
Functions of Egg-Coating Substances Secreted by Female Accessory Glands in Alderflies, Fishflies and Dobsonflies (Megaloptera)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Examined Species
2.2. Female Accessory Gland Substances
2.3. Statistics
3. Results
3.1. Egg Masses and Accessory Gland Substances
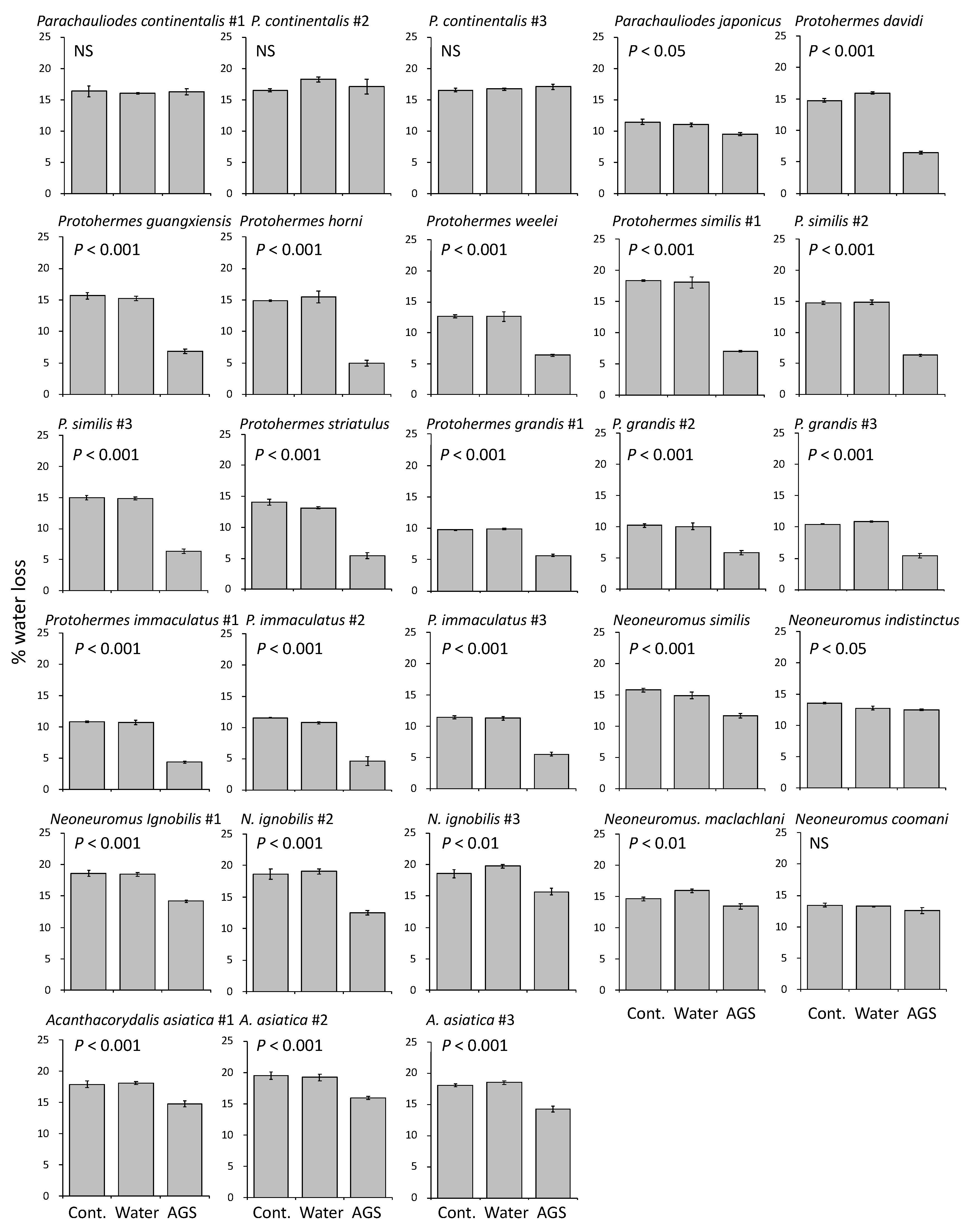
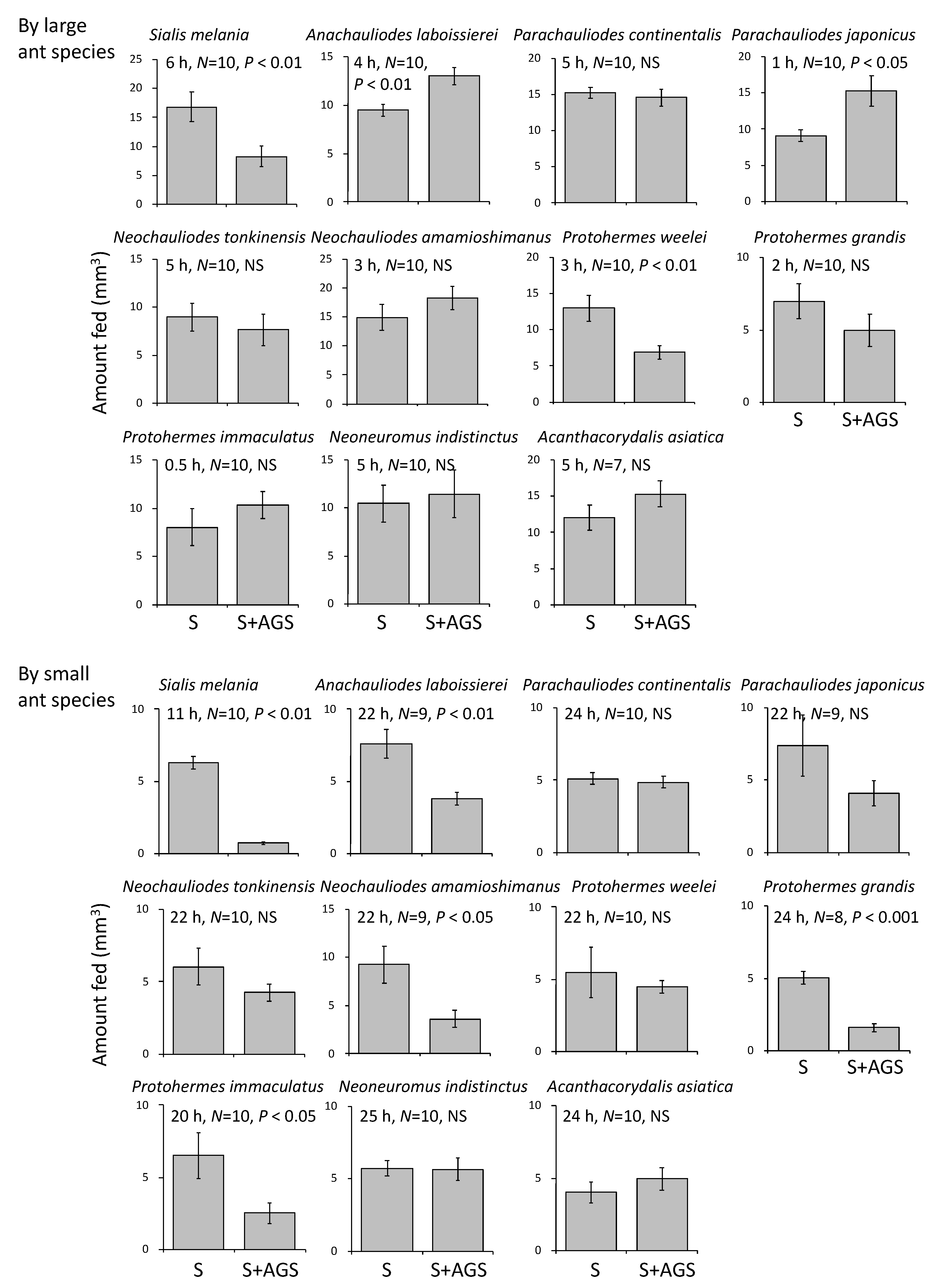
3.2. Functions of Accessory Gland Substances
4. Discussion
4.1. Functions of Egg-Coating Substances
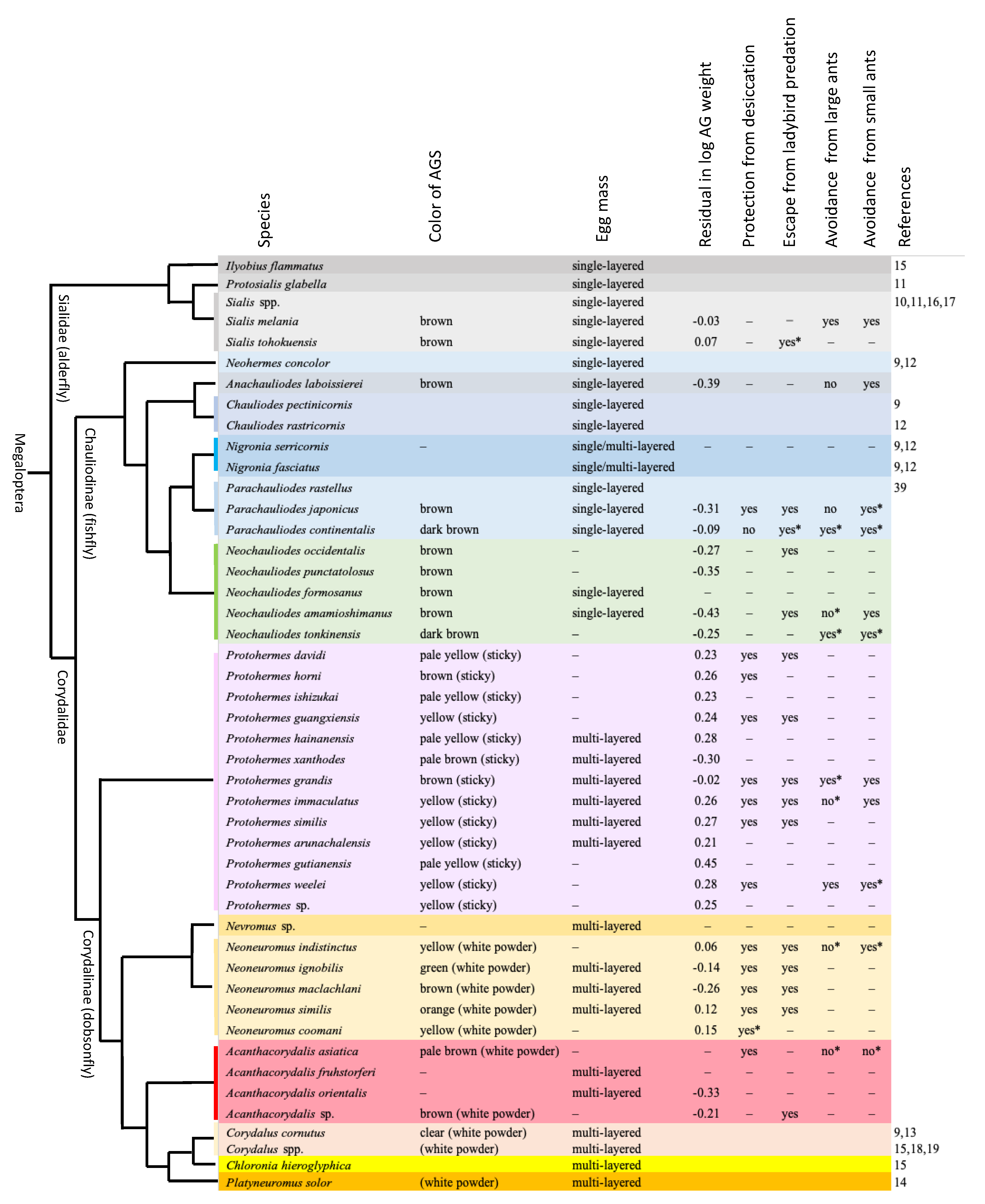
4.2. Evolutionary Patterns of Egg Mass Characteristics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trougakos, I.P.; Margaritis, L.H. Novel morphological and physiological aspects of insect eggs. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell: Oxford, UK, 2002; pp. 3–36. [Google Scholar]
- Blum, M.S.; Hilker, M. Chemical protection of insect eggs. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell: Oxford, UK, 2002; pp. 61–90. [Google Scholar]
- Gillott, C. Insect accessory reproductive glands: Key players in production and protection. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell: Oxford, UK, 2002; pp. 37–59. [Google Scholar]
- Marchini, D.; Marri, L.; Rosetto, M.; Manetti, A.G.; Dallai, R. Presence of antibacterial peptides on the laid egg chorion of the medfly Ceratitis capitata. Biochem. Biophys. Res. Commun. 1997, 240, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Boos, S.; Meunier, J.; Pichon, S.; Kölliker, M. Maternal care provides antifungal protection to eggs in the European earwig. Behav. Ecol. 2014, 25, 754–761. [Google Scholar] [CrossRef]
- Makover, V.; Ronen, Z.; Lubin, Y.; Khalaila, I. Eggshell spheres protect brown widow spider (Latrodectus geometricus) eggs from bacterial infection. J. R. Soc. Interface 2019, 16, 20180581. [Google Scholar] [CrossRef] [PubMed]
- Hinton, H.E. Biology of Insect Eggs; Pergamon Press: Oxford, UK, 1981; Volume I–III. [Google Scholar]
