
Mitochondrial Genomes Provide New Phylogenetic and Evolutionary Insights into Psilidae (Diptera: Brachycera)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and DNA Extraction
2.2. Mitochondrial Genome Sequencing and Assembly
2.3. Mitochondrial Genome Annotation and Analysis
2.4. Phylogenetic Analysis
3. Results and Discussion
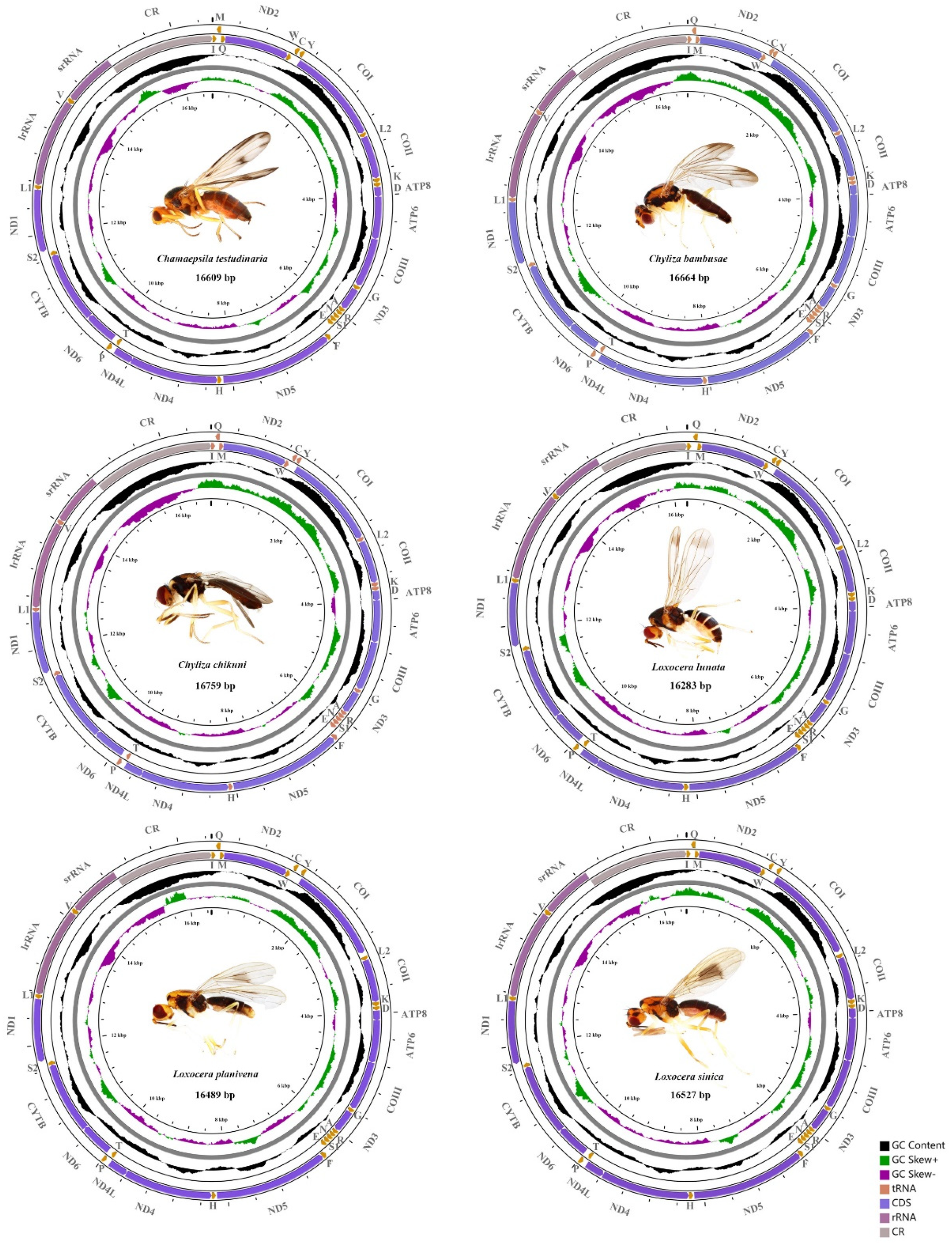
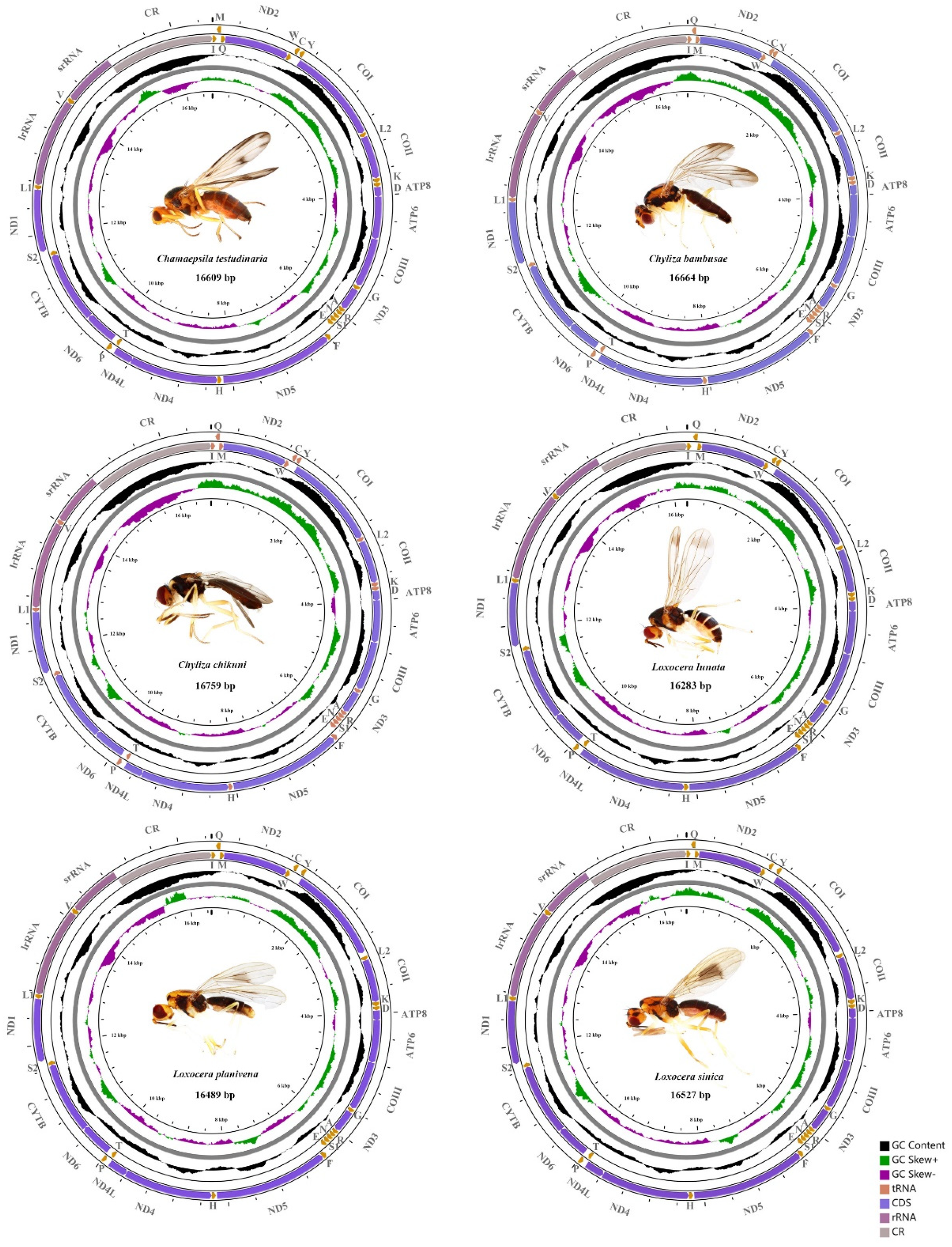
3.1. General Structure and Nucleotide Composition of Psilidae Mitogenomes
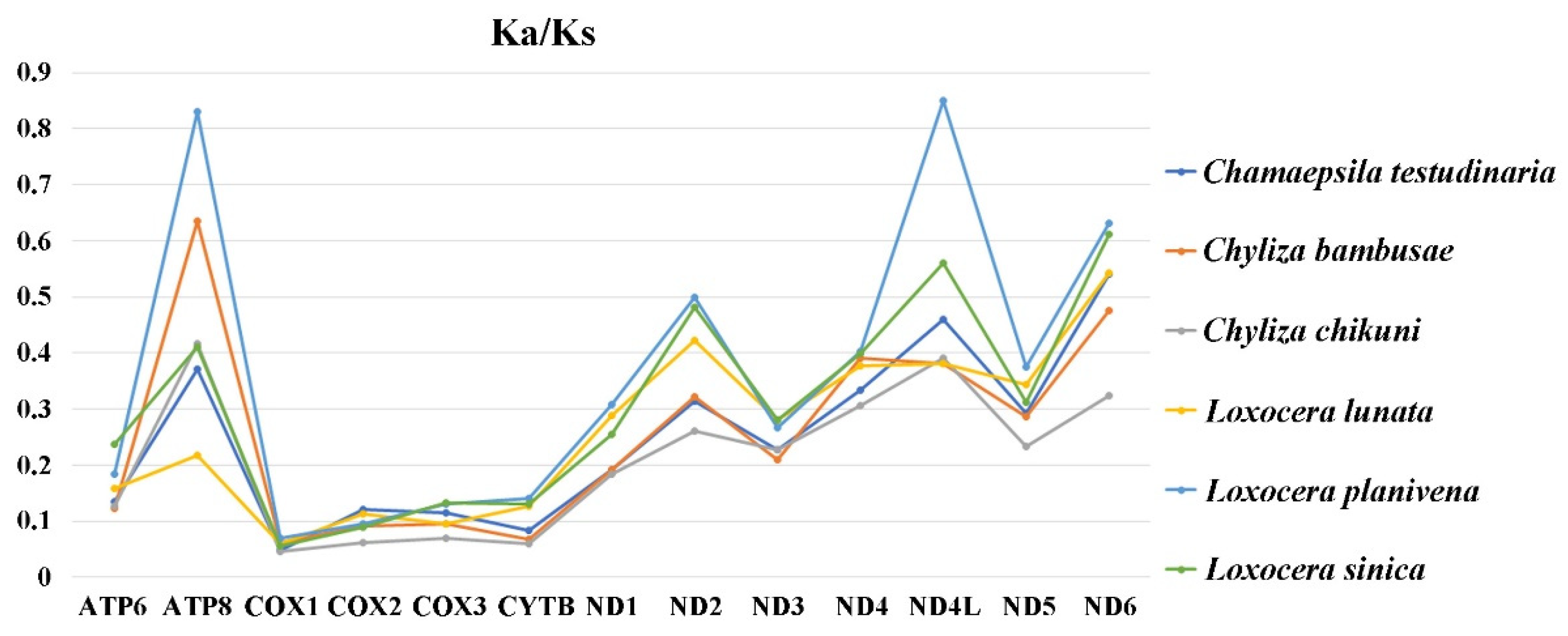
3.2. Protein-Coding Genes, Codon Usage, and Evolutionary Rates
3.3. Transfer and Ribosomal RNA Genes
3.4. Control Region
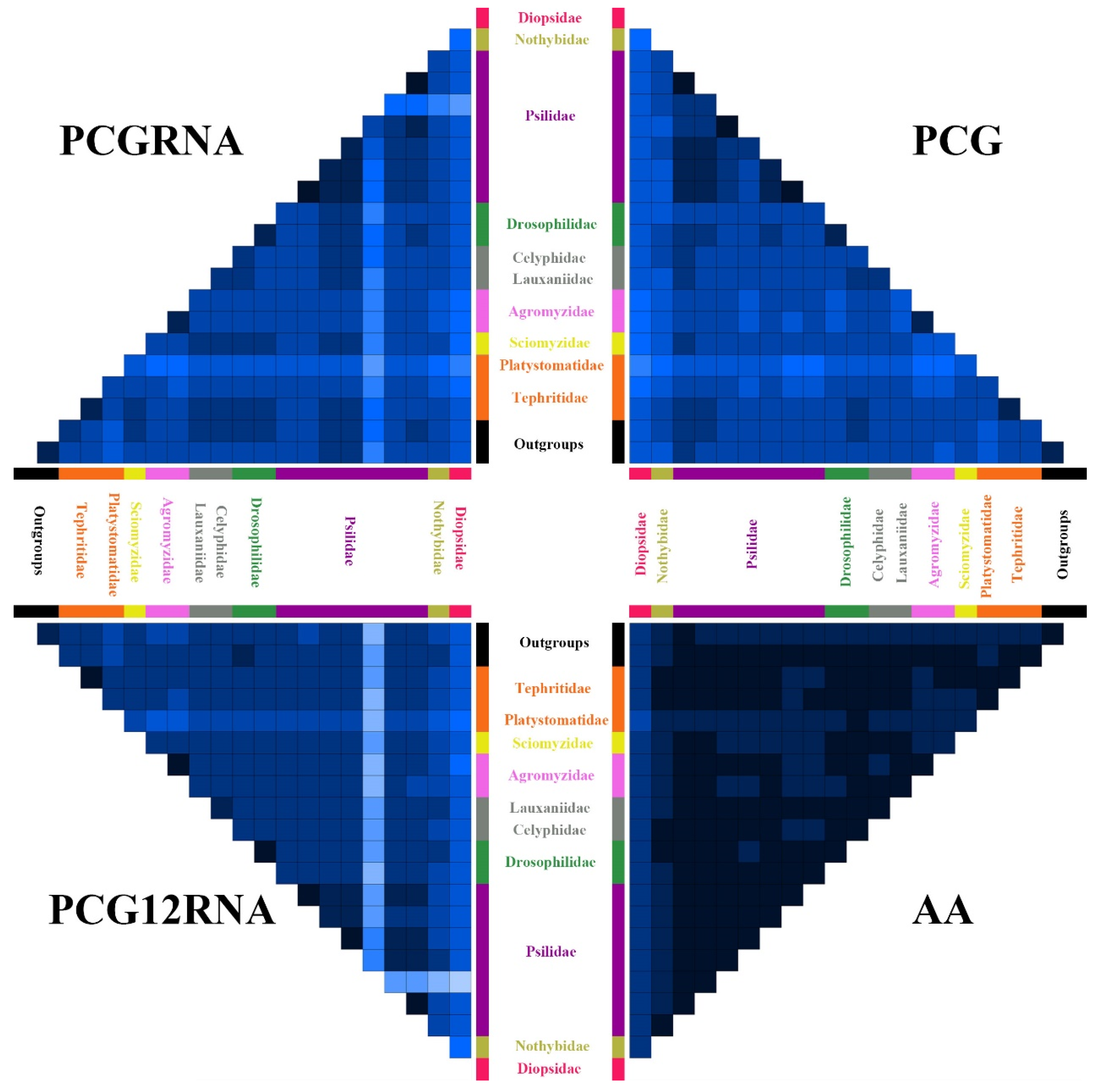
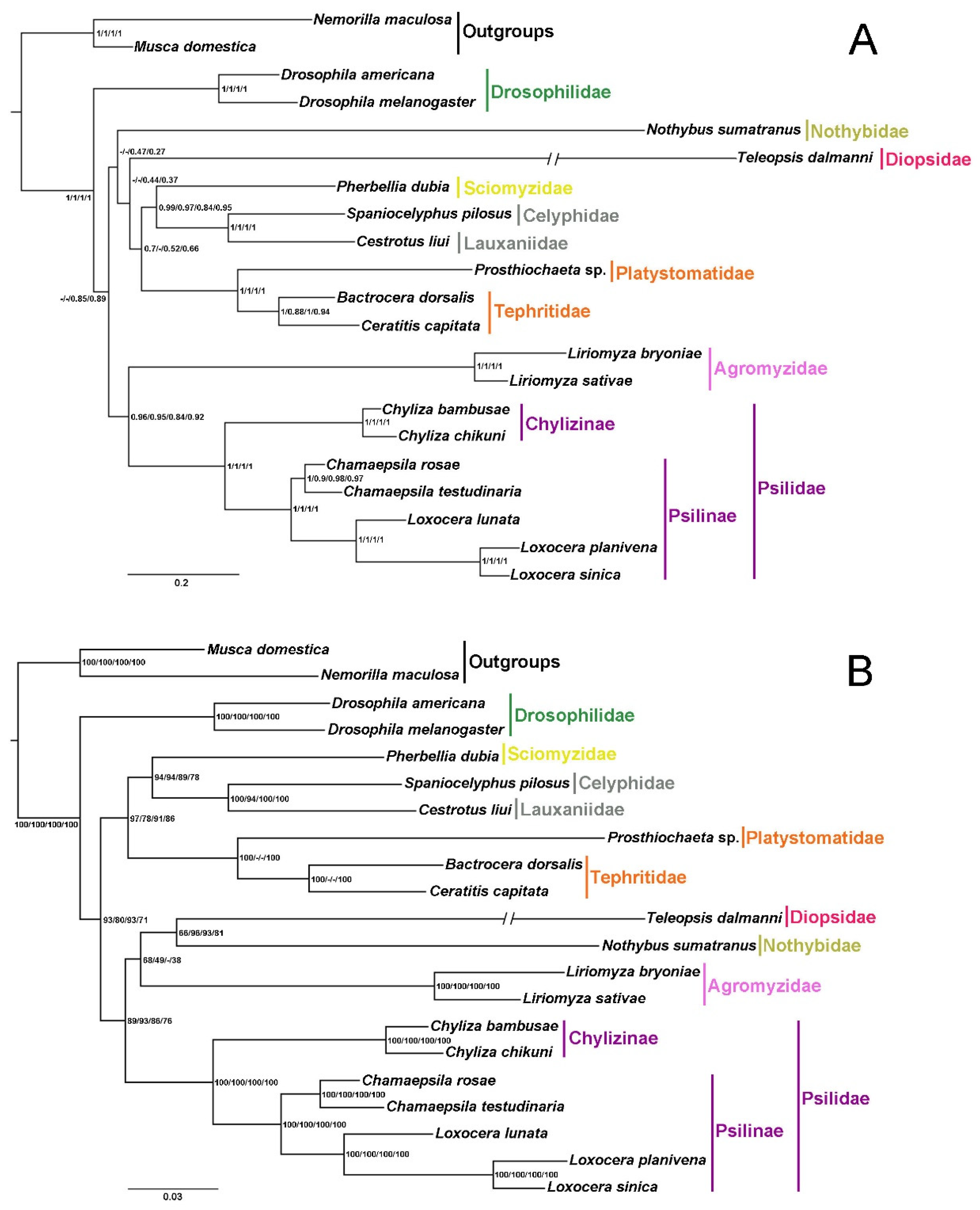
3.5. Phylogenetic Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More than just a powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taanman, J.-W. The mitochondrial genome: Structure, transcription, translation and replication. Biochim. Biophys. Acta Bioenerg. 1999, 1410, 103–123. [Google Scholar] [CrossRef] [Green Version]
- Saccone, C.; Gissi, C.; Lanave, C.; Larizza, A.; Pesole, G.; Reyes, A. Evolution of the mitochondrial genetic system: An overview. Gene 2000, 261, 153–159. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.A.; Lambkin, C.L.; Batterham, P.; Wallman, J.F.; Dowton, M.; Whiting, M.F.; Yeates, D.K.; Cameron, S.L. Beyond barcoding: A mitochondrial genomics approach to molecular phylogenetics and diagnostics of blowflies (Diptera: Calliphoridae). Gene 2012, 511, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Moritz, C.; Dowling, T.E.; Brown, W.M. Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Ann. Rev. Ecol. Syst. 1987, 18, 269–292. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Jiang, P.; Zhou, X.; Liu, J.; Sun, C.; Vogler, A.P.; Cai, W. Capturing the phylogeny of Holometabola with mitochondrial genome data and Bayesian site-heterogeneous mixture models. Genome Biol. Evol. 2016, 8, 1411–1426. [Google Scholar] [CrossRef]
- Li, H.; Leavengood, J.M.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.; Cai, W. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. R. Soc. B 2017, 284, 20171223. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Liu, G.; Wang, W.; James, P.; Colwell, D.D.; Tran, A.; Gong, S.; Cai, W.; Shao, R. Mitochondrial genome fragmentation unites the parasitic lice of eutherian mammals. Syst. Biol. 2019, 68, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Zink, R.M.; Barrowclough, G.F. Mitochondrial DNA under siege in avian phylogeography. Mol. Ecol. 2008, 17, 2107–2121. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yang, P.; Jiang, F.; Chapuis, M.-P.; Shali, Y.; Sword, G.A.; Kang, L. Mitochondrial genomes reveal the global phylogeography and dispersal routes of the migratory locust. Mol. Ecol. 2012, 21, 4344–4358. [Google Scholar] [CrossRef] [PubMed]
