
Entomological Characteristics of Malaria Transmission across Benin: An Essential Element for Improved Deployment of Vector Control Interventions

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
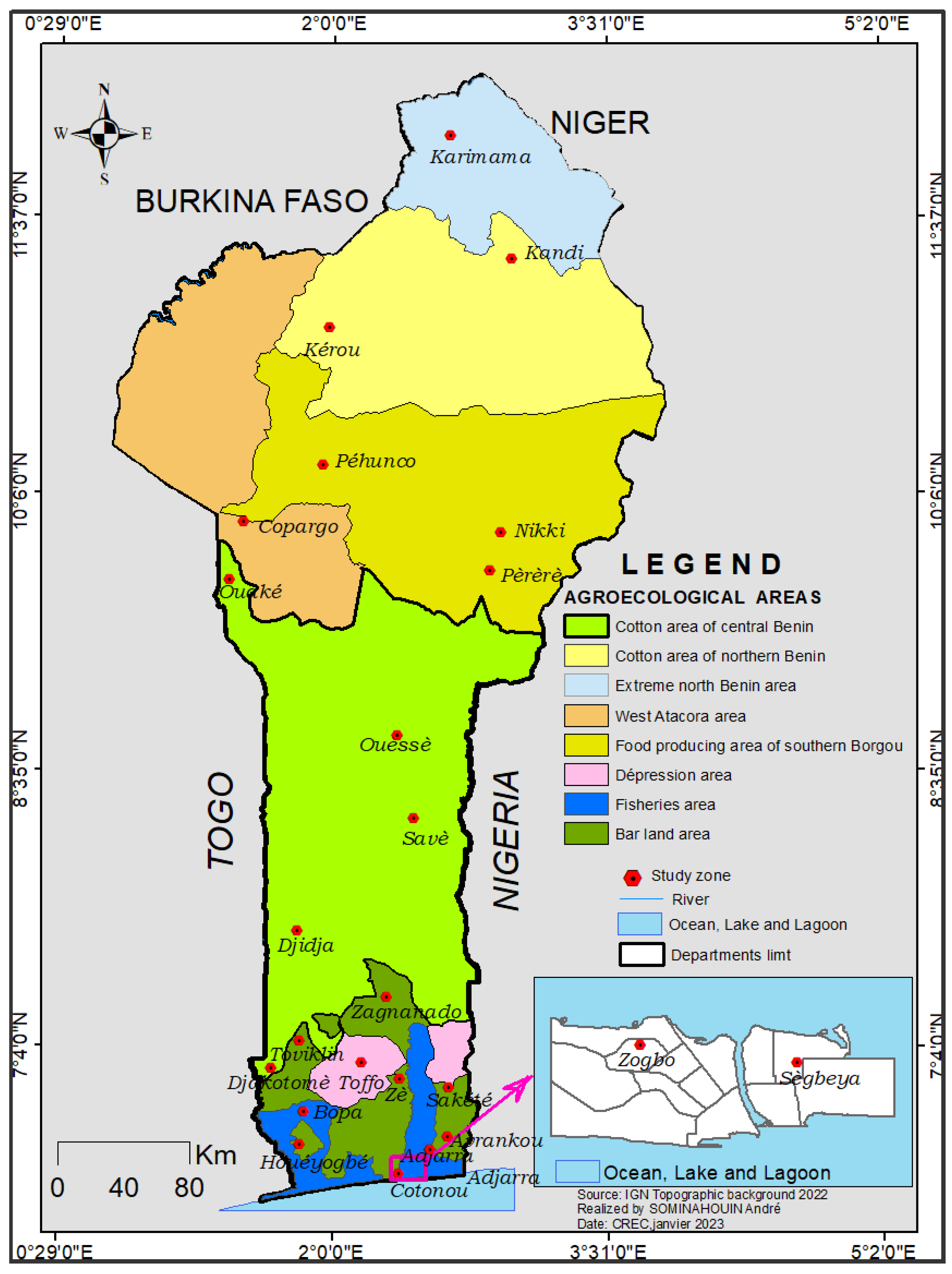
2.1. Study Sites
2.2. Monitoring of Anopheles Population Dynamics, Vector Species and Malaria Transmission
2.3. Laboratory Analysis
2.4. Elaboration of the Entomological Transmission Profile and Collection of Malaria Incidence and Prevalence Data
2.5. Data Analysis
3. Results
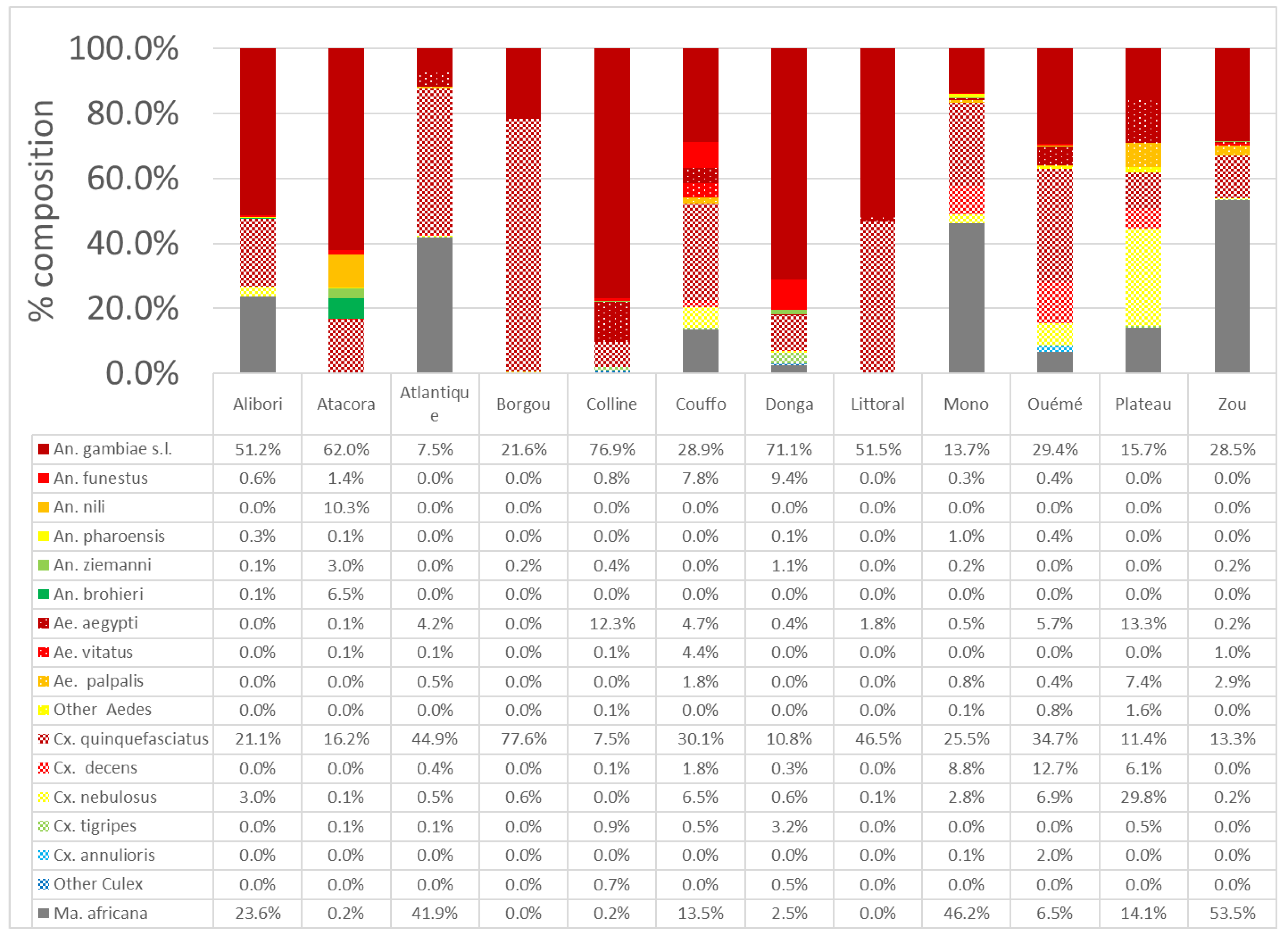
3.1. Mosquito Species Diversity and Molecular Identification of Related Species of the Anopheles Gambiae Complex
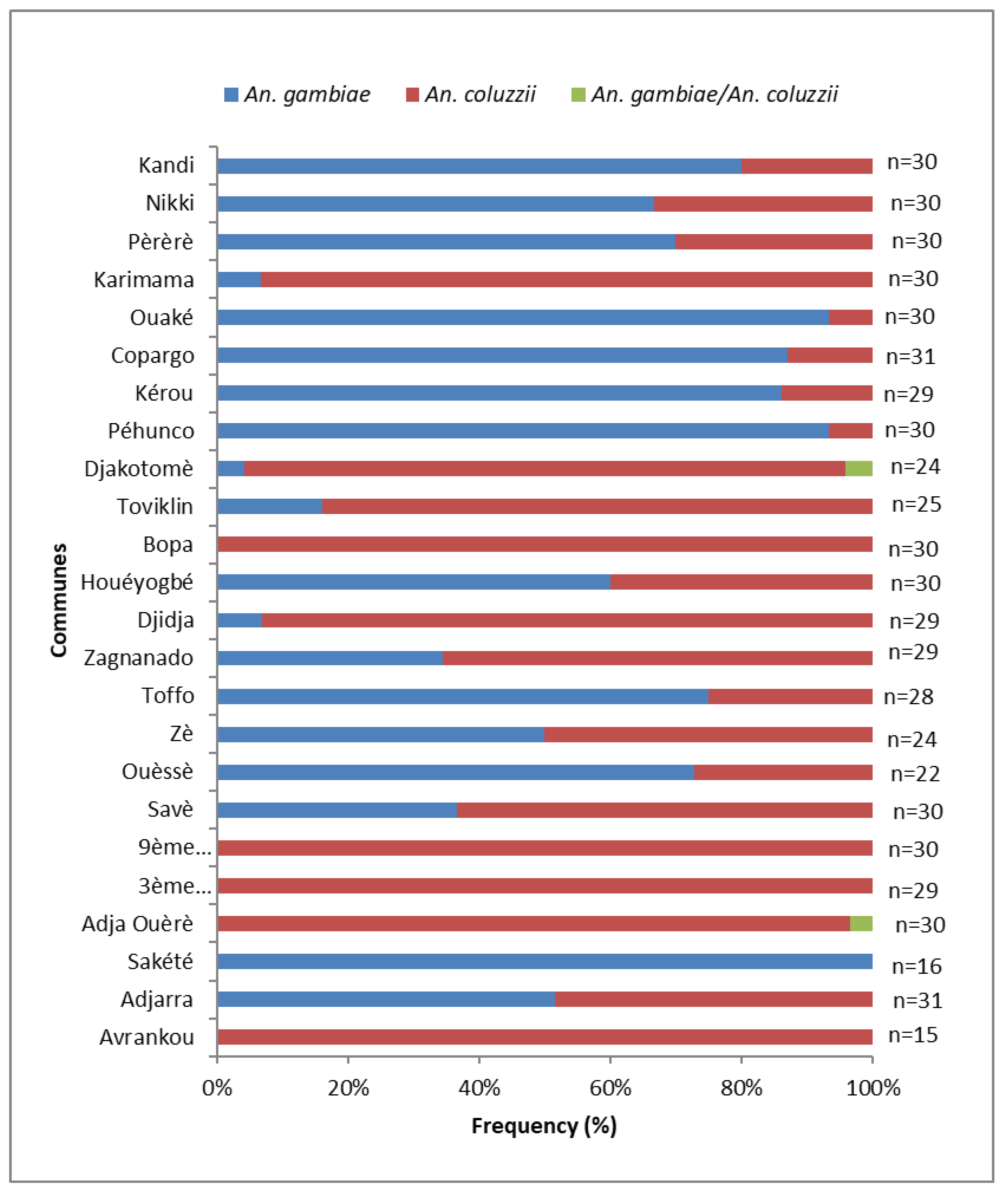
3.2. Species and Allelic Frequency of L1014F kdr and G119S Ace-1 Mutations in An. gambiae s.l. Populations Involved in Malaria Transmission in Each Commune
3.3. Anopheles Density and Biting Behavior by Commune and Department
3.4. An. gambiae Sensu Lato Density and Biting Behavior Indoors and Outdoors
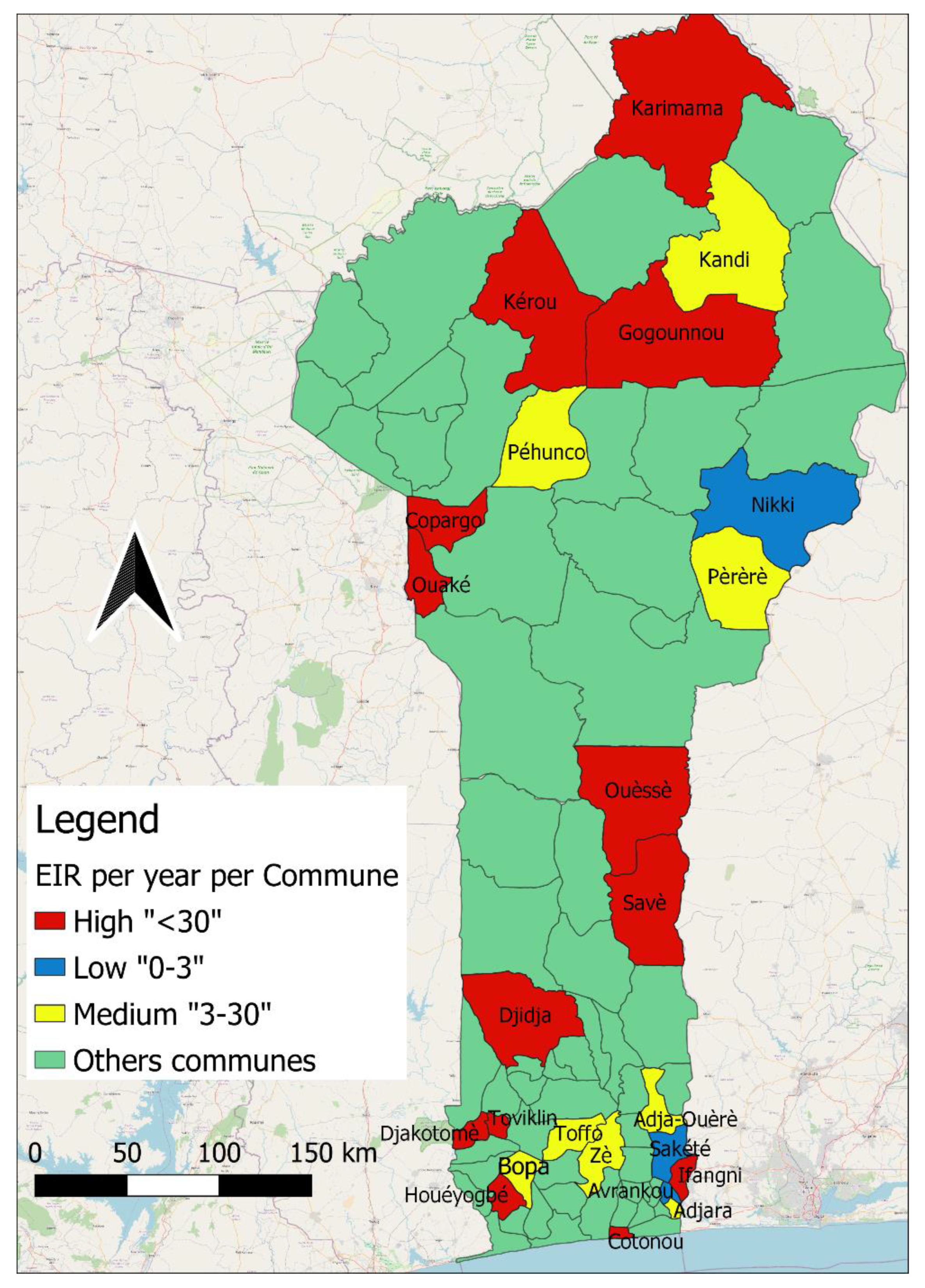
3.5. Sporozoite Index and Entomological Inoculation Rate (EIR) by Commune and by Department
3.6. Entomological Profile of Malaria Transmission across Benin