- Eisner, T.; Rossini, C.; González, A.; Iyengar, V.K.; Siegler, M.V.; Smedley, S.R. Paternal investment in egg defence. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell: Oxford, UK, 2002; pp. 91–116. [Google Scholar]
- Baker, J.R.; Neunzig, H.H. The egg masses, eggs, and first-instar larvae of Eastern North American Corydalidae. Ann. Entomol. Soc. Am. 1968, 61, 1181–1187. [Google Scholar] [CrossRef]
- Azam, K.M.; Anderson, N.H. Life history and habits of Sialis rotunda and S. californica in western Oregon. Ann. Entomol. Soc. Am. 1969, 62, 549–558. [Google Scholar] [CrossRef]
- Canterbury, L.E.; Neff, S.E. Eggs of Sialis (Sialidae: Megaloptera) in eastern North America. Can. Entomol. 1980, 112, 409–419. [Google Scholar] [CrossRef]
- Arnold, D.C.; Drew, W.A. A preliminary survey of the Megaloptera of Oklahoma. Proc. Okla. Acad. Sci. 1987, 67, 23–26. [Google Scholar]
- Mangan, B.P. Oviposition of the dobsonfly (Corydalus cornutus, Megaloptera) on a large river. Am. Midl. Nat. 1992, 127, 348–354. [Google Scholar] [CrossRef]
- Contreras-Ramos, A. Mating behavior of Platyneuromus (Megaloptera: Corydalidae), with life history notes on dobsonflies from Mexico and Costa Rica. Entomol. News 1999, 110, 125–135. [Google Scholar]
- Azevêdo, C.A.S. Taxonomia e Bionomia de Imaturos de Megaloptera (Insecta) na Amazônia Central, Brasil. Master’s Thesis, Instituto de Pesquisas da Amazônia, Manaus, Brazil, 2003. [Google Scholar]
- Takeuchi, Y. Egg-laying sites of the alderfly Sialis yamatoensis (Megaloptera: Sialidae) around ponds in Hyogo Prefecture, western Japan. Hyogo Freshw. Biol. 2005, 56, 129–135. (In Japanese) [Google Scholar]
- Takeuchi, Y. The life history of the alderfly Sialis japonica (Megaloptera: Sialidae) in Ayukaeri River in Hyogo Prefecture, western Japan. Hyogo Freshw. Biol. 2007, 59, 79–109. (In Japanese) [Google Scholar]
- Melo, A.S.; Wheeler, T.A. A new species of Pseudogaurax Malloch (Diptera: Chloropidae) reared from dobsonfly egg-masses (Megaloptera: Corydalidae) in Brazil. Zootaxa 2009, 1972, 53–58. [Google Scholar] [CrossRef]
