age | 3 | 8.663 | <0.001 * | |
| Population × JHA treatment × Age | 3 | 0.090 | 0.914 | |
| Sum of Squares Error | 460 | |||
| Male longevity (days) | Population | 1 | 1.060 | 0.304 |
| JHA treatment | 1 | 0.003 | 0.957 | |
| Age | 3 | 0.716 | 0.694 | |
| Population × JHA treatment | 1 | 0.264 | 0.607 | |
| Population × Age | 3 | 2.778 | 0.041 * | |
| JHA treatment × Age | 3 | 1.160 | 0.325 | |
| Population × JHA treatment × Age | 3 | 0.213 | 0.808 | |
| Sum of Squares Error | 460 | |||
| Ovarian development grade | Population | 1 | 1.814 | 0.064 |
| JHA treatment | 1 | 27.819 | <0.001 * | |
| Age | 3 | 0.919 | 0.508 | |
| Population × JHA treatment | 1 | 0.813 | 0.368 | |
| Population × Age | 3 | 0.250 | 0.861 | |
| JHA treatment × Age | 3 | 2.451 | 0.063 | |
| Population × JHA treatment × Age | 3 | 0.004 | 0.996 | |
| Sum of Squares Error | 460 |
| Parameters | Population | JHA Treatment | Age (Days after Emergence) | |||
|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | |
| Preoviposition period | 0.305 | <0.001 * | −0.318 | <0.001 * | 0.148 | 0.001 * |
| Lifetime fecundity | −0.185 | <0.001 * | 0.237 | <0.001 * | −0.127 | 0.006 * |
| Oviposition period | −0.097 | 0.035 * | −0.043 | 0.346 | 0.009 | 0.839 |
| Mating frequency | −0.060 | 0.196 | 0.203 | <0.001 * | −0.059 | 0.198 |
| Mating percentage | −0.057 | 0.219 | 0.186 | <0.001 * | −0.022 | 0.629 |
| Female longevity | 0.120 | 0.009 * | −0.245 | <0.001 * | 0.129 | 0.005 * |
| Male longevity | 0.048 | 0.297 | 0.000 | 0.992 | 0.055 | 0.231 |
| Ovarian development grade | −0.107 | 0.020 * | 0.239 | <0.001 * | −0.066 | 0.154 |
| Parameters | Population | JHA Treatment | Age (Days after Emergence) | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||
| Preoviposition period (days) | Residents | JHA | 3.97 ± 0.13 b | 4.90 ± 0.19 a | 5.08 ± 0.21 a | 5.24 ± 0.25 a |
| Control | 5.93 ± 0.28 a | 5.67 ± 0.25 a | 5.73 ± 0.30 a | 5.87 ± 0.23 a | ||
| Migrants | JHA | 4.97 ± 0.18 b | 6.17 ± 0.21 a | 5.89 ± 0.31 a | 6.07 ± 0.26 a | |
| Control | 7.07 ± 0.32 a | 6.68 ± 0.33 a | 6.86 ± 0.40 a | 6.87 ± 0.34 a | ||
| Lifetime fecundity (per female) | Residents | JHA | 1125.52 ± 62.56 a | 971.90 ± 50.04 ab | 879.62 ± 63.43 b | 867.93 ± 74.02 b |
| Control | 778.43 ± 59.67 a | 791.27 ± 66.55 a | 769.40 ± 63.69 a | 751.43 ± 48.26 a | ||
| Migrants | JHA | 962.40 ± 47.12 a | 756.17 ± 66.55 ab | 749.45 ± 65.45 b | 754.03 ± 64.56 ab | |
| Control | 670.40 ± 64.29 a | 636.71 ± 78.41 a | 653.03 ± 75.56 a | 636.07 ± 65.49 a | ||
| Oviposition period (days) | Residents | JHA | 5.50 ± 0.28 a | 5.58 ± 0.32 a | 6.08 ± 0.35 a | 5.48 ± 0.26 a |
| Control | 6.17 ± 0.35 a | 5.70 ± 0.37 a | 6.27 ± 0.31 a | 6.21 ± 0.31 a | ||
| Migrants | JHA | 5.23 ± 0.23 a | 5.60 ± 0.43 a | 5.66 ± 0.44 a | 5.53 ± 0.33 a | |
| Control | 5.53 ± 0.43 a | 5.46 ± 0.50 a | 5.03 ± 0.40 a | 5.57 ± 0.46 a | ||
| Mating frequency | Residents | JHA | 1.37 ± 0.11 a | 1.16 ± 0.13 a | 0.96 ± 0.12 a | 0.93 ±0.14 a |
