
The Entomopathogenic Fungus Conidiobolus coronatus Has Similar Effects on the Cuticular Free Fatty Acid Profile of Sensitive and Resistant Insects

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Fungus
2.3. Fungal Infection
2.4. Extraction of Samples, Derivatization and GC/MS Analysis
2.5. Statistics
3. Results
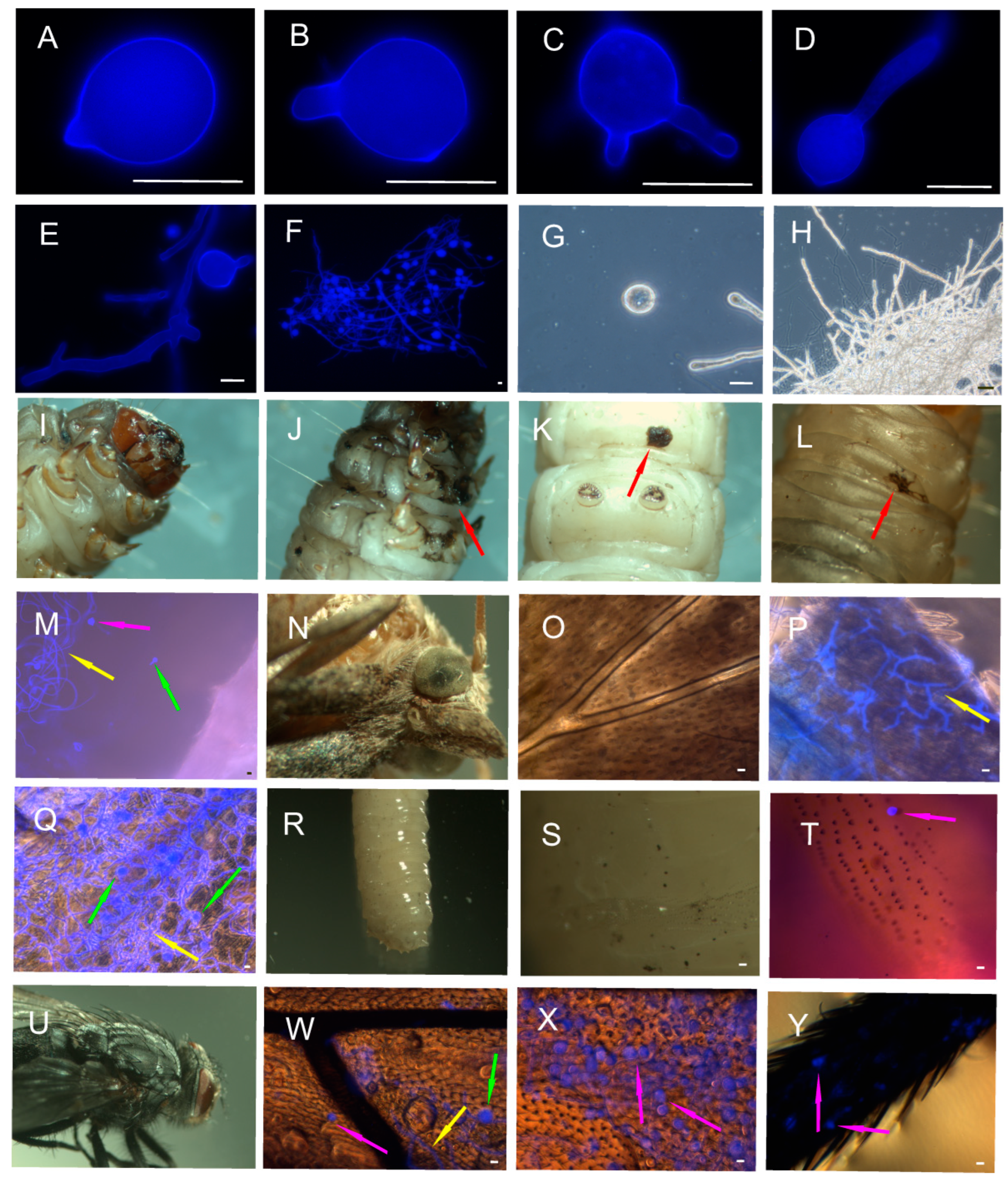
3.1. Effect of Fungal Infection on Insects
3.2. The Influence of C. coronatus on the Cuticular FFA Profiles of Galleria mellonella
3.3. The Influence of C. coronatus Treatment on the Cuticular FFA Profiles of Calliphora vicina
3.4. FFA Content in Conidiobolus coronatus Conidia
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bamisile, B.S.; Akutse, K.S.; Siddiqui, J.A.; Xu, Y. Model Application of Entomopathogenic Fungi as Alternatives to Chemical Pesticides: Prospects, Challenges, and Insights for Next-Generation Sustainable Agriculture. Front. Plant Sci. 2021, 12, 741804. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef] [PubMed]
- de Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Ferreira, J.M.; de Freitas Soares, F.E. Entomopathogenic fungi hydrolytic enzymes: A new approach to biocontrol? J. Nat. Pest. Res. 2023, 3, 100020. [Google Scholar] [CrossRef]
- Boguś, M.I.; Scheller, K. Extraction of an insecticidal protein fraction from the pathogenic fungus Conidiobolus coronatus. Acta Parasitol. 2002, 47, 66–72. [Google Scholar]
- Dromph, K.M.; Eilenberg, J.; Esbjerg, P. Natural occurrence of entomophthoralean fungi pathogenic to collembolans. J. Invert. Pathol. 2001, 78, 226–231. [Google Scholar] [CrossRef]
- Wieloch, W.; Boguś, M.I.; Ligęza, M.; Koszela-Piotrowska, I.; Szewczyk, A. Coronatin-1 isolated from entomopathogenic fungus Conidiobolus coronatus kills Galleria mellonella hemocytes in vitro and forms potassium channels in planar lipid membrane. Toxicon 2011, 58, 369–379. [Google Scholar] [CrossRef]
- Paszkiewicz, M.; Tyma, M.; Ligęza-Żuber, M.; Włóka, E.; Boguś, M.I.; Stepnowski, P. Trichothecenes production by entomopathogenic fungus Conidiobolus coronatus. Advan. Toxicol. Toxic Effects 2016, 1, 007–014. [Google Scholar] [CrossRef]
- Boguś, M.I.; Wieloch, W.; Ligęza-Żuber, M. Coronatin-2 from the entomopathogenic fungus Conidiobolus coronatus kills Galleria mellonella larvae and incapacitates hemocytes. Bull. Entomol. Res. 2017, 107, 66–76. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I. Harman and norharman, metabolites of the entomopathogenic fungus Conidiobolus coronatus (Entomophthorales), affect the serotonin levels and phagocytic activity of hemocytes, insect immunocompetent cells, in Galleria mellonella (Lepidoptera). Cell Biosci. 2019, 9, 29. [Google Scholar] [CrossRef]
- Kazek, M.; Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I. Dodecanol, metabolite of entomopathogenic fungus Conidiobolus coronatus, affects fatty acid composition and cellular immunity of Galleria mellonella and Calliphora vicina. Sci. Rep. 2021, 11, 15963. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, A.; Wrońska, A.K.; Kazek, M.; Boguś, M.I. Octanoic acid—An insecticidal metabolite of Conidiobolus coronatus (Entomopthorales) that affects two majors antifungal protection systems in Galleria mellonella (Lepidoptera): Cuticular lipids and hemocytes. Inter. J. Mol. Sci. 2022, 23, 5204. [Google Scholar] [CrossRef] [PubMed]