- Rivera-Gasperín, S.L.; Ardila-Camacho, A.; Contreras-Ramos, A. Bionomics and ecological services of Megaloptera larvae (dobsonflies, fishflies, alderflies). Insects 2019, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, G.; Leischner, T.G. The life history and feeding habits of Sialis cornuta Ross in a series of abandoned beaver ponds (Insecta; Megaloptera). Can. J. Zool. 1973, 51, 121–131. [Google Scholar] [CrossRef]
- Brown, A.V.; Fitzpatrick, L.C. Life history and population energetics of the dobsonfly, Corydalus cornutus. Ecology 1978, 59, 1091–1108. [Google Scholar] [CrossRef]
- Way, M.J.; Khoo, K.C. Role of ants in pest management. Annu. Rev. Entomol. 1992, 37, 479–503. [Google Scholar] [CrossRef]
- Lindstedt, C.; Boncoraglio, G.; Cotter, S.; Gilbert, J.; Kilner, R.M. Aposematism in the burying beetle? Dual function of anal fluid in parental care and chemical defense. Behav. Ecol. 2017, 28, 1414–1422. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Yamane, S. Food habits of some ant species inhabiting the Taisho lava of the Sakura-jima volcano, Japan (Hymenoptera, Formicidae). Rep. Fac. Sci. Kagoshima Univ. (Earth Sci. Biol.) 1990, 23, 9–22. (In Japanese) [Google Scholar]
- Butin, E.E.; Havill, N.P.; Elkinton, J.S.; Montgomery, M.E. Feeding preference of three lady beetle predators of the hemlock woolly adelgid (Homoptera: Adelgidae). J. Econ. Entomol. 2004, 97, 1635–1641. [Google Scholar] [CrossRef]
- Bayoumy, M.H.; Michaud, J.P. Cannibalism in two subtropical lady beetles (Coleoptera: Coccinellidae) as a function of density, life stage, and food supply. J. Insect Behav. 2015, 28, 387–402. [Google Scholar] [CrossRef]
- Guerra-Grenier, E. Evolutionary ecology of insect egg coloration: A review. Evol. Ecol. 2019, 33, 1–19. [Google Scholar] [CrossRef]
- Thorat, L.; Nath, B.B. Insects with survival kits for desiccation tolerance under extreme water deficits. Front. Physiol. 2018, 9, 1843. [Google Scholar] [CrossRef] [PubMed]