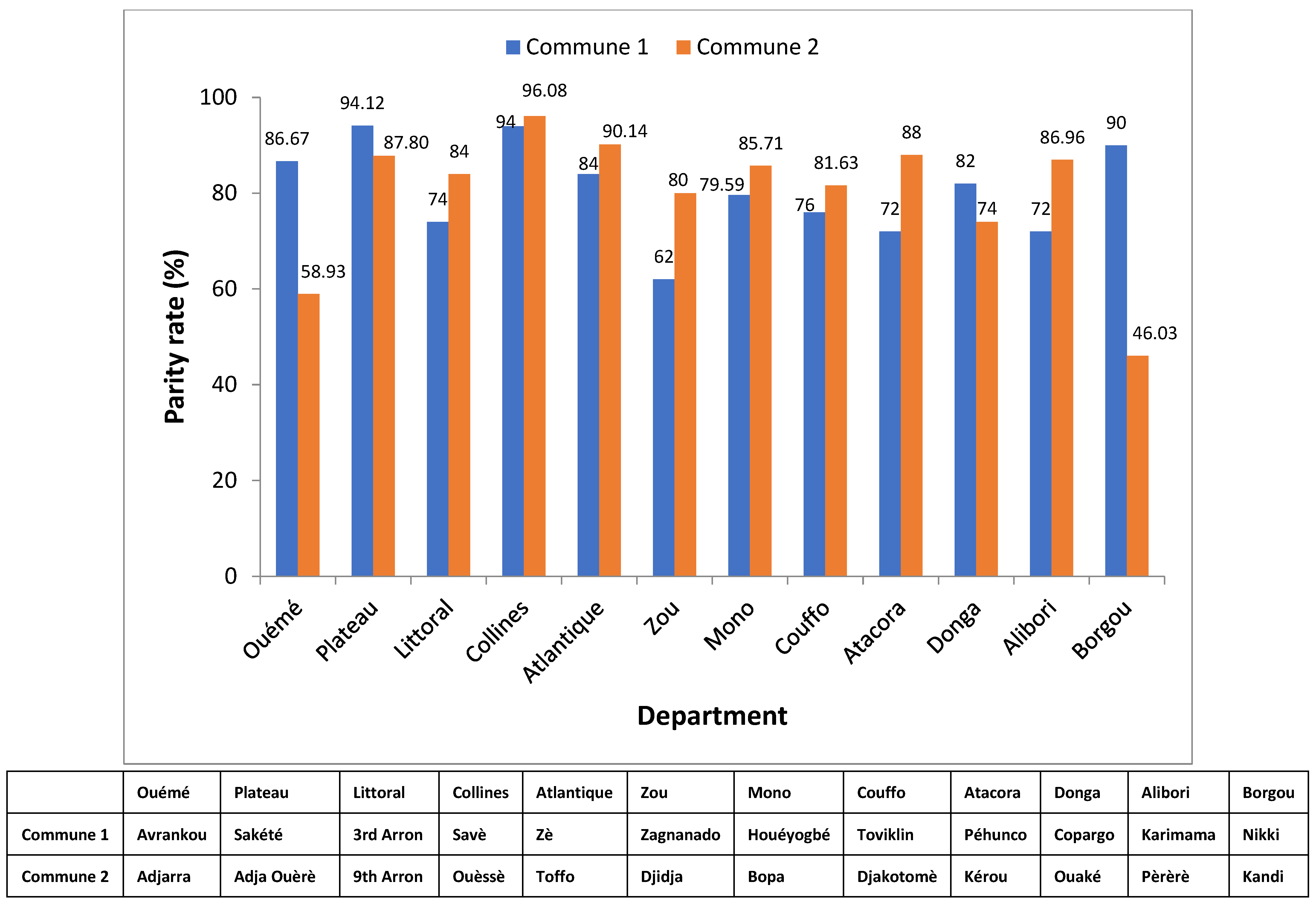
3.7. Parity rate of An. gambiae s.l., An. funestus Group and An. nili Group
3.8. An. gambiae s.l Blood Meal Origins
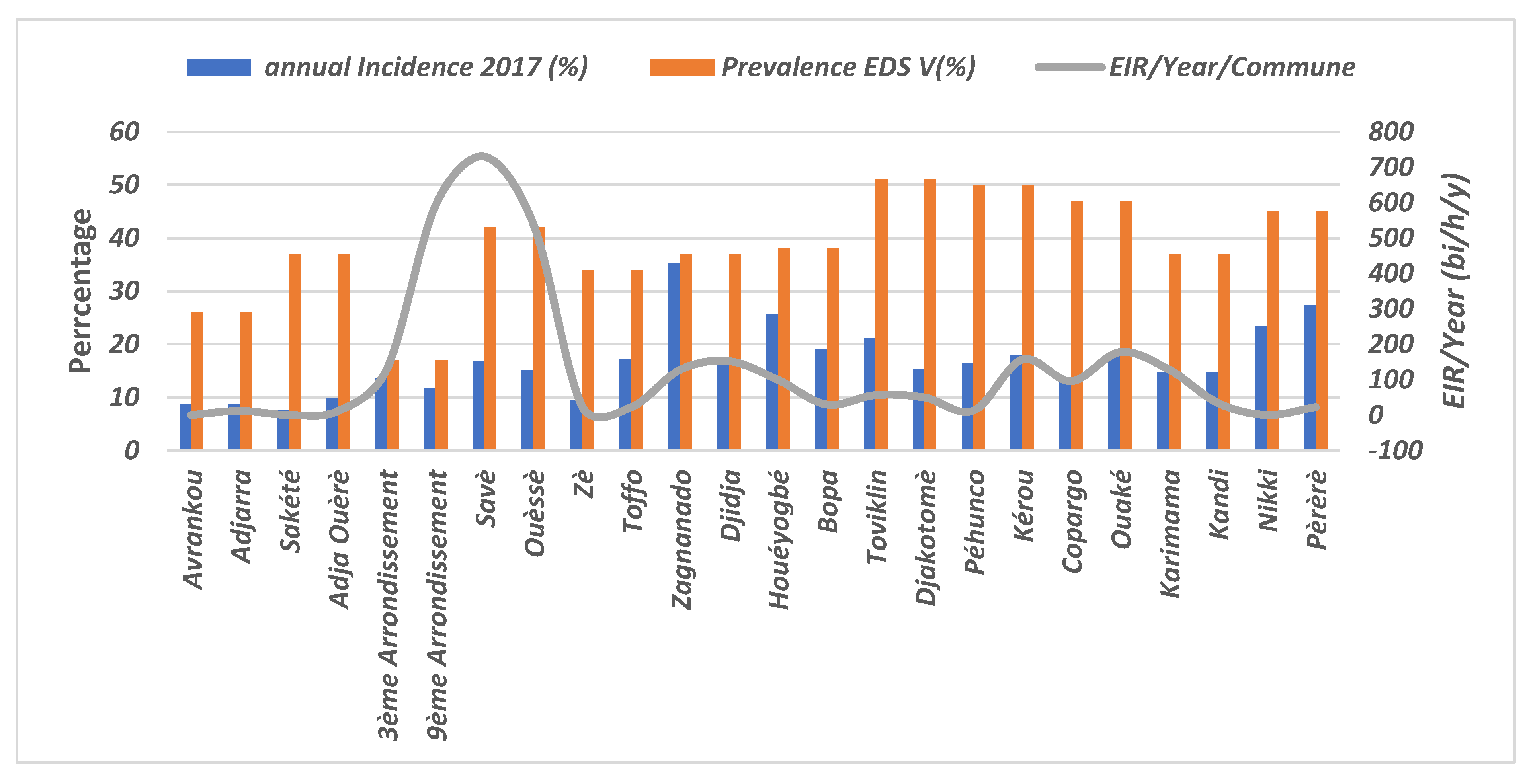
3.9. Evaluation of the EIR According to Malaria Incidence and Prevalence in the Communes
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, E.H.; Paskewitz, S.M. Malaria: Current and future prospects for control. Annu. Rev. Entomol. 1995, 4O, 195–219. [Google Scholar] [CrossRef]
- Robert, S.; Guerra, C.; Noor, A.; Myint, H.; Hay, S. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 2005, 434, 214–217. [Google Scholar]
- WHO. Malaria World Report; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- WHO. Global Report on Insecticide Resistance in Malaria Vectors: 2010–2016; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Kazembe, L.N.; Kleinschmidt, I.; Holtz, T.H. Spatial analysis and mapping of malaria risk in Malawi using point-referenced prevalence of infection data. Int. J. Health Geograph. 2006, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaria, R.B. The Global Malaria Action Plan 2012. 2013. Available online: http://www.rollbackmalaria.org/gmap (accessed on 26 February 2018).