- Salvato, P.; Simonato, M.; Battisti, A.; Negrisolo, E. The complete mitochondrial genome of the bag-shelter moth Ochrogaster lunifer (Lepidoptera: Notodontidae). BMC Genom. 2008, 9, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, R.; Zhu, X.Q.; Barker, S.C.; Herd, K. Evolution of extensively fragmented mitochondrial genomes in the lice of humans. Genome Biol. Evol. 2012, 4, 1088–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curole, J.P.; Kocher, T.D. Mitogenomics: Digging deeper with complete mitochondrial genomes. Trends Ecol. Evol. 1999, 14, 394–398. [Google Scholar] [CrossRef]
- Timmermans, M.J.T.N.; Dodsworth, S.; Culverwell, C.L.; Bocak, L.; Ahrens, D.; Littlewood, D.T.J.; Pons, J.; Vogler, A.P. Why barcode? High-throughput multiplex sequencing of mitochondrial genomes for molecular systematics. Nucleic Acids Res. 2010, 38, e197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Samuels, D.C.; Clark, T.; Guo, Y. High-throughput sequencing in mitochondrial DNA research. Mitochondrion 2014, 17, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L. Insect mitochondrial genomics implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L.; Lambkin, C.L.; Barker, S.C.; Whiting, M.F. A mitochondrial genome phylogeny of Diptera: Whole genome sequence data accurately resolve relationships over broad timescales with high precision. Syst. Entomol. 2007, 32, 40–59. [Google Scholar] [CrossRef]
- Yan, L.; Pape, T.; Elgar, M.A.; Gao, Y.; Zhang, D. Evolutionary history of stomach bot flies in the light of mitogenomics. Syst. Entomol. 2019, 44, 797–809. [Google Scholar] [CrossRef]
- Li, X.; Yan, L.; Pape, T.; Gao, Y.; Zhang, D. Evolutionary insights into bot flies (Insecta: Diptera: Oestridae) from comparative analysis of the mitochondrial genomes. Int. J. Biol. Macromol. 2020, 149, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-L.; Zhao, Y.-M.; Yan, L.-P.; Liu, W.-B.; Bu, W.-J.; Wang, X.-H.; Zheng, C.-G. Mitogenomes provide new insights into the evolutionary history of Prodiamesinae (Diptera: Chironomidae). Zool. Scr. 2022, 51, 119–132. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, D.; Kang, Z. New data on the mitochondrial genome of Nematocera (lower Diptera): Features, structures and phylogenetic implications. Zool. J. Linn. Soc. 2022, zlac012. [Google Scholar] [CrossRef]
- Marshall, S.A. Flies: The Natural History and Diversity of Diptera; Firefly Books Ltd.: Richmond Hill, ON, Canada, 2012; p. 616. [Google Scholar]
- Bayless, K.M.; Trautwein, M.D.; Meusemann, K.; Shin, S.; Petersen, M.; Donath, A.; Podsiadlowski, L.; Mayer, C.; Niehuis, O.; Peters, R.S.; et al. Beyond Drosophila: Resolving the rapid radiation of schizophoran flies with phylotranscriptomics. BMC Biol. 2021, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Shatalkin, A.I. Psilidae. Manual of Afrotropical Diptera. In Brachycera–Cyclorrhapha, Excluding Calyptratae; Krik-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2021; Volume 3, pp. 1545–1556. [Google Scholar]