- New, T.R.; Theischinger, G. Megaloptera (alderflies, dobsonflies). Handb. Zool. IV Arthropoda Insecta 1993, 33, 1–97. [Google Scholar]
- Stamp, N.E. Egg deposition patterns in butterflies: Why do some species cluster their eggs rather than deposit them singly? Am. Nat. 1980, 115, 367–380. [Google Scholar] [CrossRef]
- Clark, B.R.; Faeth, S.H. The evolution of egg clustering in butterflies: A test of the egg desiccation hypothesis. Evol. Ecol. 1998, 12, 543–552. [Google Scholar] [CrossRef]
- Yashiro, N.; Hirose, Y.; Honda, J.Y.; Takeuchi, Y.; Yashiro, T. A new species of Trichogramma (Hymenoptera: Trichogrammatidae) parasitic on eggs of the alderfly Sialis melania (Neuroptera: Sialidae) from Japan, with comments on its phylogeny and male wing polymorphism. Entomol. Sci. 2012, 15, 189–196. [Google Scholar] [CrossRef]
- Salt, G. The egg-parasite of Sialis lutaria: A study of the influence of the host upon a dimorphic parasite. Parasitology 1937, 29, 539–553. [Google Scholar] [CrossRef]
- Martin, C.H. Biological studies of two hymenopterous parasites of aquatic insect eggs. Entomol. Am. (New Ser.) 1928, 8, 105–156. [Google Scholar]
- Zhang, X.; Zhang, Y.Z.; Wang, Y.; Chen, F.Q.; Yu, F.; Zhou, Q.S. Description of three new species of Ooencyrtus (Hymenoptera: Encyrtidae) from China. Zootaxa 2014, 3790, 451–465. [Google Scholar] [CrossRef]
- Noyes, J.S.; Hirose, Y. A new species of Ooencyrtus (Hymenoptera: Encyrtidae) from Japan, parasitic in the eggs of Protohermes grandis (Megaloptera: Corydalidae). Kontyû 1997, 65, 199–204. [Google Scholar]
- Jiang, Y.; Yue, L.; Yang, F.; Gillung, J.P.; Winterton, S.L.; Price, B.W.; Contreras-Ramos, A.; Hayashi, F.; Aspöck, U.; Aspöck, H.; et al. Similar pattern, different paths: Tracing the biogeographical history of Megaloptera (Insecta: Neuropterida) using mitochondrial phylogenomics. Cladistics 2022, 38, 374–391. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F. Rapid evacuation of spermatophore contents and male post-mating behavior in alderflies (Megaloptera: Sialidae). Entomol. Sci. 1999, 2, 49–56. [Google Scholar]