- Antonio-Nkondjio, C.; Awono-Ambene, P.; Toto, J.-C.; Meunier, J.-Y.; Zebaze-Kemleu, S.; Nyambam, R.; Wondji, C.S.; Tchuinkam, T.; Fontenille, D. High malaria transmission intensity in a village close to Yaounde, the capital city of Cameroon. J. Med. Entomol. 2002, 39, 350–355. [Google Scholar] [CrossRef] [PubMed]
- White, N.J. Plasmodium knowlesi: The fifth human malaria parasite. Clin. Infect. Dis. 2008, 46, 172–173. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, P.; Robert, V. Les Anophèles, Biologie, Transmission du Paludisme et Lutte Antivectorielle; IRD Montpellier France Editions: Montpellier, France, 2009. [Google Scholar]
- Fontenille, D.; Simard, F. Unravelling complexities in human malaria transmission dynamics in Africa through a comprehensive knowledge of vector populations. Comp. Immun. Microbiol. Infect. Dis. 2004, 27, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Ossè, R.; Tokponnon, F.; Padonou Ge Sidick, A.; Aïkpon, R.; Fassinou, A.; Koukpo, C.; Sèwadé, W.; Akinro, B.; Sovi, A.; Aïssi, M.; et al. Involvement of Anopheles nili in Plasmodium falciparum transmission in North Benin. Malar. J. 2019, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Chatlod, L.R.; Kumar, S.; Toppo, S.; Haque, N.; Rahman, H. Nutritional evaluation of NB-21 hybrid Napier grass for goats. Indian J. Small Rum. 2012, 18, 261–263. [Google Scholar]
