
Rotation of Multiple Single-Gene Transgenic Crops Did Not Slow the Evolution of Resistance to Cry1F or Cry1Ie in Ostrinia furnacalis



{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of O. furnacalis
2.2. Cry Toxins
2.3. Laboratory Selection Regimes
2.4. Susceptibility Bioassays
2.5. Statistical Analysis
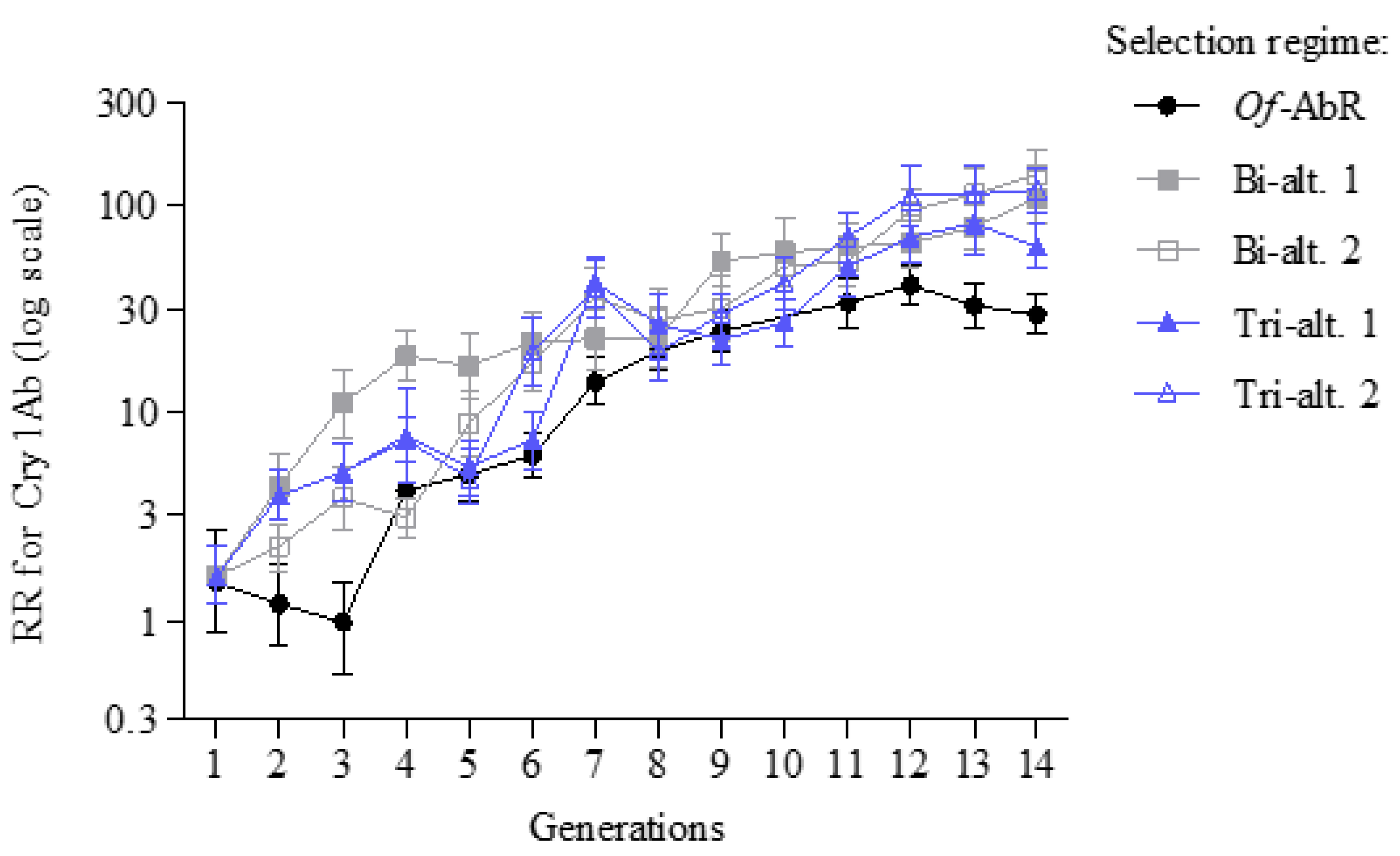
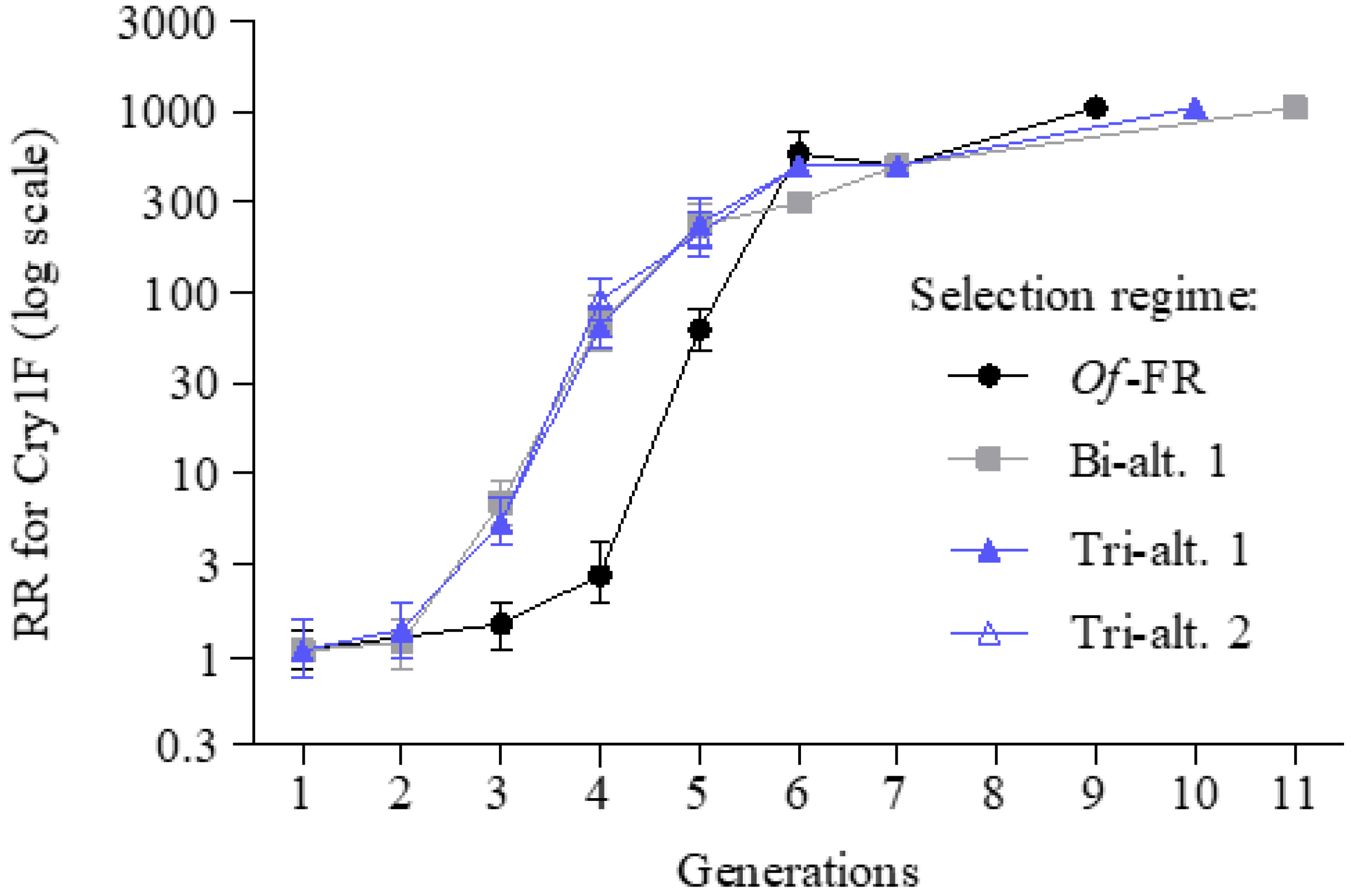
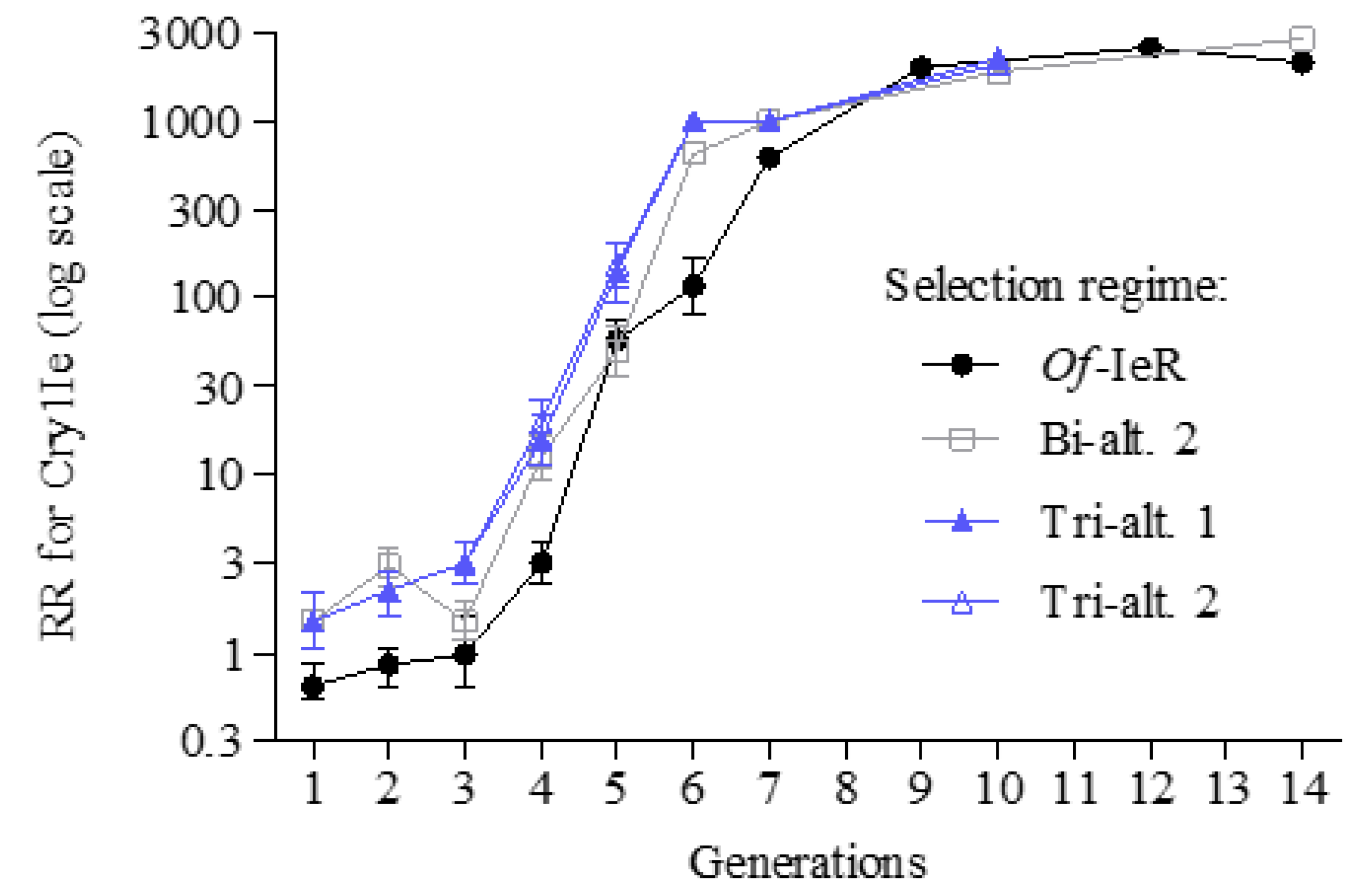
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roh, J.Y.; Choi, J.Y.; Li, M.S.; Jin, B.R.; Je, Y.H. Bacillus thuringiensis as a specific, safe, and effective tool for insect pest control. J. Microbiol. Biotechnol. 2007, 17, 547–559. [Google Scholar] [PubMed]
- Pardo-Lopez, L.; Soberon, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISAAA. Breaking Barriers with Breeding: A Primer on New Breeding Innovations for Food Security; ISAAA Brief No. 56; ISAAA: Ithaca, NY, USA, 2021. [Google Scholar]
- Carrière, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Natl. Acad. Sci. USA 2003, 100, 1519–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Guo, Y.Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.M.; Randy, M.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Smith, J.L.; Farhan, Y.; Schaafsma, A.W. Practical resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) to Cry1F Bacillus thuringiensis maize discovered in nova Scotia, Canada. Sci. Rep. 2019, 9, 18247. [Google Scholar] [CrossRef] [Green Version]
- Roush, R.T. Bt-transgenic crops: Just another pretty insecticide or a chance for a new start in resistance management? Pestic. Sci. 1997, 51, 328–334. [Google Scholar] [CrossRef]
