

The Roles of Mating, Age, and Diet in Starvation Resistance in Bactrocera oleae (Olive Fruit Fly)
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
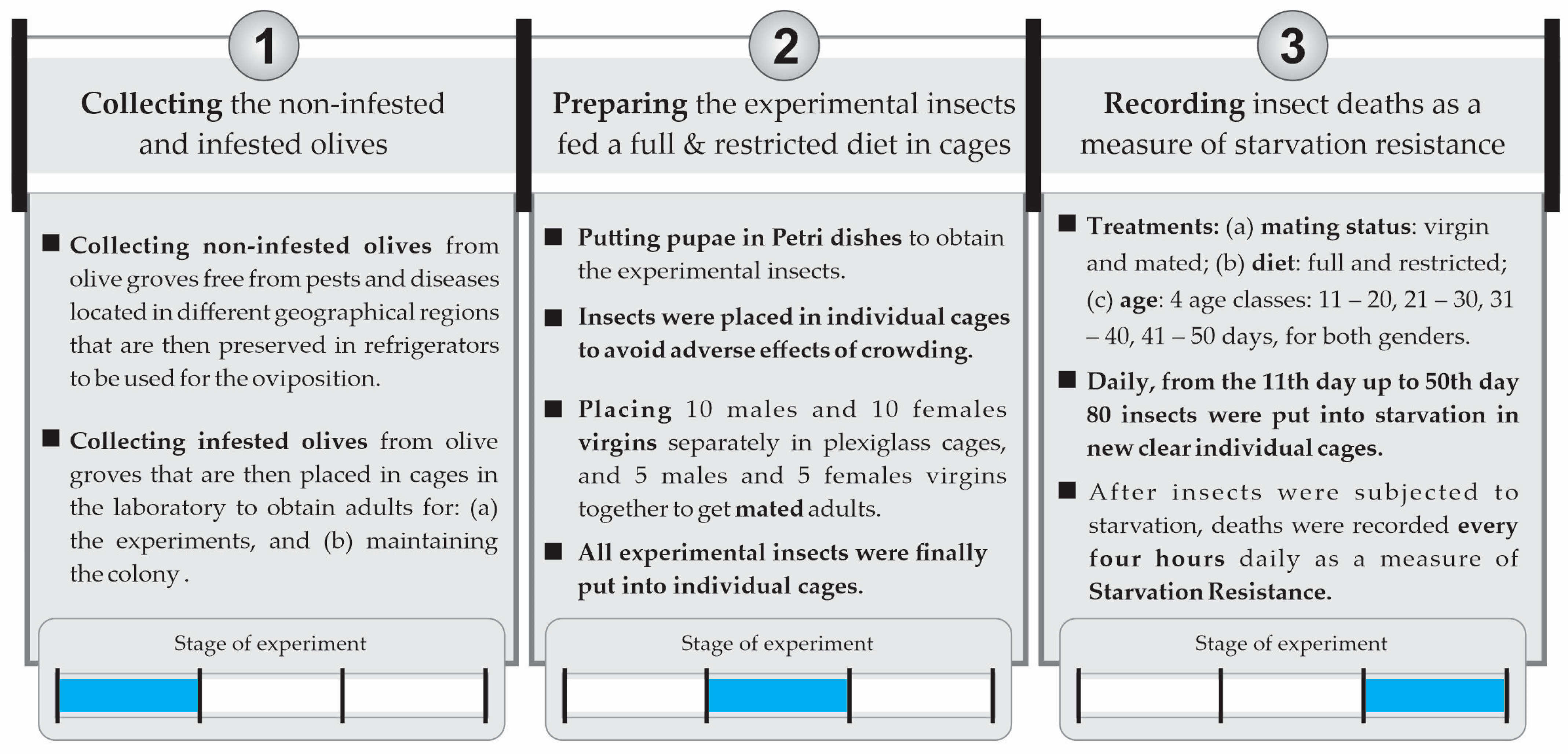
2. Materials and Methods
2.1. Origin and Handling of Experimental Insects and Olive Fruit
- (a)
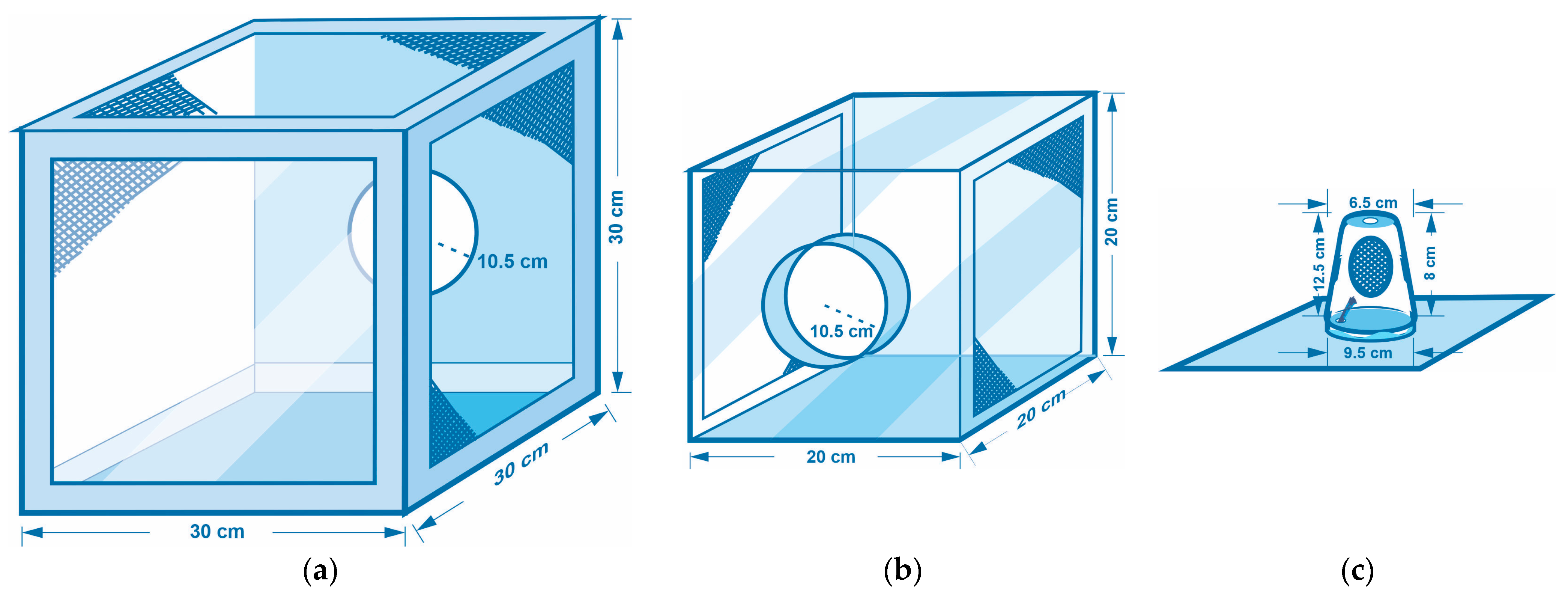
- Insect cage types used in the experiments: The wild adult olive fly lines were housed in custom-made insect cages based on the model of BugDorm cages (Model DP1000B) usually used in entomological experiments [44]. The types of insect cages used in the laboratory for the needs of the experiments were (A) BugDorm-type cages, 30 × 30 × 30 cm3 (colony cages for rearing of the olive fruit fly) (Figure 2a); (B) transparent plexiglass cages with dimensions 20 × 20 × 20 cm3 (Figure 2b), where (i) pupae gathered from infested olives in basins were placed into Petri dishes and then were transferred into the plexiglass cages for the adults to emerge (see experimental procedures and protocol for further information), and (ii) flies on the 10th day of their lives were transferred to mate or to be together with other flies of the same sex; (C) individual plastic cages (Figure 2c), where (i) flies were transferred individually upon their emergence, with either full or restricted diet and water, and (ii) flies were transferred at the appropriate age, each time to a new individual plastic cage that was thoroughly clean of any trace of food to measure the hours until death (starvation resistance).
- (b)
- Diet food types for adult fruit flies: Two different diets were used: (A) a full diet consisting of a mixture of hydrolyzed yeast (protein) in a ratio of 5:4:1 (water/sugar/yeast hydrolysate as protein) or (B) a restricted diet containing only sugar but deprived of protein. Water was supplied to all cohorts through a wetted cotton wick.
- (c)
- Harvesting (non-infested) olive fruit for the rearing of olive fruit flies: Olive fruits used in the experiments were collected from olive groves located in the region of Chalkidiki and Northern Greece. The olives were selected one-by-one by hand in the above regions from trees that were as free from pests and diseases as possible. To maintain the number of insects needed for the experiment and the genetic diversity of the experimental insect population close to that of natural populations, infested olives were constantly collected from olive groves for a period of approximately three months, and wild insects were constantly introduced to the colony. The total amount of olives needed for the experiment was roughly 200 kg (Figure 3).