- Prasertphon, S.; Tanada, Y. The formation and circulation, in Galleria, of hyphal bodies of entomophtoraceous fungi. J. Invert. Pathol. 1968, 11, 260–280. [Google Scholar] [CrossRef]
- Boguś, M.I.; Szczepanik, M. Histopathology of Conidiobolus coronatus infection in Galleria mellonella larvae. Acta Parasitol. 2000, 45, 48–54. [Google Scholar]
- Boguś, M.I.; Kędra, E.; Bania, J.; Szczepanik, M.; Czygier, M.; Jabłoński, P.; Pasztaleniec, A.; Samborski, J.; Mazgajska, J.; Polanowski, A. Different defense strategies of Dendrolimus pini, Galleria mellonella, and Calliphora vicina against fungal infection. J. Insect Physiol. 2007, 53, 909–922. [Google Scholar] [CrossRef]
- Kazek, M.; Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I. Conidiobolus coronatus induces oxidative stress and autophagy response in Galleria mellonella larvae. PLoS ONE 2020, 15, e0228407. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I. Heat shock proteins (HSP 90, 70, 60, and 27) in Galleria mellonella (Lepidoptera) hemolymph are affected by infection with Conidiobolus coronatus (Entomophthorales). PLoS ONE 2020, 15, e0228556. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Kaczmarek, A.; Kazek, M.; Boguś, M.I. Infection of Galleria mellonella (Lepidoptera) larvae with the entomopathogenic fungus Conidiobolus coronatus (Entomophthorales) induces apoptosis of hemocytes and affects the concentration of eicosanoids in the hemolymph. Front. Physiol. 2022, 12, 774086. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I. The Changes in Mitochondrial Morphology and Physiology Accompanying Apoptosis in Galleria mellonella (Lepidoptera) Immunocompetent Cells during Conidiobolus coronatus (Entomophthorales) Infection. Int. J. Mol. Sci. 2023, 24, 10169. [Google Scholar] [CrossRef]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef]
- Mukherjee, K.; Vilcinskas, A. Transgenerational epigenetic inheritance in insects. In Translational Epigenetics, 2nd ed.; Tollefsbol, T.O., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 13, pp. 315–329. [Google Scholar]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic Fungi: Interactions and Applications. Encyclopedia 2022, 2, 646–656. [Google Scholar] [CrossRef]
- Bihal, R.; Al-Khayri, J.M.; Banu, A.N.; Kudesia, N.; Ahmed, F.K.; Sarkar, R.; Arora, A.; Abd-Elsalam, K.A. Entomopathogenic Fungi: An Eco-Friendly Synthesis of Sustainable Nanoparticles and Their Nanopesticide Properties. Microorganisms 2023, 11, 1617. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Zhang, S.; Keyhani, N.O. Targeting of insect epicuticular lipids by the entomopathogenic fungus Beauveria bassiana: Hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microb. 2013, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Boucias, D.G.; Pendland, J.C. Nutritional requirements for conidial germination of several host range pathotypes of the entomopathogenic fungus Nomuraea rileyi. J. Invertebr. Pathol. 1984, 43, 288–292. [Google Scholar] [CrossRef]