- Shimonoya, M. A new species of the genus Parachauliodes (Megaloptera: Corydalidae: Chauliodinae) from Northern Kyushu, Japan. Zootaxa 2019, 4585, 151–167. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family (Subfamily) Species | Site | Adult State | No. of Females | Head Width (mm) | Forewing Length (mm) | AG Weight (mg) | |||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | ||||
| Sialidae | |||||||||
| Sialis melania Nakahara | Japan | Field-collected | 14 | 2.74 | 0.05 | 15.50 | 0.19 | 0.5 | 0.0 |
| Sialis tohokuensis Hayashi & Suda | Japan | Field-collected | 4 | 2.89 | 0.03 | 17.15 | 0.32 | 0.7 | 0.1 |
| Corydalidae (Chauliodinae) | |||||||||
| Anachauliodes laboissierei (Navás) | China | Field-collected | 2 | 5.73 | 0.03 | 51.65 | 0.45 | 6.0 | 0.0 |
| Nigronia serricornis (Say) | USA | Reared | 1 | – | – | – | |||
| Parachauliodes japonicus (McLachlan) | Japan | Reared | 15 | 6.25 | 0.08 | 44.23 | 0.66 | 10.9 | 1.6 |
| Parachauliodes continentalis (Weele) | Japan | Field-collected/Reared | 44 | 6.13 | 0.04 | 52.76 | 0.27 | 16.4 | 1.1 |
| Neochauliodes occidentalis Weele | China | Reared | 9 | 5.47 | 0.16 | 43.16 | 1.55 | 6.4 | 1.5 |
| Neochauliodes punctatolosus Liu & Yang | China | Field-collected | 2 | 4.84 | 0.01 | 40.47 | 0.64 | 3.1 | 0.5 |
| Neochauliodes formosanus (Okamoto) | China | reared | 1 | – | – | – | |||
| Neochauliodes amamioshimanus Liu, Hayashi & Yang | Japan | Reared | 13 | 4.60 | 0.06 | 36.05 | 0.43 | 2.0 | 0.4 |
| Neochauliodes tonkinensis (Weele) | China | Field-collected | 4 | 5.10 | 0.09 | 45.11 | 1.05 | 4.8 | 1.8 |
| Corydalidae (Corydalinae) | |||||||||
| Protohermes davidi Weele | China | Reared | 5 | 7.53 | 0.13 | 51.49 | 1.92 | 89.4 | 17.5 |
| Protohermes horni Navás | China | Reared | 2 | 8.28 | 0.35 | 58.56 | 149.4 | 36.8 | |
| Protohermes ishizukai Liu, Hayashi & Yang | China | Field-collected | 1 | 6.02 | 42.85 | 31.6 | |||
| Protohermes guangxiensis Yang & Yang | China | Reared | 8 | 7.26 | 0.10 | 50.51 | 0.67 | 78.1 | 5.9 |