- Shatalkin, A.I.; Merz, B. The Psilidae (Diptera) from Switzerland, with description of new Central European species. Rev. Suisse Zool. 2010, 117, 771–800. [Google Scholar] [CrossRef]
- Lonsdale, O. Family groups of Diopsoidea and Nerioidea (Diptera: Schizophora)–Definition, history and relationships. Zootaxa 2020, 4735, 1–177. [Google Scholar] [CrossRef]
- Yang, C.; Wang, X.L. A new species of Chyliza (Psilidae: Diptera) from China, injuring bamboo roots. For. Res. 1988, 1, 275–277. [Google Scholar]
- Xu, T.; Lv, R. A study on Chyliza bambusae. For. Res. 1988, 1, 278–284. [Google Scholar]
- Iwasa, M.; Hanada, T.; Kajino, Y. A new psilid species from Japan injurious to the root of carrot (Diptera: Psilidae). Appl. Entomol. Zool. 1987, 22, 310–315. [Google Scholar] [CrossRef]
- Collier, R.; Finch, S. A review of research to address carrot fly (Psila rosae) control in the UK. EPPO Bull. 2009, 39, 121–127. [Google Scholar] [CrossRef]
- Glendenning, R. Psila nigricornis Meig. (Diptera, Psilidae), a new pest of possible economic importance. Can. Entomol. 1952, 84, 107. [Google Scholar] [CrossRef]
- Ellis, P.R.; Hardman, J.A.; Saw, P.L. Host plants of the carrot fly, Psila rosae (F.) (Dipt., Psilidae). Entomol. Mon. Mag. 1992, 128, 1–10. [Google Scholar]
- Chandler, P.J. Observations on plant associations of the Psilidae (Diptera). Entomol. Rec. J. Var. 1975, 87, 13–17. [Google Scholar]
- Ferrar, P. A Guide to Breeding Habits and Immature Stages of Diptera Cyclorrhapha. In Entomonograph; Brill: Leiden, The Netherlands, 1987; Volume 8, pp. 1–478. [Google Scholar]
- Buck, M. Psilidae (Rust Flies, Psilid Flies). In Manual of Central American Diptera; Brown, B.V., Borkent, A., Cumming, J.M., Woodley, N.E., Zumbado, M.A., Eds.; NRC Research Press: Ottawa, ON, Canada, 2010; Volume 2, pp. 837–842. [Google Scholar]
- Buck, M.; Marshall, S.A. The identity of Pseudopsila, description of a new subgenus of Psila, and redefinition of Psila sensu lato (Diptera: Psilidae). Eur. J. Entomol. 2006, 103, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Buck, M.; Marshall, S.A. Revision of New World Loxocera (Diptera: Psilidae), with phylogenetic redefinition of Holarctic subgenera and species groups. Eur. J. Entomol. 2006, 103, 193–219. [Google Scholar] [CrossRef] [Green Version]
- Shatalkin, A.I. Asian species of Loxocera Meigen (Diptera, Psilidae). Russ. Entomol. J. 1998, 6, 87–97. [Google Scholar]
- Shatalkin, A.I. Afrotropical Psilidae (Diptera). 1. Genera Belobackenbardia gen. n. and Psila Meigen, 1803. Russ. Entomol. J. 2002, 10, 417–424. [Google Scholar]