- Boucias, D.G.; Pendland, J.C.; Latge, J.P. Nonspecific factors involved in attachment of entomopathogenic Deuteromycetes to host insect cuticle. Appl. Environ. Microbiol. 1988, 54, 1795–1805. [Google Scholar] [CrossRef]
- Stephou, V.K.; Tjamos, S.E.; Paplomatas, E.J.; Athanassiou, C.G. Transformation and attachment of Beauveria bassiana conidia on the cuticle of Tribolium confusum and Sitophilus oryzae in conjunction with diatomaceous earth. J. Pest Sci. 2012, 85, 387–394. [Google Scholar] [CrossRef]
- Lord, J.C.; Howard, R.W. A proposed role for the cuticular fatty amides of Liposcelis bostrychophila (Psocoptera: Liposcelidae) in preventing adhesion of entomopathogenic fungi with dry-conidia. Mycopathologia 2004, 158, 211–217. [Google Scholar] [CrossRef]
- Boucias, D.; Latge, A.J. Nonspecific induction of germination of Conidiobolus obscurus and Nomuraearileyi with host and non-host cuticle extracts. J. Invert. Pathol. 1988, 51, 168–171. [Google Scholar] [CrossRef]
- El-Sayed, G.; Ignoffo, C.; Leathers, T. Effects of cuticle source and concentration on germination of conidia of two isolates of Nomuraea rileyi. Mycopathologia 1991, 113, 95–102. [Google Scholar] [CrossRef]
- Wang, C.; St. Leger, R.J. Developmental and Transcriptional Responses to Host and Nonhost Cuticles by the Specific Locust Pathogen Metarhizium anisopliae var. acridum. Eukar. Cell. 2005, 4, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.C.; Gołębiowski, M.; Pennisi, M.; Peterson, G.; García, J.J.; Manfrino, R.G.; López Lastra, C.C. Cuticle Fatty Acid Composition and Differential Susceptibility of Three Species of Cockroaches to the Entomopathogenic Fungi Metarhizium anisopliae (Ascomycota, Hypocreales). J. Econ Entomol. 2015, 108, 752–760. [Google Scholar] [CrossRef]
- Pedrini, N.; Zhang, S.; Juarez, M.P.; Keyhani, N.O. Molecular characterization and expression analysis of a suite of cytochrome P450 enzymes implicated in insect hydrocarbon degradation in the entomopathogenic fungus Beauveria bassiana. Microbiology 2010, 156, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- St Leger, R.J.; Cooper, R.M.; Charnley, A.K. Utilization of alkanes by entomopathogenic fungi. J. Invert. Pathol. 1988, 52, 356–359. [Google Scholar] [CrossRef]
- Napolitano, R.; Juarez, M.P. Entomopathogenous fungi degrade epicuticular hydrocarbons of Triatoma infestans. Arch. Biochem. Biophys. 1997, 344, 208–214. [Google Scholar] [CrossRef]
- Crespo, R.; Juarez, M.P.; Cafferata, L.F.R. Biochemical interaction between entomopathogenous fungi and their insect-host-like hydrocarbons. Mycologia 2000, 92, 528–536. [Google Scholar] [CrossRef]
- Pedrini, N.; Crespo, R.; Juarez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2007, 146, 124–137. [Google Scholar] [CrossRef]
- Da Silva, W.O.B.; Santi, L.; Correa, A.P.F.; Silva, L.A.D.; Bresciani, F.R.; Schrank, A.; Vainstein, M.H. The entomopathogen Metarhizium anisopliae can modulate the secretion of lipolytic enzymes in response to different substrates including components of arthropod cuticle. Fungal Biol. 2010, 114, 911–916. [Google Scholar] [CrossRef]