| Protohermes hainanensis Yang & Yang | China | Field-collected | 1 | 6.59 | 48.24 | 54.4 | |||
| Protohermes xanthodes Navás | China | Field-collected | 1 | 6.00 | 44.23 | 9.1 | |||
| Protohermes grandis (Thunberg) | Japan | Reared/Field-collected | 18 | 6.79 | 0.09 | 48.62 | 0.69 | 31.3 | 4.3 |
| Protohermes immaculatus Kuwayama | Japan | Reared | 27 | 5.27 | 0.04 | 35.79 | 0.33 | 18.1 | 1.6 |
| Protohermes similis Yang & Yang | China | Reared | 18 | 6.52 | 0.06 | 45.08 | 0.59 | 49.6 | 4.4 |
| Protohermes arunachalensis Ghosh | China | Field-collected | 1 | 5.77 | 47.37 | 24.8 | |||
| Protohermes gutianensis Yang & Yang | China | Field-collected | 1 | 5.88 | 42.53 | 47.0 | |||
| Protohermes weelei Navás | China | Reared | 4 | 5.96 | 0.09 | 42.14 | 1.52 | 34.3 | 3.5 |
| Protohermes sp. | China | Reared | 2 | 5.90 | 0.60 | 41.83 | 4.18 | 30.5 | 13.50 |
| Nevromus sp. | Vietnam | Reared | 1 | – | – | – | |||
| Neoneuromus indistinctus Liu, Hayashi & Yang | China | Field-collected/Reared | 3 | 8.54 | 0.16 | 49.85 | 0.12 | 106.9 | 35.1 |
| Neoneuromus ignobilis Navás | China | Field-collected | 46 | 10.07 | 0.10 | 60.86 | 0.51 | 145.2 | 17.7 |
| Neoneuromus maclachlani (Weele) | China | Reared | 3 | 10.46 | 0.35 | 59.10 | 133.9 | 25.6 | |
| Neoneuromus similis Liu, Hayashi & Yang | China | Field-collected/Reared | 2 | 11.14 | 0.00 | 67.44 | 3.04 | 426.3 | 221.6 |
| Neoneuromus coomani Lestage | China | Reared | 1 | 8.10 | 54.00 | 105.0 | |||
| Acanthacorydalis asiatica (Wood-Mason) | Vietnam | Field-collected | 8 | – | – | – | |||
| Acanthacorydalis fruhstorferi Weele | China | Field-collected | 1 | – | – | – | |||
| Acanthacorydalis orientalis (McLachlan) | China | Field-collected | 1 | 10.49 | 68.29 | 114.1 | |||
| Acanthacorydalis sp. | China | Field-collected | 1 | 12.26 | 75.55 | 309.3 | |||
| Family | Subfamily | Species | Egg Treatment | Intact | Broken | Eaten | Total | p in χ2-Tests |
|---|---|---|---|---|---|---|---|---|
| Sialidae | Sialis tohokuensis | Egsg without AGS | 10 | 8 | 2 | 20 | 0.827 | |