- Wang, X. Two new species of the genus Chyliza (Diptera: Psilidae) from China. Entomotaxonomia 1995, 17, 100–102. [Google Scholar]
- Wang, X.; Yang, C. Psilidae. In Flies of China; Xue, W., Chao, C., Eds.; Liaoning Science and Technology Press: Shenyang, China, 1998; Volume 1, pp. 424–456. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE on-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Laslett, D.; Canbäck, B. ARWEN, a program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486–487. [Google Scholar] [CrossRef]
- Yang, Z.; Bielawski, J.R. Statistical methods for detecting molecular adaptation. Trends Ecol. Evol. 2000, 15, 496–503. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Su, S.; Yang, D. The complete mitochondrial genomes of Musca domestica and Scathophaga stercoraria (Diptera: Muscoidea: Muscidae and Scathophagidae). Mitochondrial DNA A 2016, 27, 1435–1436. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, M.; Yang, D. The mitochondrial genome of Nothybus sumatranus (Diptera: Nothybidae). Mitochondrial DNA B 2021, 6, 1573–1574. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Ding, S.; Cameron, S.L.; Mao, M.; Shi, L.; Yang, D. Mitochondrial genomes provide insights into the phylogeny of Lauxanioidea (Diptera: Cyclorrhapha). Int. J. Mol. Sci. 2017, 18, 773. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Du, Y.; Cao, J.; Huang, F. Analysis of three leafminers’ complete mitochondrial genomes. Gene 2013, 529, 1–6. [Google Scholar] [CrossRef]
- Wang, S.; Lei, Z.; Wen, J.; Wang, H.; Li, X.; Dong, B.; Ren, B. The complete mitochondrial genome of Liriomyza huidobrensis and comparison with L. trifolii and L. sativae (Diptera: Agromyzidae). Mitochondrial DNA A 2013, 25, 104–105. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, J.; Wang, L.; Hou, P.; Yang, D. The mitochondrial genome of Prosthiochaeta sp. (Diptera: Platystomatidae). Mitochondrial DNA B 2020, 5, 2557–2558. [Google Scholar] [CrossRef]
- Spanos, L.; Koutroumbas, G.; Kotsyfakis, M.; Louis, C. The mitochondrial genome of the mediterranean fruit fly, Ceratitis capitata. Insect Mol. Biol. 2000, 9, 139–144. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kück, P.; Meid, S.A.; Groß, C.; Wägele, J.W.; Misof, B. AliGROOVE–visualization of heterogeneous sequence divergence within multiple sequence alignments and detection of inflated branch support. BMC Bioinform. 2014, 15, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lartillot, N.; Rodrigue, N.; Stubbs, D.; Richer, J. PhyloBayes MPI: Phylogenetic reconstruction with infinite mixtures of profiles in a parallel environment. Syst. Biol. 2013, 62, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast-online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, Y.; Wu, Y.; Song, F.; Cai, W.; Li, H. Novel tRNA gene rearrangements in the mitochondrial genome of Camarochiloides weiweii (Hemiptera: Pachynomidae). Int. J. Biol. Macromol. 2020, 165, 1738–1744. [Google Scholar] [CrossRef]