- Crespo, R.; Juarez, M.P.; Dal Bello, G.M.; Padin, S.; Fernandez, G.C.; Pedrini, N. Increased mortality of Acanthoscelides obtectus by alkane-grown Beauveria bassiana. BioControl 2002, 47, 685–696. [Google Scholar] [CrossRef]
- Jarrold, S.L.; Moore, D.; Potter, U.; Charnley, A.K. The contribution of surface waxes to pre-penetration growth of an entomopathogenic fungus on host cuticle. Mycol. Res. 2007, 111, 240–249. [Google Scholar] [CrossRef]
- Lecuona, R.; Riba, G.; Cassier, P.; Clement, J.L. Alterations of insect epicuticular hydrocarbons during infection with Beauveria bassiana or B. brongniartii. J. Invertebr. Pathol. 1991, 58, 10–18. [Google Scholar] [CrossRef]
- Kirkland, B.H.; Cho, E.M.; Keyhani, N.O. Differential susceptibility of Amblyomma maculatum and Amblyomma americanum (Acari: Ixodidea) to the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae. Biol. Control 2004, 31, 414–421. [Google Scholar] [CrossRef]
- Boguś, M.I.; Czygier, M.; Gołębiowski, M.; Kędra, E.; Kucińska, J.; Mazgajska, J.; Samborski, J.; Wieloch, W.; Włóka, E. Effects of insect cuticular fatty acids on in vitro growth and pathogenicity of the entomopathogenic fungus Conidiobolus coronatus. Exper. Parasit. 2010, 125, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Włóka, E.; Boguś, M.I.; Wrońska, A.K.; Drozdowski, M.; Kaczmarek, A.; Sobich, J.; Gołębiowski, M. Insect cuticular compounds affect Conidiobolus coronatus (Entomopthorales) sporulation and the activity of enzymes involved in fungal infection. Sci. Rep. 2022, 12, 13641. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I.; Włóka, E.; Kazek, M.; Kaczmarek, A.; Zalewska, K. Cuticular fatty acids of Galleria mellonella (Lepidoptera) inhibit fungal enzymatic activities of pathogenic Conidiobolus coronatus. PLoS ONE 2018, 13, e0192715. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Boguś, M.I.; Włóka, E.; Wrońska, A.K.; Krawiel, A.; Kazek, M.; Zalewska, K.; Kłocińska-Biały, K.; Sobocińska, M.; Gliniewicz, A.; et al. The interaction between cuticle free fatty acids (FFAs) of the cockroaches Blattella germanica and Blatta orientalis and hydrolases produced by the entomopathogenic fungus Conidiobolus coronatus. PLoS ONE 2020, 15, e0235785. [Google Scholar] [CrossRef]
- Smith, R.J.; Grula, E.A. Toxic components on the larval surface of the Corn-Earworm (Heliothis zea) and their effects on germination and growth of Beauveria bassiana. J. Invertebr. Pathol. 1982, 39, 15–22. [Google Scholar] [CrossRef]
- Saito, T.; Aoki, J. Toxicity of free fatty acids on the larval surfaces of 2 Lepidopterous insects towards Beauveria bassiana (Bals) Vuill and Paecilomyces fumosoroseus (Wize) Brown Et Smith (Deuteromycetes, Moniliales). Appl. Entomol. Zool. 1983, 18, 225–233. [Google Scholar] [CrossRef]
- Sosa-Gomez, D.R.; Boucias, D.G.; Nation, J.L. Attachment of Metarhizium anisopliae to the southern green stink bug Nezara viridula cuticle and fungistatic effect of cuticular lipids and aldehydes. J. Invertebr. Pathol. 1997, 69, 31–39. [Google Scholar] [CrossRef]
- Gross, J.; Muller, C.; Vilcinskas, A.; Hilker, M. Antimicrobial activity of exocrine glandular secretions, hemolymph, and larval regurgitate of the mustard leaf beetle Phaedon cochlearia. J. Invertebr. Pathol. 1998, 72, 296–303. [Google Scholar] [CrossRef]