- (d)
- Maintaining the (non-infested) olive fruit flies: Immediately after harvesting, the olive fruits were placed in glass jars in the refrigerator at 6 ± 1 °C (Figure 3).
- (e)
- Collecting infested olive fruits: Mc Cain traps with an appropriate food attractant were used in the aforementioned regions to identify the period of the first adult flights and the onset of infestation in the field. Olive fruits that had been infested by the olive flies were collected from the trees and transported to the laboratory (Figure 3).
2.2. Experimental Design
2.3. Experimental Procedures and Protocol
- (a)
- Rearing the parents of the experimental insects: Adults from infested olive fruits that had been collected from trees hatched inside wooden cages with plenty of water and protein food. After the completion of their hatching, the olives of their origin were removed, and fresh olives were added (ones that we had collected and maintained in the refrigerator). After mating, the females laid their eggs in the olive fruit. These infested olives were removed from the cages, laid into basins, and covered with a suitable cloth to ensure the appropriate humidity and temperature conditions (Figure 3). After pupation and before the emergence of the adults, pupae were transferred to plexiglass cages with dimensions 20 × 20 cm, awaiting the appearance of the adults (Figure 2b). In these plexiglass cages, there was either a full or restricted diet and water (Figure 4).
- (b)
- Handling the experimental insects before the experiment: Upon emergence, adults were placed in individual plastic cages (Figure 2c) with water and food (either the full or restricted diet). At the age of 10 days, groups of 10 adults of either only females and males (both virgin) or 5 virgin males and 5 virgin females (mated) were allowed to be together in larger cages (20 × 20 cm) for one day (Figure 2b) before being placed back into individual cages (Figure 2c). After this period, the flies were placed back into individual cages (Figure 2c) to eliminate crowding and social interactions (Figure 4). Flies that had been kept with conspecifics of the opposite sex were monitored by a human observer to verify mating. We observed the flies for mating from 16:00 to 21:00 because, in this species, mate searching and courtship take place during the late evening [34,36]. Individuals that had not mated were removed from the experiment and were replaced with others that had mated.
- (c)
- Preparing the flies to undergo starvation (food deprivation): The steps followed (Figure 4): (1) initially, experimental insects (pupae) were placed in Petri dishes in plexiglass cages; (2) upon adult emergence, they were transferred individually to plastic cages with water, half with the full diet and half with the restricted diet; (3) on the 10th day, all insects were transferred to 8 plexiglass cages: (a) 4 cages with the full diet (40 adults in total: 1 cage with 10 males, 1 with 10 females, and 2 cages with 5 males and 5 females in each cage) and (b) 4 cages with the restricted diet (40 adults in total: 1 cage with 10 males, 1 with 10 females, and 2 cages with 5 males and 5 females in each cage); (4) at the end of the 10th day, insects were transferred back to their individual cages with water and with the same diet that they were fed in the plexiglass cage.
- (d)
- Recording of deaths—Calculating starvation resistance: Upon reaching the eleventh day of their adult life, ten individual adults from each treatment at a specific age (11th, 12th, up to 50th day of life) were each transferred to a new individual plastic cage (Figure 2c) thoroughly clean of any trace of food [16]. The insects’ deaths were recorded every four hours due to food deprivation during the light period (four times per day: 08:00, 12:00, 16:00, 20:00). In Figure 5, a schematic representation showing the feeding and starvation stages is given. From the 11th day up to the 50th day of their lifespan, 80 insects (3200 in total) fed the full or restricted diet (40 adults fed the full diet and 40 adults the restricted diet, in each case: 10 virgin males, 10 virgin females, 10 mated males, and 10 mated females) were subjected to starvation in new clear individual cages. Within the period from the 11th to the 50th day, every four hours, the deaths were recorded as a measure of starvation resistance. In case there was difficulty in identifying an insect’s death, a fine paintbrush was used to gently move the insect and confirm its death. Rotation of the plastic cages was performed daily to reduce potential experimental errors. Starvation resistance was finally calculated as the difference between the date and time of death and the date and time of the moment the insects were subjected to food deprivation.