- Timmermans, M.J.; Vogler, A.P. Phylogenetically informative rearrangements in mitochondrial genomes of Coleoptera, and monophyly of aquatic elateriform beetles (Dryopoidea). Mol. Phylogenet. Evol. 2012, 63, 299–304. [Google Scholar] [CrossRef]
- Tang, P.; Zhu, J.; Zheng, B.; Wei, S.; Sharkey, M.; Chen, X.; Vogler, A.P. Mitochondrial phylogenomics of the Hymenoptera. Mol. Phylogenet. Evol. 2019, 131, 8–18. [Google Scholar] [CrossRef]
- Behura, S.K.; Lobo, N.F.; Haas, B.; deBruyn, B.; Lovin, D.D.; Shumway, M.F.; Puiu, D.; Romero-Severson, J.; Nene, V.; Severson, D.W. Complete sequences of mitochondria genomes of Aedes aegypti and Culex quinquefasciatus and comparative analysis of mitochondrial DNA fragments inserted in the nuclear genomes. Insect Biochem. Mol. Biol. 2011, 41, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Beckenbach, A.T.; Joy, J.B. Evolution of the mitochondrial genomes of gall midges (Diptera: Cecidomyiidae): Rearrangement and severe truncation of tRNA genes. Genome Biol. Evol. 2009, 1, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.J.; Shi, M.; Chen, X.X.; Sharkey, M.J.; van Achterberg, C.; Ye, G.Y.; He, J.H. New views on strand asymmetry in insect mitochondrial genomes. PLoS ONE 2010, 5, e12708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, E.P.C.; Danchin, A. Base composition bias might result from competition for metabolic resources. Trends Genet. 2002, 18, 291–294. [Google Scholar] [CrossRef]
- Foerstner, K.U.; von Mering, C.; Hooper, S.D.; Bork, P. Environments shape the nucleotide composition of genomes. EMBO Rep. 2005, 6, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-H.; Lu, G.; Bork, P.; Hu, S.; Lercher, M.J. Energy efficiency trade-offs drive nucleotide usage in transcribed regions. Nat. Commun. 2016, 7, 11334. [Google Scholar] [CrossRef] [Green Version]
- Castro, L.R.; Austin, A.D.; Dowton, M. Contrasting rates of mitochondrial molecular evolution in parasitic Diptera and Hymenoptera. Mol. Biol. Evol. 2002, 19, 1100–1113. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Su, T.; Chesters, D.; Wang, S.; Ho, S.Y.W.; Zhu, C.; Chen, X.; Zhang, C. The mitochondrial genome of Elodia flavipalpis Aldrich (Diptera: Tachinidae) and the evolutionary timescale of tachinid flies. PLoS ONE 2013, 8, e61814. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Li, J.; Li, H. The complete mitochondrial genome of Syritta pipiens (Linnaeus, 1758) (Diptera: Syrphidae) and phylogenetic analysis. Mitochondrial DNA B 2021, 6, 2475–2477. [Google Scholar] [CrossRef]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef]