- Gross, J.; Schumacher, K.; Schmidtberg, H.; Vilcinskas, A. Protected by fumigants: Beetle perfumes in antimicrobial defense. J. Chem. Ecol. 2008, 34, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, M.L.; Girotti, J.R.; Mijailovsky, S.J.; Pedrini, N.; Juarez, M.P. Volatile secretions and epicuticular hydrocarbons of the beetle Ulomoides dermestoides. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Ment, D.; Churchill, A.C.L.; Gindin, G.; Belausov, E.; Glazer, I.; Rehner, S.A.; Rot, A.; Donzelli, B.G.G.; Samish, M. Resistant ticks inhibit Metarhizium infection prior to haemocoel invasion by reducing fungal viability on the cuticle surface. Environ. Microbiol. 2012, 14, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, A.; Szadziewski, R.; Stepnowski, P.; Boros-Majewska, J.; Gabriel, I.; Dawgul, M.; Kamysz, W.; Sosnowska, D.; Gołębiowski, M. Composition and antimicrobial activity of fatty acids detected in the hygroscopic secretion collected from the secretory setae of larvae of the biting midge Forcipomyia nigra (Diptera: Ceratopogonidae). J. Insect Physiol. 2012, 58, 1265–1276. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Dawgul, M.; Kamysz, W.; Boguś, M.I.; Stepnowski, P. The antifungal activity of the cuticular and internal fatty acid methyl esters and alcohols in Calliphora vomitoria. Parasitology 2013, 140, 972–985. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Urbanek, A.; Oleszczak, A.; Dawgul, M.; Kamysz, W.; Boguś, M.I.; Stepnowski, P. The antifungal activity of fatty acids of all stages of Sarcophaga carnaria L. (Diptera: Sarcophagidae). Microb. Res. 2014, 169, 279–286. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Urbanek, A.; Dawgul, M.; Kamysz, W.; Boguś, M.; Sosnowska, D.; Stepnowski, P. Antimicrobial activity of untypical lipid compounds in the cuticular and internal lipids of four fly species. J. Appl. Microb. 2014, 116, 269–287. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Urbanek, A.; Dawgul, M.; Kamysz, W.; Boguś, M.I.; Stepnowski, P. Identification and antifungal activity of the novel organic compounds found in cuticular and internal lipids of medically important flies. Microb. Res. 2015, 170, 213–222. [Google Scholar] [CrossRef]
- Kerwin, J.L. Fatty acid regulation of the germination of Erynia. J. Microb. 1984, 30, 158–161. [Google Scholar]
- Keyhani, N.O. Lipid biology in fungal stress and virulence: Entomopathogenic fungi. Fungal Biol. 2018, 122, 420–429. [Google Scholar] [CrossRef]
- Kett, S.; Pathak, A.; Turillazzi, S.; Cavalieri, D.; Marvasi, M. Antifungals, arthropods and antifungal resistance prevention: Lessons from ecological interactions. Proc. Biol. Sci. 2021, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Boguś, M.I.; Włóka, E.; Wrońska, A.; Kaczmarek, A.; Kazek, M.; Zalewska, K.; Ligęza-Żuber, M.; Gołębiowski, M. Cuticle hydrolysis of four medically important fly species by enzymes of the entomopathogenic fungus Conidiobolus coronatus. Med. Vet. Entomol. 2017, 31, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Arias-Robledo, G.; Stevens, J.R.; Wall, R. Spatial and temporal habitat partitioning by calliphorid blowflies. Med. Vet. Entomol. 2019, 33, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Stoffolano, J.G., Jr. Synanthropic Flies-A Review Including How They Obtain Nutrients, along with Pathogens, Store Them in the Crop and Mechanisms of Transmission. Insects 2022, 13, 776. [Google Scholar] [CrossRef]