| Eggs with AGS | 11 | 8 | 1 | 20 | ||||
| Corydalidae | Chauliodinae | Parachauliodes continentalis | Egsg without AGS | 6 | 7 | 47 | 60 | 0.436 |
| Eggs with AGS | 10 | 9 | 41 | 60 | ||||
| Parachauliodes japonicus | Eggs without AGS | 23 | 33 | 39 | 95 | <0.001 | ||
| Eggs with AGS | 19 | 59 | 17 | 95 | ||||
| Neochauliodes amamioshimanus | Eggs without AGS | 23 | 1 | 36 | 60 | 0.014 | ||
| Eggs with AGS | 38 | 2 | 20 | 60 | ||||
| Neochauliodes occidentalis | Eggs without AGS | 25 | 7 | 18 | 50 | <0.001 | ||
| Eggs with AGS | 43 | 6 | 1 | 50 | ||||
| Corydalinae | Protohermes davidi | Eggs without AGS | 17 | 7 | 21 | 45 | <0.001 | |
| Eggs with AGS | 45 | 0 | 0 | 45 | ||||
| Protohermes guangxiensis | Eggs without AGS | 19 | 3 | 33 | 55 | <0.001 | ||
| Eggs with AGS | 51 | 4 | 0 | 55 | ||||
| Protohermes similis | Eggs without AGS | 7 | 1 | 27 | 35 | <0.001 | ||
| Eggs with AGS | 33 | 2 | 0 | 35 | ||||
| Protohermes grandis | Eggs without AGS | 34 | 9 | 62 | 105 | <0.001 | ||
| Eggs with AGS | 60 | 19 | 26 | 105 | ||||
| Protohermes immaculatus | Eggs without AGS | 35 | 10 | 35 | 80 | <0.001 | ||
| Eggs with AGS | 53 | 14 | 13 | 80 | ||||
| Neoneuromus similis | Eggs without AGS | 8 | 3 | 29 | 40 | <0.001 | ||
| Eggs with AGS | 39 | 0 | 1 | 40 | ||||
| Neoneuromus ignobilis | Eggs without AGS | 17 | 6 | 22 | 45 | <0.001 | ||
| Eggs with AGS | 42 | 0 | 3 | 45 | ||||
| Neoneuromus indistinctus | Eggs without AGS | 3 | 1 | 46 | 50 | <0.001 | ||
| Eggs with AGS | 45 | 4 | 1 | 50 | ||||
| Neoneuromus maclachlani | Eggs without AGS | 8 | 3 | 29 | 40 | <0.001 | ||
| Eggs with AGS | 34 | 0 | 6 | 40 | ||||
| Acanthacorydalis sp. | Eggs without AGS | 5 | 4 | 26 | 35 | <0.001 | ||
| Eggs with AGS | 31 | 3 | 1 | 35 |
| Host | Parasitoid | Locality | % Eggs Parasitized | Reference | |
|---|---|---|---|---|---|
| N | Mean (Range) | ||||
| Sialidae (alderfly) | |||||
| Sialis lutaria (Linnaeus) | Trichogramma semblidis (Aurivillius) | Sweden, England | [33] | ||