- WHO. World Malaria Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Moiroux, N.; Gomez, M.B.; Pennetier, C.; Elanga, E.; Djènontin, A.; Chandre, F.; Djègbé, I.; Guis, H.; Corbel, V. Changes in Anopheles funestus biting behavior following universal coverage of long-lasting insecticidal nets in Benin. J. Infect. Dis. 2012, 206, 1622–1629. [Google Scholar] [CrossRef] [Green Version]
- Ministère de la santé du Bénin. Annuaire des Statistiques Sanitaires 2018 des Départements du Benin; Ministère de la santé du Bénin: Cotonou, Benin, 2019. [Google Scholar]
- Institut National de la Statistique et de l’Analyse Économique (INSAE) et ICF. 2018. Enquête Démographique et de Santé au Bénin, 2017–2018: Indicateurs Clés. Cotonou, Bénin et Rockville, Maryland, USA: INSAE et ICF. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwij7sLAn7D8AhXJr1YBHXk-BowQFnoECBgQAQ&url=https%3A%2F%2Finstad.bj%2Fimages%2Fdocs%2Finsae-statistiques%2Fsociales%2FSante%2FEnqu%25C3%25AAte%2520D%25C3%25A9mographique%2520et%2520de%2520Sant%25C3%25A9%2520au%2520B%25C3%25A9nin%2520 (accessed on 5 January 2020).