- Kazek, M.; Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I. Diet influences the bacterial and free fatty acid profiles of the cuticle of Galleria mellonella larvae. PLoS ONE 2019, 14, e0211697. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Wrońska, A.K.; Kazek, M.; Boguś, M.I. Metamorphosis-related changes in the free fatty acid profiles of Sarcophaga (Liopygia) argyrostoma (Robineau-Desvoidy, 1830). Sci. Rep. 2020, 10, 17337. [Google Scholar] [CrossRef]
- Sehnal, F. A critical study of the biome and biometry of the wax moth Galleria mellonella raised in varying conditions. Zeit. Wiss. Zool. 1966, 174, 53–82. [Google Scholar]
- Callaghan, A.A. Light and spore discharge in Entomophthorales. Trans. Brit. Mycol. Soc. 1969, 53, 87–97. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Mun, S.; Noh, M.Y.; Geisbrecht, E.R.; Arakane, Y. Insect Cuticular Chitin Contributes to Form and Function. Curr. Pharm. Des. 2020, 26, 3530–3545. [Google Scholar] [CrossRef]
- Moussian, B. Recent advances in understanding mechanisms of insect cuticle differentiation. Insect Bioch. Mol. Biol. 2010, 40, 363–375. [Google Scholar] [CrossRef]
- Zhang, W.; Tettamanti, G.; Bassal, T.; Heryanto, C.; Eleftherianos, I.; Mohamed, A. Regulators and signalling in insect antimicrobial innate immunity: Functional molecules and cellular pathways. Cell. Signal. 2021, 83, 110003. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, A.; Boguś, M.I. The Impact of the Entomopathogenic Fungus Conidiobolus coronatus on the Free Fatty Acid Profile of the Flesh Fly Sarcophaga argyrostoma. Insects 2021, 12, 970. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Nguyen, S.H.; Webb, H.K.; Hasan, J.; Truong, V.K.; Lamb, R.N.; Duan, X.; Tobin, M.J.; Mahon, T.J.; Crawford, R.J. Molecular Organization of the Nanoscale Surface Structures of the Dragonfly Hemianax papuensis Wing Epicuticle. PLoS ONE 2013, 8, e67893. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.; Vilela, R.; Voelz, K.; Ibrahim, A.S.; Voigt, K.; Lee, S.C. Human Fungal Pathogens of Mucorales and Entomophthorales. Cold Spring Harb Perspect Med. 2014, 5, a019562. [Google Scholar] [CrossRef]
- Dutta, S.; Sarkar, S.; Linka, U.; Dora, S. Conidiobolomycosis: A case report of rare fungal infection from the eastern India. Indian Dermatol. Online J. 2015, 6, 393–395. [Google Scholar] [CrossRef]
- Carmo, P.M.S.D.; Uzal, F.A.; Pedroso, P.M.O.; Riet-Correa, F. Conidiobolomycosis, cryptococcosis, and aspergillosis in sheep and goats: A review. J. Vet. Diagn. Invest. 2020, 32, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate humoral immune defences in mammals and insects: The same, with differences? Virulence 2018, 9, 1625–1639. [Google Scholar] [CrossRef]