| Control | 0.73 ± 0.11 a | 0.80 ± 0.11 a | 0.77 ± 0.13 a | 0.67 ± 0.11 a | ||
| Migrants | JHA | 1.07 ± 0.12 a | 0.87 ± 0.12 a | 0.93 ± 0.12 a | 0.90 ± 0.11 a | |
| Control | 0.70 ± 0.12 a | 0.61 ± 0.12 a | 0.72 ± 0.13 a | 0.83 ± 0.13 a | ||
| Mating percentage (%) | Residents | JHA | 96.67 ± 3.33 a | 83.87 ± 6.72 a | 80.77 ± 7.88 a | 72.41 ± 8.45 a |
| Control | 66.67 ± 8.75 a | 70.00 ± 8.23 a | 63.33± 8.95 a | 60.00 ± 4.44 a | ||
| Migrants | JHA | 83.33 ± 6.92 a | 70.00 ± 8.51 a | 79.31 ± 7.66 a | 76.67 ± 7.85 a | |
| Control | 60.00 ± 9.10 a | 53.57 ± 9.60 a | 58.62 ± 9.31 a | 66.67 ± 8.75 a | ||
| Female longevity (days) | Residents | JHA | 9.10 ± 0.32 b | 10.48 ± 0.35 a | 11.63 ± 0.33 a | 10.72 ± 0.32 a |
| Control | 12.10 ± 0.41 a | 11.23 ± 0.35 a | 11.69 ± 0.36 a | 11.36 ± 0.38 a | ||
| Migrants | JHA | 10.23 ± 0.27 b | 11.80 ± 0.46 a | 11.55 ± 0.45 ab | 11.43 ± 0.41 ab | |
| Control | 12.70 ± 0.42 a | 11.89 ± 0.50 a | 11.79 ± 0.44 a | 12.37 ± 0.51 a | ||
| Male longevity (days) | Residents | JHA | 14.73 ± 0.90 a | 13.48 ± 0.70 a | 13.80 ± 0.94 a | 13.28 ± 0.82 a |
| Control | 13.67 ± 0.64 a | 14.43 ± 0.67 a | 13.10 ± 0.54 a | 13.37 ± 0.49 a | ||
| Migrants | JHA | 13.57 ± 0.39 a | 13.00 ± 0.72 a | 14.38 ± 0.88 a | 15.00 ± 0.41 a | |
| Control | 13.23 ± 0.61 a | 14.04 ± 0.75 a | 14.55 ± 0.76 a | 15.17 ± 0.94 a | ||
| Ovarian development grade | Residents | JHA | 4.77 ± 0.08 a | 4.65 ± 0.09 a | 4.54 ± 0.10 a | 4.55 ± 0.09 a |
| Control | 4.30 ± 0.09 a | 4.37 ± 0.09 a | 4.37 ± 0.09 a | 4.33 ± 0.09 a | ||
| Migrants | JHA | 4.63 ± 0.09 a | 4.43 ± 0.09 a | 4.41 ± 0.09 a | 4.40 ± 0.09 a | |
| Control | 4.23 ± 0.08 a | 4.25 ± 0.08 a | 4.34 ± 0.09 a | 4.30 ± 0.09 a | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, W.; Zeng, L.; Zhang, Z.; He, H.; Wang, F.; Xie, X. Effects of Juvenile Hormone Analog and Days after Emergence on the Reproduction of Oriental Armyworm, Mythimna separata (Lepidoptera: Noctuidae) Populations. Insects 2022, 13, 506. https://doi.org/10.3390/insects13060506
Lv W, Zeng L, Zhang Z, He H, Wang F, Xie X. Effects of Juvenile Hormone Analog and Days after Emergence on the Reproduction of Oriental Armyworm, Mythimna separata (Lepidoptera: Noctuidae) Populations. Insects. 2022; 13(6):506. https://doi.org/10.3390/insects13060506
Chicago/Turabian StyleLv, Weixiang, Liting Zeng, Zhe Zhang, Hengguo He, Fang Wang, and Xingcheng Xie. 2022. "Effects of Juvenile Hormone Analog and Days after Emergence on the Reproduction of Oriental Armyworm, Mythimna separata (Lepidoptera: Noctuidae) Populations" Insects 13, no. 6: 506. https://doi.org/10.3390/insects13060506
APA StyleLv, W., Zeng, L., Zhang, Z., He, H., Wang, F., & Xie, X. (2022). Effects of Juvenile Hormone Analog and Days after Emergence on the Reproduction of Oriental Armyworm, Mythimna separata (Lepidoptera: Noctuidae) Populations. Insects, 13(6), 506. https://doi.org/10.3390/insects13060506