- Djènontin, A.; Bio-Bangana, S.; Moiroux, N.; Henry, M.-C.; Bousari, O.; Chabi, J.; Ossè, R.; Koudénoukpo, S.; Corbel, V.; Akogbeto, M.; et al. Culicidae diversity, malaria transmission and insecticide resistance alleles in malaria vectors in Ouidah–Kpomasse–Tori district from Benin (West Africa): A pre-intervention study. Parasite Vectors 2010, 3, 83. [Google Scholar] [CrossRef] [Green Version]
- Gnanguenon, V.; Govoétchan, R.; Agossa, F.R.; Ossè, R.; Oke-Agbo, F.; Azondekon, R.; Sovi, A.; Attolou, R.; Badirou, K.; Tokponnon, F.T.; et al. Transmission patterns of Plasmodium falciparum by Anopheles gambiae in Benin. Malar. J. 2014, 13, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akogbéto, M.C.; Aïkpon, R.Y.; Azondekon, R.; Padonou, G.G.; Ossè, R.A.; Agossa, F.R.; Beach, R.; Sèzonlin, M. Six years of experience in entomological surveillance of indoor residual spraying against malaria transmission in Benin: Lessons learned, challenges and outlooks. Malar. J. 2015, 14, 242. [Google Scholar] [CrossRef] [PubMed]
- Tokponnon, F.T.; Sovi, A.; Ossè, R.; Oke, M.; Gazard, D.; Akogbéto, M.C. Vector resistance to pyrethroids and impact on the efficacy of insecticide-treated mosquito nets in Southern Benin. JPVB 2018, 2, 0316. [Google Scholar]
- PNLP-Bénin. Plan Stratégique National de Lutte contre le paludisme 2017–2021; PNLP: Cotonou, Benin, 2017. [Google Scholar]
- Akogbéto, M. Lagoonal and coastal malaria at Cotonou: Entomological findings. Santé 2000, 10, 267–275. [Google Scholar] [PubMed]
- Padonou, G.G.; Sezonlin, M.; Gbedjissi, G.L.; Ayi, I.; Azondekon, R.; Djènontin, A.; Oké-Agbo, F. Biology of Anopheles gambiae and insecticide resistance: Entomological study for a large scale of indoor residual spraying in South East. J. Parasitol. Vector Biol. 2011, 3, 59–68. [Google Scholar]
- Ossè, R.A.; Aïkpon, R.; Gbédjissi, G.L.; Gnanguenon, V.; Sèzonlin, M.; Govoétchan, R.; Sovi, A.; Oussou, O. A shift from indoor residual spraying (IRS) with bendiocarb to Long-Lasting Insecticidal (mosquito) Nets (LLINs) associated with changes in malaria transmission indicators in pyrethroid resistance areas in Benin. Parasites Vectors 2013, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Sovi, A.; Govoétchan, R.; Tokponnon, F.T.; Hounkonnou, H.; Aïkpon, R.; Agossa, F.; Gnanguenon, V.; Salako, A.S.; Agossou, C.D.; Ossè, R.; et al. Impact of land-use on malaria transmission in the plateau region, southeastern Benin. Parasites Vectors 2013, 6, 352. [Google Scholar] [CrossRef] [Green Version]
- Aïkpon, R.; Sèzonlin, M.; Tokponon, F.; Okè, M.; Oussou, O.; Oké-Agbo, F.; Beach, R.; Akogbéto, M. Good performances but short lasting efficacy of Actellic 50 EC Indoor Residual Spraying (IRS) on malaria transmission in Benin. West Africa. Parasites Vectors 2014, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Tokponnon, F.T.; Ogouyémi, A.H.; Sissinto, Y.; Sovi, A.; Gnanguenon, V.; Cornelie, S.; Adéothy, A.A.; Ossè, R.; Wakpo, A.; Gbenou, D.; et al. Impact of Long-Lasting, Insecticidal Nets on Anaemia and Prevalence of Plasmodium falciparum among Children under Five Years in Areas with Highly Resistant Malaria Vectors. Malar. J. 2014, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Djouaka, R.; Riveron, J.M.; Yessoufou, A.; Tchigossou, G.; Akoton, R.; Irving, H.; Djegbe, I.; Moutairou, K.; Adeoti, R.; Tamò, M.; et al. Multiple insecticide resistance in an infected population of the malaria vector Anopheles funestus in Benin. Parasites Vectors 2016, 9, 453. [Google Scholar] [CrossRef] [Green Version]
- PNLP. Plan National de Gestion de la Résistance des Vecteurs aux Insecticides 2018–2022; PNLP: Cotonou Benin, 2018. [Google Scholar]
- Hemingway, J.; Ranson, H.; Magill, A.; Kolaczinski, J.; Fornadel, C.; Gimnig, J.; Ano, A. Averting a malaria disaster: Will insecticide resistance derail malaria control? Lancet 2016, 387, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; Lissenden, N. Insecticide resistance in African Anopheles mosquitoes: A worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Ministère du Cadre de vie Bénin. Plan National de Sécheresse 2019–2024; Ministère du Cadre de vie Bénin: Cotonou, Benin, 2015. [Google Scholar]
- Gillies, M.T.; Coetzee, M. A Supplement to the Anophelinae of Africa South of the Sahara (Afrotropical Region). Publ. S. Afr. Inst. Med. Res. 1987, 1987, 55. [Google Scholar]
- Detinova, T.S.; Gillies, M.T. Observations on the determination of the Age composition and epidemiological importance of populations of Anopheles gambiae Giles and Anopheles funestus Giles in Tanganyika. Bull. World Health Organ. 1964, 30, 23–28. [Google Scholar]
- Wirtz, R.; Zavala, F.; Charoenvit, Y.; Campbell, G.H.; Burkot, T.R.; Schneider, I.; Esser, K.M.; Beaudoin, R.L.; Andre, R.G. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull. World Health Organ. 1987, 65, 39. [Google Scholar]
- Santolamazza, F.; Mancini, E.; Simard, F.; Qi, Y.; Tu, Z.; della Torre, A. Insertion polymorphisms of SINE200 retrotransposons within speciation islands of Anopheles gambiae molecular forms. Malar. J. 2008, 7, 163. [Google Scholar] [CrossRef] [Green Version]