2.4. Statistical Analyses
3. Results
3.1. Study of Starvation Resistance in Males
3.2. Study of Starvation Resistance in Females
3.3. Percentage Starvation Resistance Differences from the Corresponding Gender Mean
4. Discussion
5. Conclusions
- Mating status: virgin adults exhibit higher starvation resistance compared to mated adults in all age classes.
- Age: younger adults exhibit higher starvation resistance in almost all treatments.
- Diet: adults that are fed a full diet containing protein show notably lower starvation resistance compared to sugar-fed ones.
- Gender: the same pattern of starvation resistance has been identified for both genders.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bengochea, P.; Christiaens, O.; Amor, F.; Viñuela, E.; Rougé, P.; Medina, P.; Smagghe, G. Insect Growth Regulators as Potential Insecticides to Control Olive Fruit Fly (Bactrocera oleae Rossi): Insect Toxicity Bioassays and Molecular Docking Approach. Pest. Manag. Sci. 2013, 69, 27–34. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Marketaki, M.; Charalampous, A.; Anagnostopoulos, C.; Bempelou, E. Residual Degradation and Toxicity of Insecticides against Bactrocera oleae. Environ. Sci. Pollut. Res. 2018, 25, 479–489. [Google Scholar] [CrossRef]
- Nardi, F.; Carapelli, A.; Dallai, R.; Roderick, G.K.; Frati, F. Population Structure and Colonization History of the Olive Fly, Bactrocera oleae (Diptera, Tephritidae). Mol. Ecol. 2005, 14, 2729–2738. [Google Scholar] [CrossRef]
- Malheiro, R.; Casal, S.; Baptista, P.; Pereira, J.A. A Review of Bactrocera oleae (Rossi) Impact in Olive Products: From the Tree to the Table. Trends Food Sci. Technol. 2015, 44, 226–242. [Google Scholar] [CrossRef]
- Berg, H.; Maneas, G.; Engström, A.S. A Comparison between Organic and Conventional Olive Farming in Messenia, Greece. Horticulturae 2018, 4, 15. [Google Scholar] [CrossRef]
- Sánchez, A.G.; Martos, N.R.; Ballesteros, E. Multiresidue Analysis of Pesticides in Olive Oil by Gel Permeation Chromatography Followed by Gas Chromatography-Tandem Mass-Spectrometric Determination. Anal. Chim. Acta 2006, 558, 53–61. [Google Scholar] [CrossRef]
- Raina, B. Olives; Elsevier Science Ltd.: Jammu, India, 2003. [Google Scholar]
- Pinheiro, L.A.; Dáder, B.; Wanumen, A.C.; Pereira, J.A.; Santos, S.A.P.; Medina, P. Side Effects of Pesticides on the Olive Fruit Fly Parasitoid Psyttalia Concolor (Szépligeti): A Review. Agronomy 2020, 10, 1755. [Google Scholar] [CrossRef]
- Bueno, A.M.; Jones, O. Alternative Methods for Controlling the Olive Fly, Bactrocera oleae, Involving Semiochemicals. In Use of Pheromones and Other Semiochemicals in Integrated Production; IOBC Wprs Bulletin: Andalusia, Spain, 2002; Volume 25. [Google Scholar]
- Ant, T.; Koukidou, M.; Rempoulakis, P.; Gong, H.F.; Economopoulos, A.; Vontas, J.; Alphey, L. Control of the Olive Fruit Fly Using Genetics-Enhanced Sterile Insect Technique. BMC Biol. 2012, 10, 51. [Google Scholar] [CrossRef]
- Ahmad, S.; Haq, I.U.; Cáceres, C.; Tomas, U.S.; Dammalage, T.; Gembinsky, K.; Paulus, H.; Vreysen, M.J.B.; Rempoulakis, P. One for All: Mating Compatibility among Various Populations of Olive Fruit Fly (Diptera: Tephritidae) for Application of the Sterile Insect Technique. PLoS ONE 2018, 13, e0206739. [Google Scholar] [CrossRef]
- Choi, K.S.; Samayoa, A.C.; Hwang, S.Y.; Huang, Y.B.; Ahn, J.J. Thermal Effect on the Fecundity and Longevity of Bactrocera dorsalis Adults and Their Improved Oviposition Model. PLoS ONE 2020, 15, e0235910. [Google Scholar] [CrossRef]