- Zheng, N.; Sun, Y.X.; Yang, L.L.; Wu, L.; Abbas, M.N.; Chen, C.; Gao, J.; Li, X.K.; Liu, C.L.; Dai, L.S. Characterization of the complete mitochondrial genome of Biston marginata (Lepidoptera: Geometridae) and phylogenetic analysis among lepidopteran insects. Int. J. Biol. Macromol. 2018, 113, 961–970. [Google Scholar] [CrossRef]
- Dashnow, H.; Tan, S.; Das, D.; Easteal, S.; Oshlack, A. Genotyping microsatellites in next-generation sequencing data. BMC Bioinform. 2015, 16, A5. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Shao, R.; Song, N.; Song, F.; Jiang, P.; Li, Z.; Cai, W. Higher-level phylogeny of paraneopteran insects inferred from mitochondrial genome sequences. Sci. Rep. 2015, 5, 8527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, M.J.T.N.; Barton, C.; Haran, J.; Ahrens, D.; Culverwell, C.L.; Ollikainen, A.; Dodsworth, S.; Foster, P.G.; Bocak, L.; Vogler, A.P. Family-level sampling of mitochondrial genomes in Coleoptera: Compositional heterogeneity and phylogenetics. Genome Biol. Evol. 2015, 8, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Song, F.; Jiang, P.; Wilson, J.-J.; Cai, W.; Li, H. Compositional heterogeneity in true bug mitochondrial phylogenomics. Mol. Phylogenet. Evol. 2018, 118, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Shatalkin, A.I. Belobackenbardia mostovskii sp.n. (Diptera, Psilidae) is the fourth species of an endemic genus found in South Africa. Russian Entomol. J. 2021, 30, 342–344. [Google Scholar] [CrossRef]
- Hennig, W. Neue untersuchungen über die familien der Diptera Schizophora (Diptera: Cyclorrhapha). Stuttg. Beitr. Naturkd. 1971, 226, 5–76. [Google Scholar]
- McAlpine, J.F. Phylogeny and Classification of the Muscomorpha. In Manual of Nearctic Diptera; McAlpine, J.F., Wood, D.M., Eds.; Research Branch Agriculture Canada: Ottawa, ON, Canada, 1989; Volume 3, pp. 1397–1518. [Google Scholar]
- Lambkin, C.; Pape, T.; Sinclair, B.J.; Courtney, G.W.; Skevington, J.H.; Meier, R.; Yeates, D.K.; Blagoderov, V.; Wiegmann, B.M. The phylogenetic relationships among infraorders and superfamilies of Diptera based on morphological evidence. Syst. Entomol. 2013, 38, 164–179. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic radiations in the fly tree of life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, A.R.; Sarkar, I.N. The impact of taxon sampling on phylogenetic inference: A review of two decades of controversy. Brief. Bioinform. 2012, 13, 122–134. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superfamily | Family | Species | GenBank Number | Reference |
|---|---|---|---|---|
| Outgroup | ||||
| Muscoidea | Muscidae | Musca domestica | NC_024855 | [59] |
| Oestroidea | Tachinidae | Nemorilla maculosa | MG786426 | Direct submission |
| Ingroup | ||||
| Diopsoidea | Diopsidae | Teleopsis dalmanni | CM026973 | Direct submission |
| Nothybidae | Nothybus sumatranus | MW387954 | [60] | |
| Psilidae | Chamaepsila rosae | MT941918 | Direct submission | |
| Chamaepsila testudinaria | ON258616 | Present study | ||