- Carrière, Y.; Tabashnik, B.E. Reversing insect adaptation to transgenic insecticidal plants. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.Z.; Cao, J.; Li, Y.; Collins, H.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat. Biotechnol. 2003, 21, 1493–1497. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carriere, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef]
- Bourguet, D.; Delmotte, F.; Franck, P.; Guillemaud, T.; Reboud, X.; Vacher, C.; Walker, A.S. Heterogeneity of selection and the evolution of resistance. Trends Ecol. Evol. 2013, 28, 110–118. [Google Scholar]
- Ni, M.; Ma, W.; Wang, X.; Gao, M.; Dai, Y.; Wei, X.; Zhang, L.; Peng, Y.; Chen, S.; Ding, L.; et al. Next-generation transgenic cotton: Pyramiding RNAi and Bt counters insect resistance. Plant Biotechnol. J. 2017, 15, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Q.; Quan, Y.D.; Yang, J.; Shu, C.L.; Wang, Z.Y.; Zhang, J.; Gatehouse, A.M.R.; Tabashnik, B.E.; He, K.L. Evolution of Asian corn borer resistance to Bt toxins used singly or in pairs. Toxins 2019, 11, 461. [Google Scholar] [CrossRef] [Green Version]
- Roush, R.T. Designing resistance management programs: How can you choose? Pestic. Sci. 1989, 26, 423–441. [Google Scholar] [CrossRef]
- Zhao, J.Z.; Collins, H.L.; Shelton, A.M. Testing insecticide resistance management strategies: Mosaic versus rotations. Pest Manag. Sci. 2010, 66, 1101–1105. [Google Scholar] [CrossRef]
- Afidchao, M.M.; Musters, C.J.; Snoo, G.R. Asian corn borer (ACB) and non-ACB pests in GM corn (Zea mays L.) in the Philippines. Pest Manag. Sci. 2013, 69, 792–801. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Liu, Y.J.; Ren, Y.; Liu, Y.; Liang, G.M.; Song, F.P.; Bai, S.X.; Wang, J.H.; Wang, G.Y. Overexpression of a novel cry1Ie gene confers resistance to Cry1Ac-resistant cotton bollworm in transgenic lines of maize. Plant Cell Tissue Organ Cult. 2013, 115, 151–158. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Baxter, S.W.; Zhao, J.Z.; Shelton, A.M.; Gould, F.; Tabashnik, B.E. The diversity of Bt resistance genes in species of Lepidoptera. J. Invertebr. Pathol. 2007, 95, 192–197. [Google Scholar] [CrossRef]
- Ferré, J.; Van Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef]
- Zhao, C.; Jurat-Fuentes, J.L.; Abdelgaffar, H.M.; Pan, H.; Song, F.; Zhang, J. Identification of a new cry1I-type gene as a candidate for gene pyramiding in corn to control Ostrinia species larvae. Appl. Environ. Microb. 2015, 81, 3699–3705. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Yang, J.; Quan, Y.D.; Wang, Z.Y.; Cai, W.Z.; He, K.L. Characterization of Asian corn borer resistance to Bt toxin Cry1Ie. Toxins 2017, 9, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wang, Z.; Zhang, J.; He, K.; Ferry, N.; Gatehouse, A.M.R. Cross-resistance of Cry1Ab-selected Asian corn borer to other Cry toxins. J. Appl. Entomol. 2010, 134, 429–438. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Wang, Y.D.; Wang, Z.Y.; Bravo, A.; Soberon, M.; He, K.L. Genetic basis of Cry1F-resistance in a laboratory select-ed Asian corn borer strain and its cross-resistance to other Bacillus thuringiensis toxins. PLoS ONE 2016, 11, e0161189. [Google Scholar]