- Koekemoer, L.L.; Kamau, L.; Hunt, R.H.; Coetzee, M. A cocktail polymerase chain reaction assay to identify members of the Anopheles funestus (Diptera: Culicidae) group. Am. J. Trop. Med. Hygen. 2002, 66, 804–811. [Google Scholar] [CrossRef]
- Kengne, P.; Awono, A.P.; Antonio-Nkondjio, C. Molecular identification of the Anopheles nili group African malaria vectors. Med. Vet. Entomol. 2003, 17, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Weill, M.; Malcolm, C.; Chandre, F.; Mogensen, K.; Berthomieu, A.; Marquine, M.; Raymond, M. The unique mutation in ace-1 giving high insecticide resistance is easily detectable in mosquito vectors. Insect Mol. Biol. 2004, 13, 1–7. [Google Scholar] [CrossRef]
- Beier, J.C.; Perkins, P.V.; Wirtz, R.A.; Koros, J.; Diggs, D.; Gargan, I.T.P.; Koech, D.K. Bloodmeal identification by direct enzyme-linked immunosorbent assay (ELISA) tested on Anopheles (Diptera: Culicidae) in Kenya. J. Med. Entomol. 1988, 25, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Robert, V. La Transmission, Composante Majeure de L’épidémiologie du Paludisme; Groupe de Recherche sur le Paludisme: Montpellier, France, 2004. [Google Scholar]
- Salako, A.S.; Ahogni, I.; Aïkpon, R.; Sidick, A.; Dagnon, F.; Sovi, A.; Sominahouin, A.A.; Agossa, F.; Iyikirenga, L.; Akogbeto, M.C. Insecticide resistance status, frequency of L1014F Kdr and G119S Ace-1 mutations, and expression of detoxification enzymes in Anopheles gambiae (s.l.) in two regions of northern Benin in preparation for indoor residual spraying. Parasites Vectors 2018, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Akogbeto, M.; Di Deco, M. Distribution of members of the Anopheles gambiae complex and their chromosomal variants in Benin and Togo, West Africa. Afr. Zool. 1995, 109, 443–454. [Google Scholar]
- Djouaka, R.; Irving, H.; Tukur, Z. Exploring mechanisms of multiple insecticide resistance in a population of the malaria vector Anopheles funestus in Benin. PLoS ONE 2011, 6, e27760. [Google Scholar] [CrossRef]
- Aïkpon, R.; Osse, R.; Govoetchan, R.; Sovi, A.; Oke-Agbo, F.; Akogbeto, M.C. Entomological baseline data on malaria transmission and susceptibility of Anopheles gambiae to insecticides in preparation for indoor residual spraying (IRS) in Atacora, (Benin). J. Parasitol. Vector Biol. 2013, 5, 102–111. [Google Scholar]
- Akogbéto, M.C.; Salako, A.S.; Dagnon, F.; Aïkpon, R.; Kouletio, M.; Sovi, A.; Sezonlin, M. Blood feeding behaviour comparison and contribution of Anopheles coluzzii and Anopheles gambiae, two sibling species living in sympatry, to malaria transmission in Alibori and Donga region, northern Benin, West Africa. Malar. J. 2018, 17, 307. [Google Scholar] [CrossRef]
- Akogbeto, M.; Romano, R. Infectivity of Anopheles melas vis-a-vis Plasmodium falciparum in the coastal lagoon area of Benin. Bull. Soc. Pathol. Exot. 1999, 92, 57–61. [Google Scholar]
- Yadouleton, A.; Asidi, A.; Djouaka, R.; Braïma, J.; Agossou, C.; Akogbeto, M. Development of vegetable farming: A cause of the emergence of insecticide resistance in populations of Anopheles gambiae in urban areas of Benin. Malar. J. 2009, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Yadouleton, A.; Martin, T.; Padonou, G.; Chandre, F.; Asidi, A.; Djogbenou, L.; Dabiré, R.; Aïkpon, R.; Boko, M.; Glitho, I.; et al. Cotton pest management practices and the selection of pyrethroid resistance in Anopheles gambiae population in Northern Benin. Parasite Vectors 2011, 4, 60. [Google Scholar] [CrossRef] [Green Version]
- Fontenille, D.; Cohuet, A.; Awono-Ambene, P.; Kengne, P.; Antonio-Nkondjio, C.; Wondji, C.; Simard, F. Vecteurs de paludisme: Du terrain à la génétique moléculaire. Rev. Epidémiologie St. Publique 2005, 53, 283–290. [Google Scholar] [CrossRef]
- Omumbo, J.A.; Guerra, C.A.; Hay, S.I.; Snow, R.W. The influence of urbanization on measures of Plasmodium falciparum infection prevalence in East Africa. Acta Trop. 2005, 93, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-J.; Lengeler, C.; Mtasiwa, D.; Mshana, T.; Manane, L.; Maro, G.; Tanner, M. Rapid urban malaria appraisal (RUMA) II: Epidemiology of urban malaria in Dar es Salaam (Tanzania). Malar. J. 2006, 5, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-J.; Lengeler, C.; Smith, T.; Vounatsou, P.; Diadie, D.; Pritroipa, X.; Convelbo, N.; Kientga, M.; Tanner, M. Rapid urban malaria appraisal (RUMA) I: Epidemiology of urban malaria in Ouagadougou. Malar. J. 2006, 4, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierrat, C. Des Moustiques et des Hommes: Les Territoires du Paludisme à Tori-Bossito (sud du Bénin). Thèse de Doctorat de Géographie, Université Paris I Panthéon-Sorbonne, Paris, France, 2010. [Google Scholar]
- Govoetchan, R.; Gnanguenon, V.; Azondékon, R.; Agossa, R.F.; Sovi, A.; Oké-Agbo, F.; Ossè, R.; Akogbéto, M. Evidence for perennial malaria in rural and urban areas under the Sudanian climate of Kandi, Northeastern Benin. Parasite Vectors 2014, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Terlouw, D.J.; Morgah, K.; Wolkon, A.; Dare, A.; Dorkenoo, A.; Eliades, M.J.; Eng, J.V.; Sodahlon, Y.K.; Ter Kuile, F.O.; Hawley, W.A. Impact of mass distribution of free long-lasting insecticidal nets on childhood malaria morbidity: The Togo national integrated child health campaign. Malar. J. 2010, 9, 199. [Google Scholar] [CrossRef] [Green Version]