| Sialis infumata Newman | Trichogramma semblidis (Aurivillius) | North America | [34] | ||
| Sialis californica Banks | Trichogramma semblidis (Aurivillius) | North America | ? | (37–65) | [10] |
| Sialis rotunda Banks | Trichogramma semblidis (Aurivillius) | North America | ? | around 14 | [10] |
| Sialis cornuta Ross | Trichogramma semblidis (Aurivillius) | North America | 12 | 58.1 a | [20] |
| Sialis cornuta Ross | Trichogramma semblidis (Aurivillius) | North America | 5 | 77.7 b | [20] |
| Sialis melania Nakahara | Trichogramma tajimaense Yashiro, Hirose & Honda | Japan (Hyogo) | 4 | 56.2 (28.9–94.3) | [32] |
| Corydalidae: Corydalinae (dobsonfly) | |||||
| Protohermes sp. | Ooencyrtus longicauda Zhang & Zhang | China (Hunan) | 1 | 6.6 | [35] |
| Protohermes xanthodes Navás | Ooencyrtus protohermesis Zhang & Zhang | China (Beijing) | 1 | 6.8 | [35] |
| Protohermes grandis Thunberg | Ooencyrtus yoshidai Noyes & Hirose | Japan (Nagano) | [36] | ||
| Protohermes grandis Thunberg | Ooencyrtus yoshidai Noyes & Hirose | Japan (Niigata) | 1 | 16.2 | This study |
| Protohermes grandis Thunberg | Ooencyrtus yoshidai Noyes & Hirose | Japan (Sado Island) | 1 | 28.1 | This study |
| Corydalus spp. | Pseudogaurax idiogenes Wheeler c | Brazil (Sao Paulo) | not so high | [18] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, P.; Liu, X.; Hayashi, F. Functions of Egg-Coating Substances Secreted by Female Accessory Glands in Alderflies, Fishflies and Dobsonflies (Megaloptera). Insects 2022, 13, 766. https://doi.org/10.3390/insects13090766
Yu P, Liu X, Hayashi F. Functions of Egg-Coating Substances Secreted by Female Accessory Glands in Alderflies, Fishflies and Dobsonflies (Megaloptera). Insects. 2022; 13(9):766. https://doi.org/10.3390/insects13090766
Chicago/Turabian StyleYu, Pei, Xingyue Liu, and Fumio Hayashi. 2022. "Functions of Egg-Coating Substances Secreted by Female Accessory Glands in Alderflies, Fishflies and Dobsonflies (Megaloptera)" Insects 13, no. 9: 766. https://doi.org/10.3390/insects13090766