- Fanson, B.G.; Taylor, P.W. Additive and Interactive Effects of Nutrient Classes on Longevity, Reproduction, and Diet Consumption in the Queensland Fruit Fly (Bactrocera tryoni). J. Insect Physiol. 2012, 58, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Powell, K.S.; Weldon, C.W.; Taylor, P.W. The Ecology of Bactrocera tryoni (Diptera: Tephritidae): What Do We Know to Assist Pest Management? Ann. Appl. Biol. 2011, 158, 26–54. [Google Scholar] [CrossRef]
- Aita, R.C.; Kees, A.M.; Aukema, B.H.; Hutchison, W.D.; Koch, R.L. Effects of Starvation, Age, and Mating Status on Flight Capacity of Laboratory-Reared Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). Environ. Entomol. 2021, 50, 532–540. [Google Scholar] [CrossRef]
- Gerofotis, C.D.; Kouloussis, N.A.; Koukougiannidou, C.; Papadopoulos, N.T.; Damos, P.; Koveos, D.S.; Carey, J.R. Age, Sex, Adult and Larval Diet Shape Starvation Resistance in the Mediterranean Fruit Fly: An Ecological and Gerontological Perspective. Sci. Rep. 2019, 9, 10704. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.Y.A.; Hoy, M.A. Genetic Improvement of a Parasitoid Biological Control Agent: Artificial Selection for Insecticide Resistance in Aphytis Melinus (Hymenoptera: Aphelinidae). J. Econ. Entomol. 1988, 81, 1539–1550. [Google Scholar]
- Harwood, J.F.; Chen, K.; Müller, H.G.; Wang, J.L.; Vargas, R.I.; Carey, J.R. Effects of Diet and Host Access on Fecundity and Lifespan in Two Fruit Fly Species with Different Life-History Patterns. Physiol. Entomol. 2013, 38, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Perez-Mendoza, J.; Campbell, J.F.; Throne, J.E. Influence of Age, Mating Status, Sex, Quantity of Food, and Long-Term Food Deprivation on Red Flour Beetle (Coleoptera: Tenebrionidae) Flight Initiation. J. Econ. Entomol. 2011, 104, 2078–2086. [Google Scholar] [CrossRef]
- Zhang, D.W.; Xiao, Z.J.; Zeng, B.P.; Li, K.; Tang, Y.L. Insect Behavior and Physiological Adaptation Mechanisms under Starvation Stress. Front. Physiol. 2019, 10, 163. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D.; Messing, R.H.; Canale, A. Population-Level Lateralized Aggressive and Courtship Displays Make Better Fighters Not Lovers: Evidence from a Fly. Behav. Process. 2015, 115, 163–168. [Google Scholar] [CrossRef]
- Kouloussis, N.A.; Damos, P.T.; Ioannou, C.S.; Tsitsoulas, C.; Papadopoulos, N.T.; Nestel, D.; Koveos, D.S. Age Related Assessment of Sugar and Protein Intake of Ceratitis capitata in Ad Libitum Conditions and Modeling Its Relation to Reproduction. Front. Physiol. 2017, 8, 271. [Google Scholar] [CrossRef]
- Adnan, S.M.; Mendez, V.; Morelli, R.; Akter, H.; Farhana, I.; Taylor, P.W. Dietary Methoprene Supplement Promotes Early Sexual Maturation of Male Queensland Fruit Fly Bactrocera tryoni. J. Pest. Sci. 2018, 91, 1441–1454. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Carey, J.R.; Papadopoulos, N.T. Effects of Early-Life Protein Starvation on Longevity and Sexual Performance of Male Medfly. PLoS ONE 2019, 14, e0219518. [Google Scholar] [CrossRef]
- Goenaga, J.; Mensch, J.; Fanara, J.J.; Hasson, E. The Effect of Mating on Starvation Resistance in Natural Populations of Drosophila melanogaster. Evol. Ecol. 2012, 26, 813–823. [Google Scholar] [CrossRef]
- Service, P.M. The Effect of Mating Status on Lifespan, Egg Laying, and Starvation Resistance in Drosophila melanogaster in Relation to Selection on Longevity. J. Insect Physiol. 1989, 35, 447–452. [Google Scholar] [CrossRef]
- Hicks, S.K.; Hagenbuch, K.L.; Meffert, L.M. Variable Costs of Mating, Longevity, and Starvation Resistance in Musca Domestica (Diptera: Muscidae). Environ. Entomol. 2004, 33, 779–786. [Google Scholar] [CrossRef]
- Tasnin, M.S.; Merkel, K.; Clarke, A.R. Effects of Advanced Age on Olfactory Response of Male and Female Queensland Fruit Fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). J. Insect Physiol. 2020, 122, 104024. [Google Scholar] [CrossRef]
- Llandres, A.L.; Marques, G.M.; Maino, J.L.; Kooijman, S.A.L.M.; Kearney, M.R.; Casas, J. A Dynamic Energy Budget for the Whole Life-Cycle of Holometabolous Insects. Ecol. Monogr. 2015, 85, 353–371. [Google Scholar] [CrossRef]
- Edmunds, D.; Wigby, S.; Perry, J.C. ‘Hangry’ Drosophila: Food Deprivation Increases Male Aggression. Anim. Behav. 2021, 177, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Knoppien, P.; Van Der Pers, J.N.C.; Van Delden, W. Quantification of Locomotion and the Effect of Food Deprivation on Locomotor Activity in Drosophila. J. Insect Behav. 2000, 13, 27–43. [Google Scholar] [CrossRef]
- Krittika, S.; Yadav, P. Trans-Generational Effect of Protein Restricted Diet on Adult Body and Wing Size of Drosophila melanogaster. R. Soc. Open Sci. 2022, 9, 211325. [Google Scholar] [CrossRef]
- Flatt, T. Survival Costs of Reproduction in Drosophila. Exp. Gerontol. 2011, 46, 369–375. [Google Scholar] [CrossRef]
- Kokkari, A.I.; Milonas, P.G.; Anastasaki, E.; Floros, G.D.; Kouloussis, N.A.; Koveos, D.S. Determination of Volatile Substances in Olives and Their Effect on Reproduction of the Olive Fruit Fly. J. Appl. Entomol. 2021, 145, 841–855. [Google Scholar] [CrossRef]
- Terzidou, A.; Koveos, D.; Kouloussis, N. Mating Competition between Wild and Artificially Reared Olive Fruit Flies. Crops 2022, 2, 247–257. [Google Scholar] [CrossRef]
- Terzidou, A.M.; Koveos, D.S.; Papadopoulos, N.T.; Carey, J.R.; Kouloussis, N.A. Artificial Diet Alters Activity and Rest Patterns in the Olive Fruit Fly. PLoS ONE 2023, 18, e0274586. [Google Scholar] [CrossRef]
- Jacome, I.; Aluja, M.; Liedo, P.; Nestel, D. The Influence of Adult Diet and Age on Lipid Reserves in the Tropical Fruit Fly Anastrepha serpentina (Diptera: Tephritidae). J. Insect Physiol. 1995, 41, 1079–1086. [Google Scholar] [CrossRef]
- Utgés, M.E.; Vilardi, J.C.; Oropeza, A.; Toledo, J.; Liedo, P. Pre-Release Diet Effect on Field Survival and Dispersal of Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae). J. Appl. Entomol. 2013, 137, 163–177. [Google Scholar] [CrossRef]
- Yuval, B.; Maor, M.; Levy, K.; Kaspi, R.; Taylor, P.; Shelly, T. Breakfast of Champions or Kiss of Death? Survival and Sexual Performance of Protein-Fed, Sterile Mediterranean Fruit Flies (Diptera: Tephritidae). Fla. Entomol. 2007, 90, 115–122. [Google Scholar] [CrossRef]
- Shelly, T.E.; Kennelly, S.S. Starvation and the Mating Success of Wild Male Mediterranean Fruit Flies (Diptera: Tephritidae). J. Insect Behav. 2003, 16, 171–179. [Google Scholar] [CrossRef]
- McCue, M.D. Starvation Physiology: Reviewing the Different Strategies Animals Use to Survive a Common Challenge. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 1–18. [Google Scholar] [PubMed]
- Jose Gonzalez Minero, F.; Candau, P.; Morales, J.; Tomas, C. Forecasting Olive Crop Production Based on Ten Consecutive Years of Monitoring Airborne Pollen in Andalusia (Southern Spain). Agric. Ecosyst. Environ. 1998, 69, 201–215. [Google Scholar] [CrossRef]
- Rossini, L.; Bruzzone, O.A.; Contarini, M.; Bufacchi, L.; Speranza, S. A Physiologically Based ODE Model for an Old Pest: Modeling Life Cycle and Population Dynamics of Bactrocera oleae (Rossi). Agronomy 2022, 12, 2298. [Google Scholar] [CrossRef]
- Marubbi, T.; Cassidy, C.; Miller, E.; Koukidou, M.; Martin-Rendon, E.; Warner, S.; Loni, A.; Beech, C. Exposure to Genetically Engineered Olive Fly (Bactrocera oleae) Has No Negative Impact on Three Non-Target Organisms. Sci. Rep. 2017, 7, 11478. [Google Scholar] [CrossRef]
- Austad, S.N.; Fischer, K.E. Sex Differences in Lifespan. Cell Metab. 2016, 23, 1022–1033. [Google Scholar] [PubMed]
- Toothaker, L.E. Multiple Comparison Procedures; Sage Publications, Inc.: Thousand Oaks, CA, USA, 1993. [Google Scholar]
- Field, A. Discovering Statistics Using IBM SPSS Statistics: And Sex and Drugs and Rock “N” Roll, 4th ed.; Sage: Los Angeles, CA, USA; London, UK; New Delhi, India, 2013. [Google Scholar]
- Rush, B.; Sandver, S.; Bruer, J.; Roche, R.; Wells, M.; Giebultowicz, J. Mating Increases Starvation Resistance and Decreases Oxidative Stress Resistance in Drosophila melanogaster Females. Aging Cell 2007, 6, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake, E.W.M.T.D.; Clarke, A.R.; Schutze, M.K. Effect of Body Size, Age, and Premating Experience on Male Mating Success in Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Entomol. 2017, 110, 2278–2281. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.T.; Liedo, P.; Müller, H.G.; Wang, J.L.; Molleman, F.; Carey, J.R. Cost of Reproduction in Male Medflies: The Primacy of Sexual Courting in Extreme Longevity Reduction. J. Insect Physiol. 2010, 56, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Terzidou, A.; Kouloussis, N.; Papanikolaou, G.; Koveos, D. Acoustic Characteristics of Sound Produced by Males of Bactrocera oleae Change in the Presence of Conspecifics. Sci. Rep. 2022, 12, 13086. [Google Scholar] [CrossRef]
- Benelli, G.; Canale, A.; Bonsignori, G.; Ragni, G.; Stefanini, C.; Raspi, A. Male Wing Vibration in the Mating Behavior of the Olive Fruit Fly Bactrocera oleae (Rossi) (Diptera: Tephritidae). J. Insect Behav. 2012, 25, 590–603. [Google Scholar] [CrossRef]
- Benelli, G.; Daane, K.M.; Canale, A.; Niu, C.Y.; Messing, R.H.; Vargas, R.I. Sexual Communication and Related Behaviours in Tephritidae: Current Knowledge and Potential Applications for Integrated Pest Management. J. Pest. Sci. 2014, 87, 385–405. [Google Scholar]
- Meats, A.; Holmes, H.M.; Kelly, G.L. Laboratory Adaptation of Bactrocera tryoni (Diptera: Tephritidae) Decreases Mating Age and Increases Protein Consumption and Number of Eggs Produced per Milligram of Protein. Bull. Entomol. Res. 2004, 94, 517–524. [Google Scholar] [CrossRef]
- Aceves-Aparicio, E.; Pérez-Staples, D.; Arredondo, J.; Corona-Morales, A.; Morales-Mávil, J.; Díaz-Fleischer, F. Combined Effects of Methoprene and Metformin on Reproduction, Longevity, and Stress Resistance in Anastrepha ludens (Diptera: Tephritidae): Implications for the Sterile Insect Technique. J. Econ. Entomol. 2021, 114, 142–151. [Google Scholar] [CrossRef]
- Belyi, A.A.; Alekseev, A.A.; Fedintsev, A.Y.; Balybin, S.N.; Proshkina, E.N.; Shaposhnikov, M.V.; Moskalev, A.A. The Resistance of Drosophila melanogaster to Oxidative, Genotoxic, Proteotoxic, Osmotic Stress, Infection, and Starvation Depends on Age According to the Stress Factor. Antioxidants 2020, 9, 1239. [Google Scholar] [CrossRef] [PubMed]
- Weldon, C.W.; Taylor, P.W. Desiccation Resistance of Adult Queensland Fruit Flies Bactrocera tryoni Decreases with Age. Physiol. Entomol. 2010, 35, 385–390. [Google Scholar] [CrossRef]
- Carey, J.R.; Harshman, L.G.; Liedo, P.; Müller, H.G.; Wang, J.L.; Zhang, Z. Longevity-Fertility Trade-Offs in the Tephritid Fruit Fly, Anastrepha ludens, across Dietary-Restriction Gradients. Aging Cell 2008, 7, 470–477. [Google Scholar] [CrossRef]
- Oviedo, A.; Nestel, D.; Papadopoulos, N.T.; Ruiz, M.J.; Prieto, S.C.; Willink, E.; Vera, M.T. Management of Protein Intake in the Fruit Fly Anastrepha fraterculus. J. Insect Physiol. 2011, 57, 1622–1630. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, W.; Tao, X.; Wang, D.; Lu, L.; He, Y. Using Two-Sex Life Table Traits to Assess the Fruit Preference and Fitness of Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2936–2945. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Nakas, C.T.; Carey, J.R.; Papadopoulos, N.T. Condition-Dependent Effects of Mating on Longevity and Fecundity of Female Medflies: The Interplay between Nutrition and Age of Mating. PLoS ONE 2013, 8, e70181. [Google Scholar] [CrossRef]
- Gerofotis, C.D.; Yuval, B.; Ioannou, C.S.; Nakas, C.T.; Papadopoulos, N.T. Polygyny in the Olive Fly—Effects on Male and Female Fitness. Behav. Ecol. Sociobiol. 2015, 69, 1323–1332. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Koskinioti, P.; Bourtzis, K.; Papadopoulos, N.T. Effect of Wolbachia Infection and Adult Food on the Sexual Signaling of Males of the Mediterranean Fruit Fly Ceratitis capitata. Insects 2022, 13, 737. [Google Scholar] [CrossRef] [PubMed]
- Shelly, T.E.; Kennelly, S. Influence of male diet on male mating success and longevity and female remating in the mediterranean fruit fly (diptera: Tephritidae) under laboratory conditions. Fla. Entomol. 2002, 85, 572–579. [Google Scholar] [CrossRef]
- Gregoriou, M.E.; Reczko, M.; Kakani, E.G.; Tsoumani, K.T.; Mathiopoulos, K.D. Decoding the Reproductive System of the Olive Fruit Fly, Bactrocera oleae. Genes 2021, 12, 355. [Google Scholar] [CrossRef] [PubMed]
- Herndon, L.A.; Wolfnert, M.F. A Drosophila Seminal Fluid Protein, Acp26Aa, Stimulates Egg Laying in Females for 1 Day after Mating. Proc. Natl. Acad. Sci. USA 1995, 92, 10114–10118. [Google Scholar] [CrossRef]
- Chen, E.H.; Hou, Q.L.; Wei, D.D.; Jiang, H.B.; Wang, J.J. Phenotypes, Antioxidant Responses, and Gene Expression Changes Accompanying a Sugar-Only Diet in Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). BMC Evol. Biol. 2017, 17, 194. [Google Scholar] [CrossRef] [PubMed]
- Nestel, D.; Papadopoulos, N.T.; Liedo, P.; Gonzales-Ceron, L.; Carey, J.R. Trends in Lipid and Protein Contents during Medfly Aging: An Harmonic Path to Death. Arch. Insect Biochem. Physiol. 2005, 60, 130–139. [Google Scholar] [CrossRef]
- Chen, D.; Han, H.L.; Li, W.J.; Wang, J.J.; Wei, D. Expression and Role of Vitellogenin Genes in Ovarian Development of Zeugodacus cucurbitae. Insects 2022, 13, 452. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balampekou, E.I.; Koveos, D.S.; Kapranas, A.; Menexes, G.C.; Kouloussis, N.A. The Roles of Mating, Age, and Diet in Starvation Resistance in Bactrocera oleae (Olive Fruit Fly). Insects 2023, 14, 841. https://doi.org/10.3390/insects14110841
Balampekou EI, Koveos DS, Kapranas A, Menexes GC, Kouloussis NA. The Roles of Mating, Age, and Diet in Starvation Resistance in Bactrocera oleae (Olive Fruit Fly). Insects. 2023; 14(11):841. https://doi.org/10.3390/insects14110841
Chicago/Turabian StyleBalampekou, Evangelia I., Dimitrios S. Koveos, Apostolos Kapranas, Georgios C. Menexes, and Nikos A. Kouloussis. 2023. "The Roles of Mating, Age, and Diet in Starvation Resistance in Bactrocera oleae (Olive Fruit Fly)" Insects 14, no. 11: 841. https://doi.org/10.3390/insects14110841