- Stuckey, E.M.; Smith, T.A.; Chitnis, N. Estimating malaria transmission through mathematical models. Trends Parasitol. 2013, 29, 477–482. [Google Scholar] [CrossRef]
- Carneiro, I.; Roca-Feltrer, A.; Griffin, J.T.; Smith, L.; Tanner, M.; Schellenberg, J.A.; Greenwood, B.; Schellenberg, D. Age patterns of malaria vary with severity, transmission intensity and seasonality in sub-Saharan Africa: A systematic review and pooled analysis. PLoS ONE 2010, 5, e8988. [Google Scholar] [CrossRef] [Green Version]
- Stuckey, E.M.; Smith, T.; Chitnis, N. Seasonally Dependent Relationships between Indicators of Malaria Transmission and Disease Provided by Mathematical Model Simulations. PLoS Comput. Biol. 2014, 10, e1003812. [Google Scholar] [CrossRef] [Green Version]
- Garrett-Jones, C. Prognosis for interruption of malaria transmission through the assessment of mosquito vectorial capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef]
- Kelly-Hope, L.A.; McKenzie, F.E. The multiplicity of malaria transmission: A review of entomological inoculation rate measurements and methods across sub-Saharan Africa. Malar. J. 2009, 8, 19. [Google Scholar] [CrossRef]
- Gnanguenon, V.; Agossa, F.R.; Badirou, K.; Govoétchan, R.; Anagonou, R.; Oke-Agbo, F.; Azondekon, R.; AgbanrinYoussouf, R.; Attolou, R.; Tokponnon, F.T.; et al. Malaria vectors resistance to insecticides in Benin: Current trends and mechanisms involved. Parasite Vectors 2015, 8, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahouédo, G.A.; Cornelie, S.; Djègbè, I.; Ahlonsou, J.; Aboubakar, S.; Soares, C.; Akogbéto, M.; Corbel, V. Dynamics of pyrethroid resistance in malaria vectors in southern Benin following a large-scale implementation of vector control interventions. Parasite Vectors 2016, 9, 385. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Department | Communes (Latitude and Longitude) | Total Collected | Man Night | HBR/Commune | P1 | HBR/Department |
|---|---|---|---|---|---|---|
| Ouémé | Avrankou (6°33′32.07″ N 2°38′56.93″ E) | 14 | 32 | 0.438 | <0.001 | 1.125 a |
| Adjarra (6°31′22.69″ N 2°39′46.03″ E) | 58 | 32 | 1.813 | |||
| Plateau | Sakété (6°44′12.47″ N 2°39′49.01″ E) | 18 | 32 | 0.563 | 0.003 | 0.922 a |
| Adja Ouèrè (6°57′13.25″ N 2°34′44.25″ E) | 41 | 32 | 1.281 | |||
| Littoral | 3ème Arrond (6°22′12.76″ N 2°23′28.43″ E) | 964 | 32 | 30.125 | <0.001 | 34.594 b |
| 9ème Arrond (6°23′24.15″ N 2°24′03.44″ E) | 1250 | 32 | 39.063 | |||
| Collines | Savè (8°01′59.46″ N 2°29′08.17″ E) | 627 | 32 | 19.594 | 0.78 | 19.406 c |
| Ouèssè (8°29′35.07″ N 2°25′25.42″ E) | 615 | 32 | 19.219 | |||
| Atlantique | Zè (6°42′06.88″ N 2°20′31.84″ E) | 26 | 32 | 0.813 | <0.001 | 1.547 d |
| Toffo (6°50′24.08″ N 2°04′45.67″ E) | 73 | 32 | 2.281 | |||
| Zou | Zagnanado (7°16′47.86″ N 2°24′14.50″ E) | 149 | 32 | 4.656 | 0 | 9.219 e |
| Djidja (7°20′45.80″ N 1°56′08.19″ E) | 441 | 32 | 13.781 | |||
| Mono | Houéyogbé (6°31′52.28″ N 1°52′15.52″ E) | 273 | 32 | 8.531 | <0.001 | 6.906 f |
| Bopa (6°42′04.67″ N 1°57′01.73″ E) | 169 | 32 | 5.281 | |||
| Couffo | Toviklin (6°56′24.12″ N 1°46′23.30″ E) | 73 | 32 | 2.28 | 0.016 | 2.781 g |
| Djakotomè (6°53′45.68″ N 1°41′47.51″ E) | 105 | 32 | 3.281 | |||
| Atacora | Péhunco (10°30′02.35″ N 2°13′56.68″ E) | 128 | 32 | 4 | 0 | 15.094 h |
| Kérou (10°49′51.82″ N 2°06′38.25″ E) | 838 | 32 | 26.188 | |||
| Donga | Copargo (9°50′22.22″ N 1°32′51.32″ E) | 254 | 32 | 7.938 | 0.028 | 8.75 e |
| Ouaké (9°39′47.81″ N 1°23′12.90″ E) | 306 | 32 | 9.563 | |||
| Alibori | Karimama (12°04′08.91″ N 3°11′02.00″ E) | 684 | 32 | 21.375 | 0 | 11.078 h |
| Kandi (11°07′51.02″ N 2°56′05.75″ E) | 90 | 32 | 2.813 | |||
| Borgou | Nikki (9°55′51.14″ N 3°12′36.40″ E) | 10 | 32 | 0.313 | 0 | 1.562 d |
| Pèrèrè (9°47′31.29″ N 2°59′26.45″ E) | 25 | 32 | 0.781 | |||
| Total | 7231 | 768 | 9.415 |
| Departments | Communes | Indoor Total | Outdoor Total | HBR Indoor | HBR Outdoor | P1 |
|---|---|---|---|---|---|---|
| Ouémé | Avrankou | 9 | 5 | 0.563 | 0.313 | 0.285 |
| Adjarra | 32 | 26 | 2 | 1.625 | 0.431 | |
| Plateau | Sakété | 13 | 5 | 0.813 | 0.313 | 0.059 |
| Adja Ouèrè | 23 | 18 | 1.438 | 1.125 | 0.435 | |
| Littoral | 3ème Arrondissment | 469 | 495 | 29.313 | 30.938 | 0.402 |
| 9ème Arrondissment | 632 | 618 | 39.5 | 38.625 | 0.692 | |
| Collines | Savè | 256 | 371 | 16 | 23.188 | <0.001 |
| Ouèssè | 347 | 268 | 21.688 | 16.75 | 0.001 | |
| Atlantique | Zè | 20 | 6 | 1.25 | 0.375 | 0.006 |
| Toffo | 50 | 23 | 3.125 | 1.438 | 0.002 | |
| Zou | Zagnanado | 79 | 70 | 4.938 | 4.375 | 0.461 |
| Djidja | 247 | 194 | 15.438 | 12.125 | 0.012 | |
| Mono | Houéyogbé | 174 | 99 | 10.875 | 6.188 | <0.001 |
| Bopa | 114 | 55 | 7.125 | 3.438 | <0.001 | |
| Couffo | Toviklin | 48 | 25 | 3 | 1.563 | 0.007 |
| Djakotomè | 65 | 40 | 4.063 | 2.5 | 0.019 | |
| Atacora | Péhunco | 74 | 54 | 4.625 | 3.375 | 0.077 |
| Kérou | 430 | 408 | 26.875 | 25.5 | 0.447 | |
| Donga | Copargo | 122 | 132 | 7.625 | 8.25 | 0.53 |