- Baio, J.E.; Jaye, C.; Sullivan, E.; Rasmussen, M.H.; Fischer, D.A.; Gorb, S.; Weidner, T. NEXAFS imaging to characterize the physio-chemical composition of cuticle from African Flower Scarab Eudicella gralli. Nat Commun. 2019, 10, 4758. [Google Scholar] [CrossRef]
- Jafarpour, M.; Eshghi, S.; Darvizeh, A.; Gorb, S.; Rajabi, H. Functional significance of graded properties of insect cuticle supported by an evolutionary analysis. J. R. Soc. Interface 2020, 17, 20200378. [Google Scholar] [CrossRef]
- Jullien, A.; Neradovskiy, M.; Mitov, M. Hyperspectral topography of the twisted, cholesteric patterns of an insect cuticle under various conditions of helix obliquity. APL Photonics 2020, 5, 096102. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Insect Species | Larvae | Adults |
|---|---|---|
| Galleria mellonella | 33 ± 11 | 8 ± 11 |
| Calliphora vicina | 5 ± 7 | 4 ± 6 |
| Galleria mellonella Larvae (µg/g of Body Mass ± SD) | Galleria mellonella Adults (µg/g of Body Mass ± SD) | |||||
|---|---|---|---|---|---|---|
| FFA | Control | Fungal Infection 24 hpi | Fungal Infection 48 hpi | Control | Fungal Infection 24 hpi | Fungal Infection 48 hpi |
| C4:0 | 0.72 ± 0.28 | ND | ND | ND | ND | ND |
| C5:0 | 0.36 ± 0.13 | ND | ND | ND | ND | ND |
| C6:0 | 1.11 ± 0.47 | 1.95 ± 0.69 | 1.29 ± 0.65 | 2.04 ± 1.03 | 0.67 ± 0.25 | 0.03 ± 0 |
| C7:0 | 0.96 ± 0.34 | 0.90 ± 0.92 | 0.68 ± 0.23 | ND | 0.60 ± 0.57 | 0.02 ± 0 |
| C8:0 | 1.63 ± 0.05 A | 2.93 ± 1.68 | 1.80 ± 0.35 | 4.36 ± 1.47 A,l | 2.60 ± 0.69 m | 0.27 ± 0.01 l,m |
| C9:0 | 1.34 ± 0.17 B | 3.36 ± 1.39 a | 3.86 ± 0.85 a | 112.02 ± 6.48 B,n | 44.36 ± 1.17 n | 4.82 ± 0.20 n |
| C10:0 | 0.96 ± 0.30 b | 3.04 ± 0.98 b,c | 0.75 ± 0.17 c | 4.37 ± 3.18 | 3.31 ± 1.21 o | 0.17 ± 0 o |
| C11:0 | ND | ND | ND | 1268.38 ± 361.77 p | 424.06 ± 10.48 p | 29.71 ± 0.67 p |
| C12:0 | ND | 214.66 ± 24.19 | ND | 23.94 ± 5.77 q | 20.87 ± 1.70 r | 0.32 ± 0.01 q,r |
| C13:0 | ND | 121.50 ± 13.35 d | 12.84 ± 1.59 d | 7.92 ± 0.25 s | 52.19 ± 2.30 s | 0.64 ± 0.03 s |
| C14:1 | 2.02 ± 1.35 | ND | ND | 12.12 ± 6.61 | ND | ND |
| C14:0 | 28.69 ± 9.39 e | 519.57 ± 41.47 e | 71.73 ± 9.65 e | 43.94 ± 13.15 t | 209.09 ± 4.93 t | 1.28 ± 0.07 t |
| C15:0 | 7.12 ± 5.79 f | 151.04 ± 13.18 f,g | 14.68 ± 7.35 g | 17.93 ± 11.24 u | 55.01 ± 2.19 u | 0.36 ± 0 u |
| C16:1 | 18.34 ± 13.72 C,h | 90.37 ± 14.67 h | 41.63 ± 3.67 h | 264.61 ± 9.36 C,v | ND | 1.34 ± 0.03 v |
| C16:0 | 329.52 ± 154.13 D,i | 4680.44 ± 273.90 i | 1676.10 ± 246.23 i | 3687.79 ± 323.09 D,w | 905.38 ± 14.58 w | 51.54 ± 2.02 w |
| C18:2 | ND | ND | ND | 62.52 ± 84.21 | ND | ND |
| C18:1 | 80.19 ± 49.57 E,j | 410.94 ± 21.45 j | 238.09 ± 38.94 j | 730.16 ± 142.87 E,x | 315.02 ± 17.77 x | 9.92 ± 0.07 x |
| C18:0 | 12.27 ± 2.72 F | 40.74 ± 24.93 | ND | 168.87 ± 10.78 F,y | 22.27 ± 6.44 y | 1.14 ± 0 y |
| C20:1 | 1.33 ± 0.37 G | ND | ND | 6.29 ± 2.83 G | ND | ND |
| Sum of FFAs | 485.49 ± 238.78 H,k | 6241.45 ± 432.83 k | 2063.45 ± 309.69 k | 6417.28 ± 984.12 H,z | 2055.45 ± 64.33 z | 101.58 ± 3.13 z |
| Calliphora vicina Larvae (µg/g of Body Mass ± SD) | Calliphora vicina Adults (µg/g of Body Mass ± SD) | |||||
|---|---|---|---|---|---|---|
| FFA | Control | Fungal Infection 24 hpi | