
Caddisflies (Trichoptera) of Protected Calcareous Fen Habitats: Assemblages, Environmental Drivers, Indicator Species, and Conservation Issues

 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Research Sites
2.2. Field and Laboratory Procedures
2.3. Data Analyses
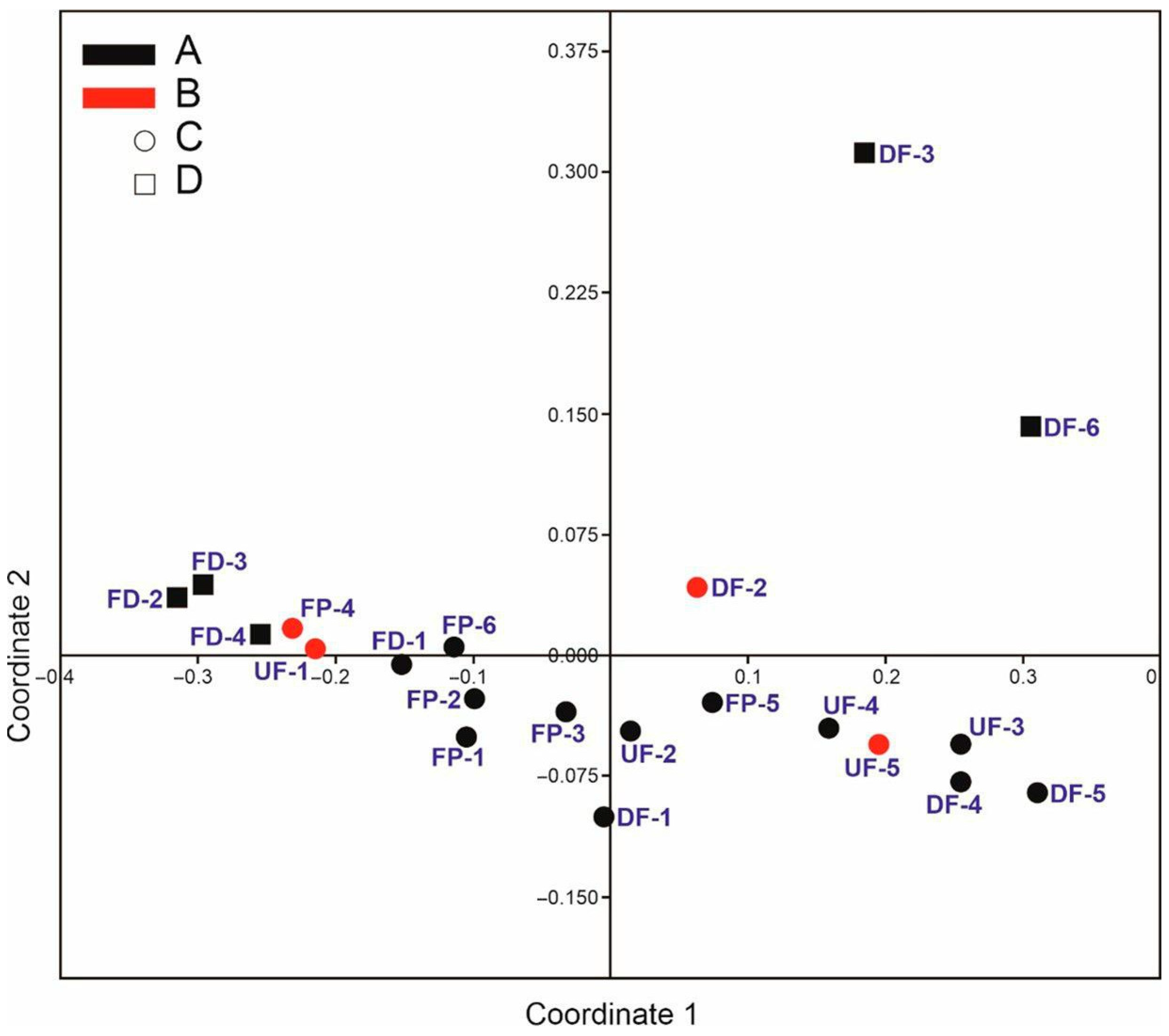
3. Results
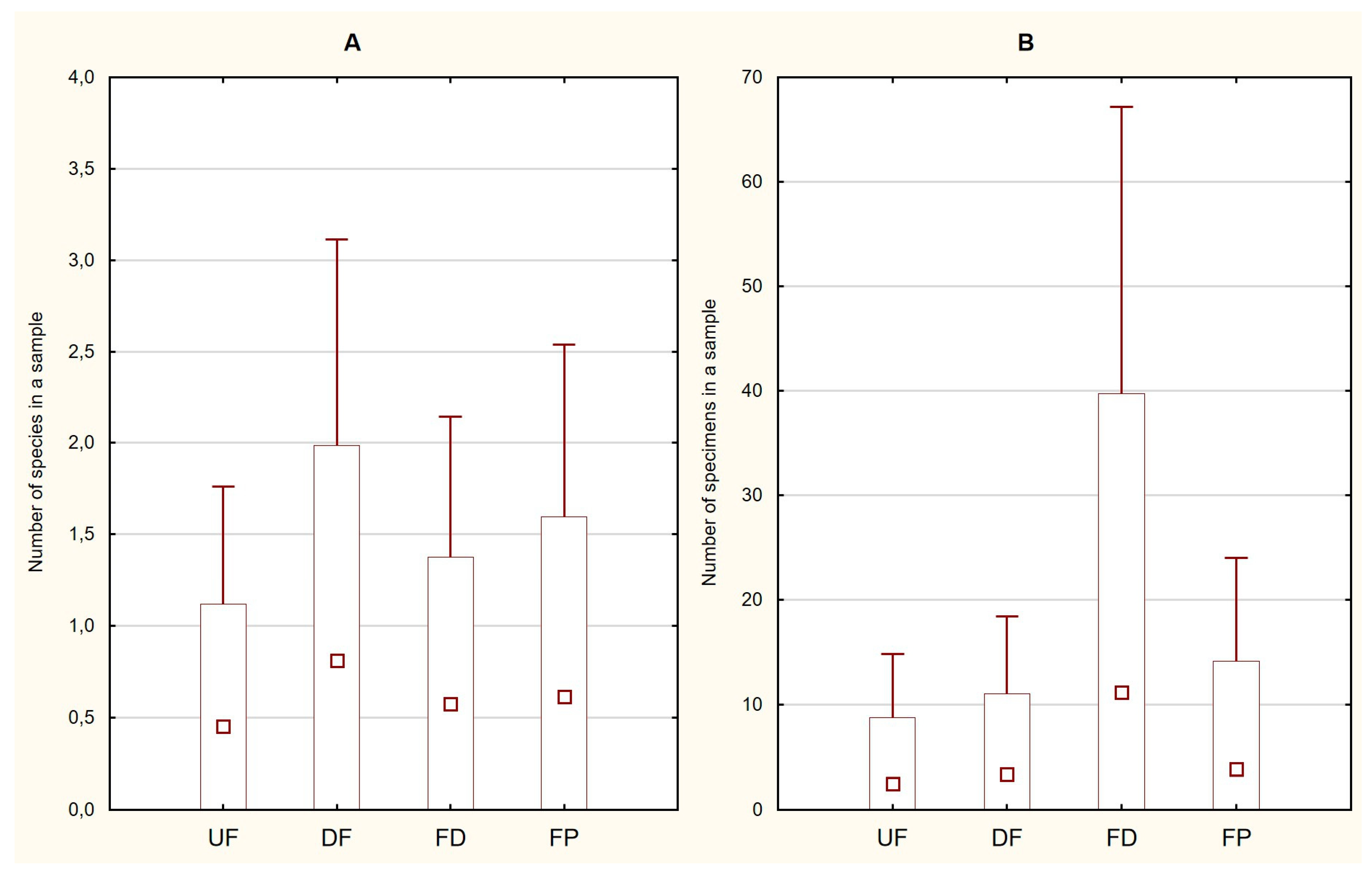
3.1. General Characteristics of the Trichopteran Assemblages and Ecological Groups
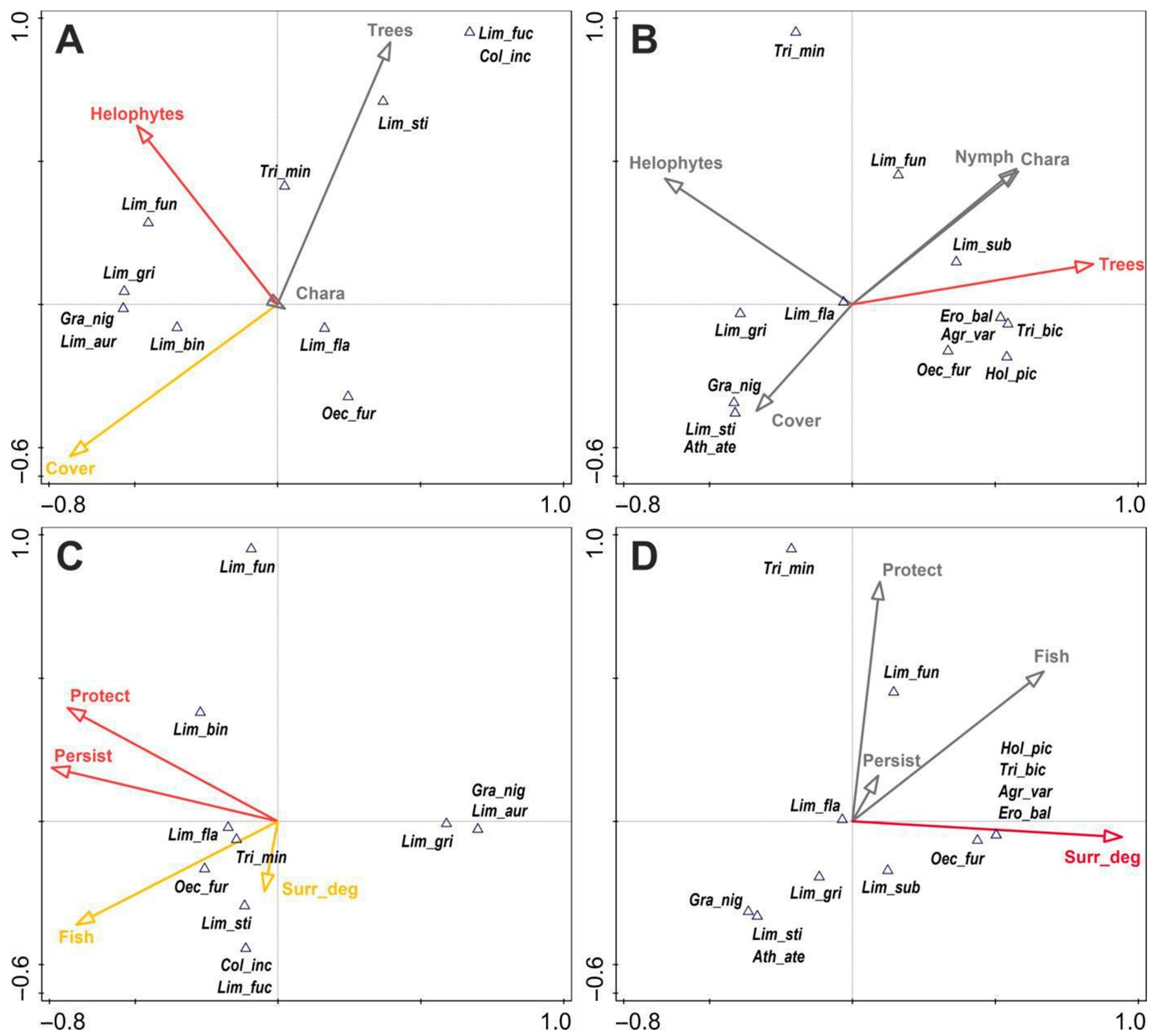
3.2. Environmental Factors Influencing the Trichopteran Assemblages
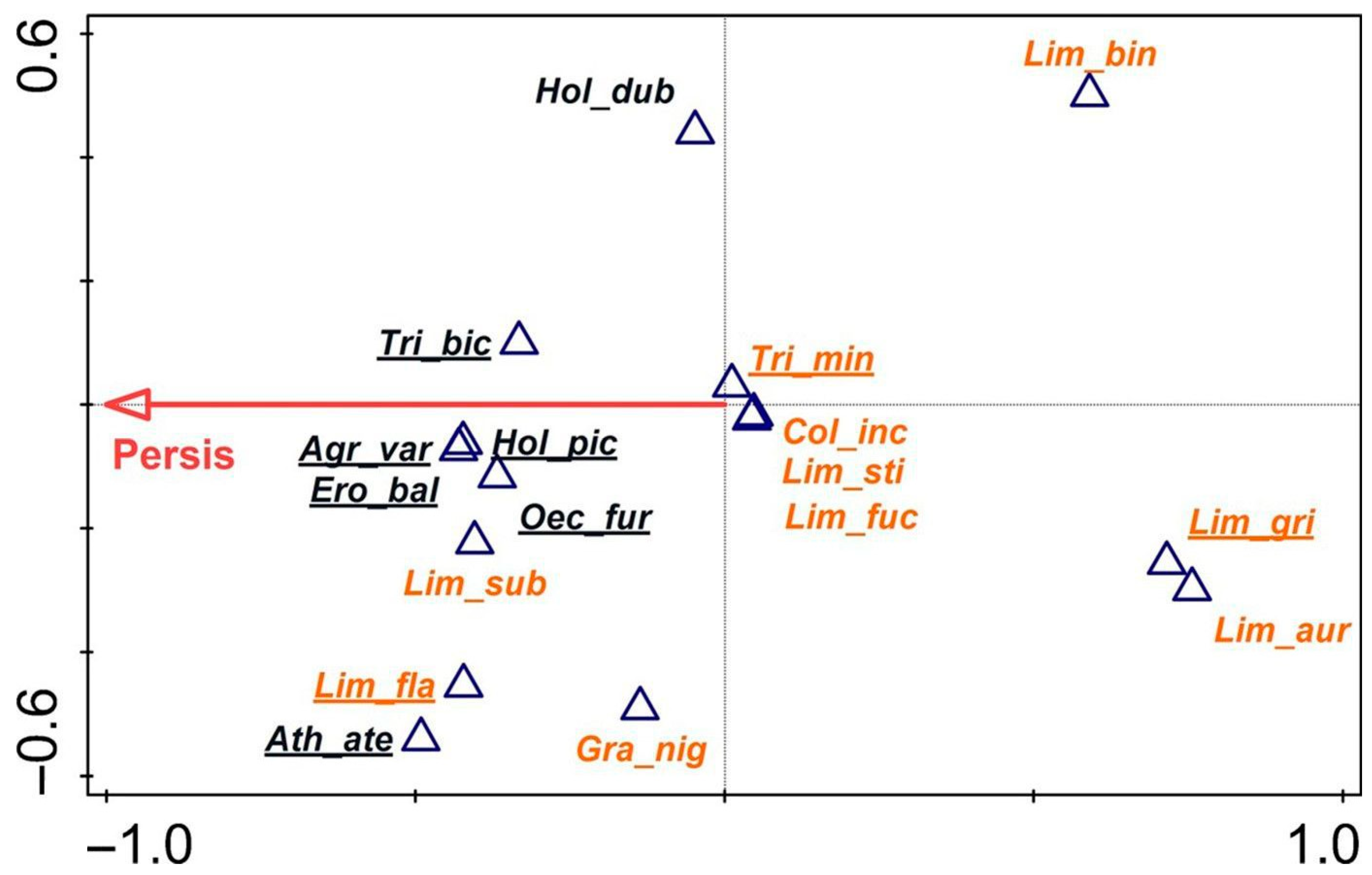
3.3. Protected Fens, Reference and Indicator Species
4. Discussion
4.1. Caddisflies of Calcareous Fen Habitats—Species Richness and Ecological Indices
4.2. Key Habitat Factors for Trichopterans in the Context of Protection and Conservation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamers, L.P.; Vile, M.A.; Grootjans, A.P.; Acreman, M.C.; van Diggelen, R.; Evans, M.G.; Richardson, C.J.; Rochefrot, L.; Kooijman, A.M.; Roelofs, J.G.M.; et al. Ecological restoration of rich fens in Europe and North America: From trial and error to an evidence-based approach. Biol. Rev. 2015, 90, 182–203. [Google Scholar]
- Batzer, D.; Wu, H.; Wheeler, T.; Eggert, S. Peatland invertebrates. In Invertebrates in Freshwater Wetlands: An International Perspective on Their Ecology; Batzer, D., Boix, D., Eds.; Springer: Cham, Switzerland, 2016; pp. 219–250. [Google Scholar]
- Grootjans, A.; Šefferová Stanová, V.; Jansen, A. Calcareous Mires of Slovakia: Landscape Setting, Management and Restoration Prospects; KNNV Uitgeverij: Zeist, The Netherlands, 2014. [Google Scholar]
- Stańko, R.; Wołejko, L.; Pawlaczyk, P. (Eds.) A Guidebook on Good Practices of Alkaline Fen Conservation; Klub Przyrodników Publishing House: Świebodzin, Poland, 2018. [Google Scholar]
- Ilnicki, P. Torfowiska i Torf; Wydawnictwo Akademii Rolniczej w Poznaniu: Poznań, Poland, 2002. [Google Scholar]
- Wołejko, L.; Pawlaczyk, P.; Stańko, R. (Eds.) Torfowiska Alkaliczne w Polsce—Zróżnicowanie, Zasoby, Ochrona; Wydawnictwo Klubu Przyrodników: Świebodzin, Poland, 2019. [Google Scholar]
- McBride, A.; Diack, I.; Droy, N.; Hamill, B.; Jones, P.; Schutten, J.; Skinner, A.; Street, M. The Fen Management Handbook; Scottish Natural Heritage: Perth, UK, 2011. [Google Scholar]
- Buczyński, P. Dragonflies (Odonata) of Anthropogenic Waters in Middle-Eastern Poland; Mantis: Olsztyn, Poland, 2015. [Google Scholar]
- Buczyńska, E.; Buczyński, P. Aquatic insects of man-made habitats: Environmental factors determining the distribution of Caddisflies (Trichoptera), Dragonflies (Odonata), and Beetles (Coleoptera) in Acidic Peat Pools. J. Insect Sci. 2019, 19, 17. [Google Scholar]
- Kalaninová, D.; Bulánková, E.; Šporka, F. Caddisflies (Trichoptera) as good indicators of environmental stress in mountain lotic ecosystems. Biologia 2014, 69, 1030–1045. [Google Scholar]
- Sun, Y.; Takemon, Y.; Yamashiki, Y. Freshwater spring indicator taxa of benthic invertebrates. Ecohydrol. Hydrobiol. 2020, 20, 622–631. [Google Scholar]
- Graf, W.; Murphy, J.; Dahl, J.; Zamora-Muñoz, C.; López-Rodríguez, M.J.; Schmidt-Kloiber, A.; Dataset “Trichoptera”. The Taxa and Autecology Database for Freshwater Organisms, Version 8.0. Available online: www.freshwaterecology.info (accessed on 8 September 2023).
- Morse, J.C.; Frandsen, P.B.; Graf, W.; Thomas, J.A. Diversity and ecosystem services of Trichoptera. Insects 2019, 10, e125. [Google Scholar]
- Wallace, I.D. A Review of the Status of the Caddis Flies (Trichoptera) of Great Britain. Species Status No. 27; Natural England Commissioned Report, Number 191; Natural England: Worcester, UK, 2016. [Google Scholar]
- Petruželová, J.; Bojková, J.; Hubáčková, L.; Šorfová, V.; Syrovátka, V.; Horsák, M. Factors explaining community contrast of Trichoptera assemblages at insular Western Carpathian spring fens to the adjacent headwaters. Int. Rev. Hydrobiol. 2020, 105, 20–32. [Google Scholar]
- Czachorowski, S.; Buczyński, P.; Walczak, U.; Pakulnicka, J. Gatunki osłonowe (parasolowe) w ochronie owadów. Przegl. Przyr. 2000, 11, 139–148. [Google Scholar]
- Kondracki, J. Geografia Regionalna Polski; PWN: Warsaw, Poland, 2000. [Google Scholar]
- Michalczyk, Z. Położenie Polesia Lubelskiego. In Polesie. Środowisko, Melioracje. Tom 3. Polesie Polskie; Urban, D., Dobrowolski, R., Jeznach, J., Eds.; Wydawnictwo SGGW: Warsaw, Poland, 2020; pp. 79–84. [Google Scholar]
- Solon, J.; Borzyszkowski, J.; Bidłasik, M.; Richling, A.; Badora, K.; Balon, J.; Brzezińska-Wójcik, T.; Chabudziński, Ł.; Dobrowolski, R.; Grzegorczyk, I.; et al. Physico-geographical mesoregions of Poland: Verification and adjustment of boundaries on the basis of contemporary spatial data. Geogr. Pol. 2018, 91, 143–170. [Google Scholar] [CrossRef]
- Herbichowa, M.; Wołejko, L. Torfowiska nakredowe (Cladietum marisci, Caricetum buxbaumii, Schoenetum nigricantis). In Wody Słodkie i Torfowiska. Poradniki Ochrony Siedlisk i Gatunków Natura 2000—Podręcznik Metodyczny; Herbich, J., Ed.; Ministerstwo Środowiska: Warsaw, Poland, 2004; pp. 163–171. [Google Scholar]
- Buczek, A. 7210 Torfowiska nakredowe. In Monitoring Siedlisk Przyrodniczych, Przewodnik Metodyczny, Część I; Mróz, W., Ed.; GIOŚ: Warsaw, Poland, 2010; pp. 161–173. [Google Scholar]
- EEA [European Environment Agency]. Calcareous Fens with Cladium Mariscus and Species of the Caricion Davallianae. Available online: https://eunis.eea.europa.eu/habitats/10149 (accessed on 8 September 2023).
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria, 1964. [Google Scholar]
- Regulation of the Minister of the Environment of September 9, 2011 on the list of alien plant and animal species which, if released into the natural environment, may threaten native species or natural habitats. Dz. Ustaw 2011, 210, pos. 1260.
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’eau Douce: Systématique, Biologie, Écologie; CNRS Editions: Paris, France, 2000. [Google Scholar]
- Hershkovitz, Y.; Dahm, V.; Lorenz, A.W.; Hering, D. A multi-trait approach for the identification and protection of European freshwater species that are potentially vulnerable to the impacts of climate change. Ecol. Indic. 2015, 50, 150–160. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral. Ecol. 1993, 18, 117–143. [Google Scholar]
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Szczęsny, B. Trichoptera Chruściki. In Czerwona Lista Zwierząt Ginących i Zagrożonych w Polsce; Głowaciński, Z., Ed.; Wydawnictwo Instytutu Ochrony Przyrody PAN: Kraków, Poland, 2002; pp. 76–79. [Google Scholar]
- Kubiak, M.; Stotzem, H.C.; Schütte, K.; Peters, R.S. A commented historic inventory of the caddisfly fauna (Trichoptera) of the fen complex “Eppendorfer Moor” in Northern Germany, based on material collected by G. Ulmer 1896–1907. Lauterbornia 2014, 77, 201–217. [Google Scholar]
- Hannigan, E.; Kelly-Quinn, M. Composition and structure of macroinvertebrate communities in contrasting open-water habitats in Irish peatlands: Implications for biodiversity conservation. Hydrobiologia 2012, 692, 19–28. [Google Scholar] [CrossRef]
- Brown, L.E.; Rachmunder, S.J.; Beadle, J.M.; Holden, J. Macroinvertebrate community assembly in pools created during peatland restoration. Sci. Total Environ. 2016, 569–570, 361–372. [Google Scholar]
- Boucenna, H.; Satour, A.; Hezil, W.; Taferghoust, M.; Samraoui, F.; Samraoui, B. Diversity, distribution, and conservation of the Trichoptera and their habitats in north-eastern Algeria. Aquat. Conserv. Mar. Freshw. Ecos. 2023, 33, 502–516. [Google Scholar]
- Painter, D. Macroinvertebrate distributions and the conservation calue of aquatic Coleoptera, Mollusca and Odonata in the ditches of traditionally managed and grazing fen at Wicken Fen, UK. J. Appl. Ecol. 1999, 36, 33–48. [Google Scholar] [CrossRef]
- Armitage, P.D.; Szostkiewicz, K.; Blackburn, J.H.; Nesbitt, I. Ditch communities: A major contributor to floodplain biodiversity. Aquat. Conserv. Mar. Freshw. Ecos. 2003, 13, 165–185. [Google Scholar]
- Langheinrich, U.; Tischew, S.; Gersberg, R.M.; Lüuderitz, V. Ditches and canals in management of fens: Opportunity or risk? A case study in the Drömling Natural Park, Germany. Wetl. Ecol. Manag. 2004, 12, 429–445. [Google Scholar]
- Clarke, S.J. Conserving freshwater biodiversity: The value, status and management of high quality ditch systems. J. Nat. Conserv. 2015, 24, 93–100. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Torma, A.; Bátori, Z.; Šeat, J.; Popović, M.; Gallé, R.; Gallé-Szpisjak, N.; Erdős, L.; Vinkó, T.; Kelemen, A.; et al. Turning old foes into new allies—Harnessing drainage canals for biodiversity conservation in a desiccated European lowland region. J. Appl. Ecol. 2022, 59, 89–102. [Google Scholar] [CrossRef]
- Falarz, M. (Ed.) Climate Change in Poland: Past, Present, Future; Springer Nature: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Grzybowski, M.; Glińska-Lewczuk, K. The principal threats to the peatlands habitats, in the continental bioregion of Central Europe—A case study of peatland conservation in Poland. J. Nat. Conserv. 2020, 53, 125778. [Google Scholar] [CrossRef]
- Grzywaczewski, G.; Cios, S.; Sparks, T.H.; Buczek, A.; Tryjanowski, P. Effect of habitat burning on the number of singing males of the Aquatic Warbler Acrocephalus paludicola. Acta Ornithol. 2014, 49, 175–185. [Google Scholar] [CrossRef]
- IUCN [International Union for Conservation of Nature]. The IUCN Red List of Threatened Species. Version 2022-2. 2022. Available online: https://www.iucnredlist.org (accessed on 8 September 2023).
- Buczyńska, E.; Czachorowski, S.; Buczyński, P. Issues concerning the conservation of the rare caddis Erotesis baltica McLachlan, 1877 (Trichoptera: Leptoceridae) in Poland. J. Ent. Res. Soc. 2015, 17, 87–95. [Google Scholar]
- Borisova, N.V.; Ruchin, A.B.; Khapugin, A.A.; Semishin, G.B. Non-metric multidimensional scaling analysis of composition of trichopterofauna from two protected areas (Republic of Mordovia, Russia). Inland Water Biol. 2023, 16, 16–27. [Google Scholar] [CrossRef]
- GDOŚ [Generalna Dyrekcja Ochrony Środowiska]. Geoserwis. Mapy. Available online: https://geoserwis.gdos.gov.pl/mapy/ (accessed on 8 September 2023).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Site | p | |||
|---|---|---|---|---|---|
| UF | DF | FD | FP | ||
| TEMP [°C] | 19.6 (7.27–33.29) | 17.5 (7.4–30.7) | 15.3 (7.4–26.6) | 15.9 (6.39–26.79) | >0.0001 |
| pH | 9.00 (706–10.48) | 8.50 (68.6–10.69) | 8.00 (6.94–10.50) | 8.20 (7.16–11.08) | 0.1239 |
| ORP [mV] | −172 (−453–206) | −169 (405–220) | −163 (−402–177) | −177 (−472–250) | 0.0014 |
| DO [mg·dm−3] | 3.20 (0.00–9.35) | 3.50 (0.0–10.60) | 3.79 (0.00–10.25) | 2.56 (0.00–10.42) | >0.0001 |
| EC [µS·cm−1] | 639 (138–1232) | 634 (294–2203) | 838 (372–3232) | 555 (205–1848) | 0.0032 |
| TDS [ppm] | 322 (84–616) | 366 (125–2041) | 422 (178–1616) | 274 (103–924) | >0.0001 |
| Species | Code | Sites | Habitats | |||
|---|---|---|---|---|---|---|
| UF | DF | FD | FP | |||
| Agrypnia varia (F.) | Agr_var | FP-2 | – | – | – | 0.004 |
| Athripsodes aterrimus (Stephens, 1836) | Ath_ate | FD-2 | – | – | 0.006 | – |
| Colpotaulius incisus (Curtis, 1834) | Col_inc | DF-1 | – | 0.028 | – | – |
| Erotesis baltica McLachlan, 1877 * | Ero_bal | FP-2 | – | – | – | 0.004 |
| Grammotaulius nigropunctatus (Retzius, 1783) | Gra_nig | DF-3, FD-2, FD-3 | – | 0.014 | 0.028 | – |
| Holocentropus dubius (Rambur, 1842) | Hol_dub | UF-2 | 0.005 | – | – | – |
| Holocentropus picicornis (Stephens, 1836) | Hol_pic | FP-1, FP-2 | – | – | – | 0.007 |
| Limnephilus auricula Curtis, 1834 | Lim_aur | DF-3 | – | 0.021 | – | – |
| Limnephilus binotatus Curtis, 1834 | Lim_bin | DF-4 | – | 0.007 | – | – |
| Limnephilus flavicornis (F.) | Lim_fla | all study sites | 0.636 | 0.330 | 2.861 | 0.956 |
| Limnephilus fuscicornis (Rambur, 1842) | Lim_fuc | DF-1 | – | 0.014 | – | – |
| Limnephilus fuscinervis (Zetterstedt, 1840)* | Lim_fun | UF-3, DF-2, FP-2, FP-4 | 0.005 | 0.056 | – | 0.007 |
| Limnephilus griseus (L.) | Lim_gri | DF-2, DF-3, DF-6, FD-3, FP-6 | – | 0.358 | 0.006 | 0.004 |
| Limnephilus stigma Curtis, 1834 | Lim_sti | DF-1, DF-5, FD-2 | – | 0.028 | 0.006 | – |
| Limnephilus subcentralis Brauer, 1857 | Lim_sub | FD-4, FP-3 | – | – | 0.006 | 0.004 |
| Oecetis furva (Rambur, 1842) | Oec_fur | UF-1, UF-2, FP-1, FP-2, FP-6 | 0.015 | – | – | 0.041 |
| Triaenodes bicolor (Curtis, 1834) | Tri_bic | FP-1, FP-2, FP-3 | – | – | – | 0.022 |
| Trichostegia minor (Curtis, 1834) | Tri_min | UF-2, UF-4, DF-1, DF-2, FP-4 | 0.015 | 0.014 | – | 0.007 |
| S= | 5 | 10 | 6 | 10 | ||
| N= | 132 | 124 | 524 | 285 | ||
| H= | 0.32 | 1.50 | 0.10 | 0.50 | ||
| E= | 0.27 | 0.44 | 0.18 | 0.16 | ||
| D= | 0.88 | 0.32 | 0.98 | 0.82 | ||
| Chao1= | 6 | 10 | 12 | 11.5 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buczyńska, E.; Tarkowski, A.; Sugier, P.; Płaska, W.; Zawal, A.; Janicka, A.; Buczyński, P. Caddisflies (Trichoptera) of Protected Calcareous Fen Habitats: Assemblages, Environmental Drivers, Indicator Species, and Conservation Issues. Insects 2023, 14, 850. https://doi.org/10.3390/insects14110850
Buczyńska E, Tarkowski A, Sugier P, Płaska W, Zawal A, Janicka A, Buczyński P. Caddisflies (Trichoptera) of Protected Calcareous Fen Habitats: Assemblages, Environmental Drivers, Indicator Species, and Conservation Issues. Insects. 2023; 14(11):850. https://doi.org/10.3390/insects14110850
Chicago/Turabian StyleBuczyńska, Edyta, Adam Tarkowski, Piotr Sugier, Wojciech Płaska, Andrzej Zawal, Anna Janicka, and Paweł Buczyński. 2023. "Caddisflies (Trichoptera) of Protected Calcareous Fen Habitats: Assemblages, Environmental Drivers, Indicator Species, and Conservation Issues" Insects 14, no. 11: 850. https://doi.org/10.3390/insects14110850
APA StyleBuczyńska, E., Tarkowski, A., Sugier, P., Płaska, W., Zawal, A., Janicka, A., & Buczyński, P. (2023). Caddisflies (Trichoptera) of Protected Calcareous Fen Habitats: Assemblages, Environmental Drivers, Indicator Species, and Conservation Issues. Insects, 14(11), 850. https://doi.org/10.3390/insects14110850