| Ouaké | 159 | 147 | 9.938 | 9.188 | 0.687 | |
| Alibori | Karimama | 342 | 342 | 21.375 | 21.375 | 1 |
| Kandi | 23 | 67 | 1.438 | 4.188 | <0.001 | |
| Borgou | Nikki | 5 | 5 | 0.313 | 0.313 | 1 |
| Pèrèrè | 13 | 12 | 0.813 | 0.75 | 0.841 | |
| Total | 3746 | 3485 | 11.706 | 10.891 |
| Departement | Commune | Total Tested | N Positif | SI | HBR | Commune EIR | EIR 95%CI | EIR | EIR/Year/ |
|---|---|---|---|---|---|---|---|---|---|
| Commune | Department | Department | |||||||
| Ouémé | Avrankou | 14 | 0 | 0 | 0.44 | 0 | 0–0.008 | 0.016 a | 5.7 |
| Adjarra | 57 | 1 | 0.018 | 1.81 | 0.032 | 0.024–0.041 | |||
| Plateau | Sakété | 18 | 0 | 0 | 0.57 | 0 | 0–0.006 | 0.016 a | 5.63 |
| Adja Ouèrè | 41 | 1 | 0.024 | 1.29 | 0.031 | 0.022–0.042 | |||
| Littoral | 3ème Arrondissment | 250 | 3 | 0.012 | 30.13 | 0.362 | 0.348–0.375 | 0.992 b | 357.1 |
| 9ème Arrondissment | 308 | 13 | 0.042 | 39.1 | 1.649 | 1.623–1.674 | |||
| Collines | Savè | 87 | 9 | 0.103 | 19.6 | 2.027 | 1.974–2.081 | 1.663 c | 598.82 |
| Ouèssè | 193 | 15 | 0.078 | 19.2 | 1.494 | 1.463–1.524 | |||
| Atlantique | Zè | 25 | 2 | 0.08 | 0.81 | 0.065 | 0.049–0.085 | 0.063 d | 22.73 |
| Toffo | 73 | 2 | 0.027 | 2.28 | 0.062 | 0.053–0.073 | |||
| Zou | Zagnanado | 105 | 8 | 0.076 | 4.66 | 0.355 | 0.335–0.375 | 0.403 e | 145.08 |
| Djidja | 261 | 8 | 0.031 | 13.78 | 0.422 | 0.409–0.437 | |||
| Mono | Houéyogbé | 217 | 7 | 0.032 | 8.53 | 0.275 | 0.263–0.288 | 0.18 f | 64.67 |
| Bopa | 129 | 2 | 0.016 | 5.28 | 0.082 | 0.073–0.091 | |||
| Couffo | Toviklin | 73 | 5 | 0.068 | 2.28 | 0.156 | 0.141–0.173 | 0.153 g | 55.24 |
| Djakotomè | 72 | 3 | 0.042 | 3.28 | 0.137 | 0.122–0.153 | |||
| Atacora | Péhunco | 116 | 1 | 0.009 | 4 | 0.034 | 0.029–0.041 | 0.211 h | 76.1 |
| Kérou | 241 | 4 | 0.017 | 26.19 | 0.435 | 0.42–0.45 | |||
| Donga | Copargo | 60 | 2 | 0.033 | 7.94 | 0.265 | 0.242–0.289 | 0.39 e | 140.45 |
| Ouaké | 97 | 5 | 0.052 | 9.57 | 0.493 | 0.469–0.518 | |||
| Alibori | Karimama | 300 | 5 | 0.017 | 21.37 | 0.356 | 0.344–0.368 | 0.238 i | 85.63 |
| Kandi | 90 | 3 | 0.092 | 2.81 | 0.094 | 0.046–0.083 | |||
| Borgou | Nikki | 10 | 0 | 0 | 0.31 | 0 | −0.106 | 0.042 j | 15.07 |
| Pèrèrè | 25 | 2 | 0.077 | 0.78 | 0.062 | 0.082–0.012 | |||
| Total | 2875 | 101 | 0.0351 | 9.42 | 0.331 |
| Department | Commune | Total Collected | Blood Meal Origins | |||
|---|---|---|---|---|---|---|
| N Human (%) | N Cow (%) | N Sheep (%) | N Pig (%) | |||
| Ouémé | Avrankou | 4 | 3 (75) | 1 (25) | ||
| Adjarra | 6 | 5 (83.33) | 1 (16.67) | |||
| Plateau | Sakété | 1 | 1 (100) | |||
| Adja Ouèrè | 11 | 11 (100) | 1 (9.1) | |||
| Littoral | 3ème Arrondissment | 29 | 29 (100) | |||
| 9ème Arrondissment | 45 | 45 (100) | ||||
| Collines | Savè | 45 | 39 (86.67) | 3 (6.67) | 3 (6.67) | |
| Ouèssè | 20 | 20 (100) | ||||
| Atlantique | Zè | 3 | 3 (100) | |||
| Toffo | 1 | 1 (100) | ||||
| Zou | Zagnanado | 14 | 14 (100) | |||
| Djidja | 45 | 45 (100) | 3 (6.67) | |||
| Mono | Houéyogbé | 45 | 42 (93.33) | 3 (6.67) | 3 (6.67) | |
| Bopa | 15 | 14 (93.33) | 1 (6.67) | 1 (6.67) | ||
| Couffo | Toviklin | 20 | 20 (100) | |||
| Djakotomè | 15 | 15 (100) | 2 (13.33) | |||
| Atacora | Péhunco | 12 | 12 (100) | |||
| Kérou | 15 | 10 (66.67) | 1 (2.79) | 1 (6.67) | ||
| Donga | Copargo | 20 | 20 (100) | |||
| Ouaké | 15 | 12 (100) | ||||
| Alibori | Karimama | 75 | 60 (80) | 5 (6.67) | 5 (2.79) | |
| Kandi | 8 | 6 (75) | 1 (12.5) | 1 (12.5) | ||
| Borgou | Nikki | 0 | ||||
| Pèrèrè | 1 | 1 (100) | ||||
| Total | 465 | 428 (92.04) | 13 (2.79) | 15 (3.22) | 8 (1.72) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokponnon, T.F.; Ossè, R.; Padonou, G.G.; Affoukou, C.D.; Sidick, A.; Sewade, W.; Fassinou, A.; Koukpo, C.Z.; Akinro, B.; Messenger, L.A.; et al. Entomological Characteristics of Malaria Transmission across Benin: An Essential Element for Improved Deployment of Vector Control Interventions. Insects 2023, 14, 52. https://doi.org/10.3390/insects14010052
Tokponnon TF, Ossè R, Padonou GG, Affoukou CD, Sidick A, Sewade W, Fassinou A, Koukpo CZ, Akinro B, Messenger LA, et al. Entomological Characteristics of Malaria Transmission across Benin: An Essential Element for Improved Deployment of Vector Control Interventions. Insects. 2023; 14(1):52. https://doi.org/10.3390/insects14010052
Chicago/Turabian StyleTokponnon, Tatchémè Filémon, Razaki Ossè, Germain Gil Padonou, Cyriaque Dossou Affoukou, Aboubakar Sidick, Wilfried Sewade, Arsène Fassinou, Côme Z. Koukpo, Bruno Akinro, Louisa A. Messenger, and et al. 2023. "Entomological Characteristics of Malaria Transmission across Benin: An Essential Element for Improved Deployment of Vector Control Interventions" Insects 14, no. 1: 52. https://doi.org/10.3390/insects14010052
APA StyleTokponnon, T. F., Ossè, R., Padonou, G. G., Affoukou, C. D., Sidick, A., Sewade, W., Fassinou, A., Koukpo, C. Z., Akinro, B., Messenger, L. A., Okê, M., Tchévoédé, A., Ogouyemi-Hounto, A., Gazard, D. K., & Akogbeto, M. (2023). Entomological Characteristics of Malaria Transmission across Benin: An Essential Element for Improved Deployment of Vector Control Interventions. Insects, 14(1), 52. https://doi.org/10.3390/insects14010052