| Chyliza bambusae | ON258617 | Present study | ||
| Chyliza chikuni | ON258618 | Present study | ||
| Loxocera lunata | ON258619 | Present study | ||
| Loxocera planivena | ON258620 | Present study | ||
| Loxocera sinica | ON258621 | Present study | ||
| Ephydroidea | Drosophilidae | Drosophila americana | MK659804 | Direct submission |
| Drosophila melanogaster | NC_024511 | Direct submission | ||
| Lauxanioidea | Celyphidae | Spaniocelyphus pilosus | KX372562 | [61] |
| Lauxaniidae | Cestrotus liui | KX372559 | [61] | |
| Opomyzoidea | Agromyzidae | Liriomyza bryoniae | JN570504 | [62] |
| Liriomyza sativae | JQ862475 | [63] | ||
| Sciomyzoidea | Sciomyzidae | Pherbellia dubia | MT628567 | Direct submission |
| Tephritoidea | Platystomatidae | Prosthiochaeta sp. | MT528242 | [64] |
| Tephritidae | Bactrocera dorsalis | KT343905 | Direct submission | |
| Ceratitis capitata | NC_000857 | [65] |
| Species | Regions | Length (bp) | T% | C% | A% | G% | A + T% | AT Skew | GC Skew |
|---|---|---|---|---|---|---|---|---|---|
| Chamaepsila testudinaria | Whole genome | 16,609 | 37.3 | 12.2 | 41.5 | 9.1 | 78.8 | 0.053 | −0.147 |
| PCGs | 11,184 | 44.5 | 11.4 | 32.2 | 11.9 | 76.7 | −0.16 | 0.023 | |
| 1st codon position | 5537 | 37 | 12.6 | 41.6 | 8.8 | 78.6 | 0.058 | −0.181 | |
| 2nd codon position | 5536 | 39.9 | 10.5 | 42.2 | 7.4 | 82.1 | 0.028 | −0.171 | |
| 3rd codon position | 5536 | 35 | 13.4 | 40.6 | 11 | 75.6 | 0.075 | −0.1 | |
| tRNAs | 1469 | 38.7 | 9.9 | 38.5 | 12.9 | 77.3 | −0.003 | 0.132 | |
| rRNAs | 2131 | 43.7 | 6.3 | 38.2 | 11.7 | 81.9 | −0.066 | 0.299 | |
| Control region | 1658 | 37.1 | 6.3 | 51.7 | 4.8 | 88.8 | 0.165 | −0.135 | |
| Chyliza bambusae | Whole genome | 16,664 | 36.7 | 13.1 | 41.6 | 8.6 | 78.3 | 0.062 | −0.207 |
| PCGs | 11,182 | 44.4 | 11.6 | 31.4 | 12.6 | 75.8 | −0.172 | 0.041 | |
| 1st codon position | 5555 | 38.5 | 12.5 | 42.4 | 6.5 | 81 | 0.048 | −0.317 | |
| 2nd codon position | 5555 | 35.2 | 13.1 | 41.5 | 10.2 | 76.7 | 0.081 | −0.124 | |
| 3rd codon position | 5554 | 36.3 | 13.7 | 40.8 | 9.1 | 77.2 | 0.058 | −0.2 | |
| tRNAs | 1457 | 39 | 9.8 | 38.6 | 12.7 | 77.6 | −0.005 | 0.132 | |
| rRNAs | 2116 | 43.1 | 6.1 | 38.4 | 12.5 | 81.5 | −0.058 | 0.347 | |
| Control region | 1858 | 37.1 | 7.3 | 52.4 | 3.2 | 89.6 | 0.171 | −0.392 | |
| Chyliza chikuni | Whole genome | 16,759 | 36.5 | 14.1 | 40.5 | 9 | 77 | 0.052 | −0.222 |
| PCGs | 11,182 | 43.4 | 12.7 | 30.8 | 13.2 | 74.1 | −0.17 | 0.019 | |
| 1st codon position | 5587 | 36 | 13.7 | 40.8 | 9.5 | 76.9 | 0.062 | −0.182 | |
| 2nd codon position | 5586 | 36.8 | 14.4 | 39.5 | 9.3 | 76.3 | 0.035 | −0.214 | |
| 3rd codon position | 5586 | 36.6 | 14.2 | 41.2 | 8.1 | 77.8 | 0.059 | −0.274 | |
| tRNAs | 1456 | 39 | 9.8 | 38.5 | 12.6 | 77.5 | −0.006 | 0.125 | |
| rRNAs | 2114 | 42.8 | 6.4 | 38.3 | 12.6 | 81 | −0.055 | 0.327 | |