- Wang, X.; Xu, Y.; Huang, J.; Jin, W.; Yang, Y.; Wu, Y. CRISPR-mediated knockout of the ABCC2 gene in Ostrinia furnacalis confers high-level resistance to the Bacillus thuringiensis Cry1Fa toxin. Toxins 2020, 12, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.Y.; Zhou, D.R.; He, K.L. Studies on mass rearing of Asian corn borer: Development of a satisfactory non-agar semi-artificial diet and its use. Acta Phytophylacica Sin 1999, 26, 324–328. [Google Scholar]
- Guo, S.; Zhang, C.; Lin, X.; Zhang, Y.; He, K.; Song, F.; Zhang, J. Purification of an active fragment of Cry1Ie toxin from Bacillus thuringiensis. Protein Expr. Purif. 2011, 78, 204–208. [Google Scholar] [CrossRef]
- Roush, R.T. Two-toxin strategies for management of insect resistant transgenic crops: Can pyramiding succeed where pesticide mixtures have not? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1998, 353, 1777–1786. [Google Scholar] [CrossRef] [Green Version]
- Gould, F. Bt-resistance management—Theory meets data. Nat. Biotechnol. 2003, 21, 1450–1451. [Google Scholar] [CrossRef]
- Zhang, T.T.; He, M.X.; Gatehouse, A.; Wang, Z.Y.; Edwards, M.; Li, Q.; He, K.L. Inheritance patterns, dominance and cross-resistance of Cry1Ab-and Cry1Ac-selected Ostrinia furnacalis (Guenée). Toxins 2014, 6, 2694–2707. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.J.G.; Storer, N.P.; Siegfried, B.D. Inheritance of Cry1F resistance in laboratory-selected European corn borer and its survival on transgenic corn expressing the Cry1F toxin. Bull. Entomol. Res. 2008, 98, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Coates, B.S.; Sumerford, D.V.; Siegfried, B.D.; Hellmich, R.L.; Abel, C.A. Unlinked genetic loci control the reduced transcription of aminopeptidase N 1 and 3 in the European corn borer and determine tolerance to Bacillus thuringiensis Cry1Ab toxin. Insect Biochem. Mol. Biol. 2013, 43, 1152–1160. [Google Scholar] [CrossRef] [Green Version]