| Control region | 1941 | 38.6 | 8.5 | 49.4 | 3.5 | 88 | 0.122 | −0.416 | |
| Loxocera lunata | Whole genome | 16,283 | 38.9 | 11.9 | 40.7 | 8.5 | 79.6 | 0.023 | −0.165 |
| PCGs | 11,196 | 43.8 | 10.9 | 33.6 | 11.7 | 77.4 | −0.132 | 0.038 | |
| 1st codon position | 5428 | 40.3 | 12.4 | 38.8 | 8.5 | 79.1 | −0.02 | −0.184 | |
| 2nd codon position | 5428 | 39.2 | 11.3 | 41.2 | 8.3 | 80.4 | 0.025 | −0.151 | |
| 3rd codon position | 5427 | 37 | 12 | 42.2 | 8.8 | 79.2 | 0.065 | −0.157 | |
| tRNAs | 1458 | 40.2 | 8.9 | 38.6 | 12.3 | 78.8 | −0.02 | 0.165 | |
| rRNAs | 2118 | 43.5 | 5.9 | 39.3 | 11.3 | 82.8 | −0.051 | 0.315 | |
| Control region | 1389 | 45.3 | 5.3 | 46.6 | 2.9 | 91.9 | 0.014 | −0.292 | |
| Loxocera planivena | Whole genome | 16,489 | 38.8 | 11.8 | 41.1 | 8.2 | 80 | 0.029 | −0.177 |
| PCGs | 11,183 | 44.3 | 11 | 33.5 | 11.2 | 77.7 | −0.139 | 0.009 | |
| 1st codon position | 5497 | 35 | 11.2 | 43.9 | 9.9 | 78.9 | 0.113 | −0.059 | |
| 2nd codon position | 5496 | 38.7 | 14.4 | 37.2 | 9.7 | 75.9 | −0.02 | −0.194 | |
| 3rd codon position | 5496 | 42.8 | 9.8 | 42.3 | 5.1 | 85.1 | −0.006 | −0.318 | |
| tRNAs | 1458 | 39.7 | 8.6 | 40.2 | 11.5 | 79.9 | 0.006 | 0.147 | |
| rRNAs | 2123 | 42.7 | 5.9 | 40 | 11.4 | 82.7 | −0.033 | 0.319 | |
| Control region | 1534 | 45.7 | 5.7 | 45.2 | 3.5 | 90.9 | −0.006 | −0.243 | |
| Loxocera sinica | Whole genome | 16,527 | 38.8 | 11.8 | 41.2 | 8.3 | 80 | 0.031 | −0.175 |
| PCGs | 11,183 | 44.3 | 11.1 | 33.2 | 11.4 | 77.5 | −0.143 | 0.013 | |
| 1st codon position | 5509 | 38.4 | 10.4 | 42 | 9.1 | 80.5 | 0.045 | −0.064 | |
| 2nd codon position | 5509 | 37.2 | 14.4 | 38.7 | 9.8 | 75.9 | 0.02 | −0.189 | |
| 3rd codon position | 5509 | 40.7 | 10.5 | 43 | 5.8 | 83.6 | 0.027 | −0.285 | |
| tRNAs | 1455 | 39.7 | 8.7 | 40.3 | 11.3 | 80.1 | 0.008 | 0.131 | |
| rRNAs | 2125 | 42.9 | 5.9 | 40.1 | 11.1 | 83 | −0.034 | 0.308 | |
| Control region | 1585 | 44.5 | 5.2 | 46.9 | 3.3 | 91.5 | 0.026 | −0.215 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Yang, D. Mitochondrial Genomes Provide New Phylogenetic and Evolutionary Insights into Psilidae (Diptera: Brachycera). Insects 2022, 13, 518. https://doi.org/10.3390/insects13060518
Zhou J, Yang D. Mitochondrial Genomes Provide New Phylogenetic and Evolutionary Insights into Psilidae (Diptera: Brachycera). Insects. 2022; 13(6):518. https://doi.org/10.3390/insects13060518
Chicago/Turabian StyleZhou, Jiale, and Ding Yang. 2022. "Mitochondrial Genomes Provide New Phylogenetic and Evolutionary Insights into Psilidae (Diptera: Brachycera)" Insects 13, no. 6: 518. https://doi.org/10.3390/insects13060518
APA StyleZhou, J., & Yang, D. (2022). Mitochondrial Genomes Provide New Phylogenetic and Evolutionary Insights into Psilidae (Diptera: Brachycera). Insects, 13(6), 518. https://doi.org/10.3390/insects13060518