- Janmaat, A.F.; Myers, J. Rapid evolution and the cost of resistance to Bacillus thuringiensis in greenhouse populations of cabbage loopers, Trichoplusia ni. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 2263–2270. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Gould, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics and ecology. Annu. Rev. Entomol. 1998, 43, 701–726. [Google Scholar] [CrossRef] [Green Version]
- McGaughey, W.H.; Whalon, M.E. Managing insect resistance to Bacillus thuringiensis toxins. Science 1992, 258, 1451–1455. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Quan, Y.; Wang, Z.; He, K. Rotation of Multiple Single-Gene Transgenic Crops Did Not Slow the Evolution of Resistance to Cry1F or Cry1Ie in Ostrinia furnacalis. Insects 2023, 14, 74. https://doi.org/10.3390/insects14010074
Wang Y, Quan Y, Wang Z, He K. Rotation of Multiple Single-Gene Transgenic Crops Did Not Slow the Evolution of Resistance to Cry1F or Cry1Ie in Ostrinia furnacalis. Insects. 2023; 14(1):74. https://doi.org/10.3390/insects14010074
Chicago/Turabian StyleWang, Yueqin, Yudong Quan, Zhenying Wang, and Kanglai He. 2023. "Rotation of Multiple Single-Gene Transgenic Crops Did Not Slow the Evolution of Resistance to Cry1F or Cry1Ie in Ostrinia furnacalis" Insects 14, no. 1: 74. https://doi.org/10.3390/insects14010074
APA StyleWang, Y., Quan, Y., Wang, Z., & He, K. (2023). Rotation of Multiple Single-Gene Transgenic Crops Did Not Slow the Evolution of Resistance to Cry1F or Cry1Ie in Ostrinia furnacalis. Insects, 14(1), 74. https://doi.org/10.3390/insects14010074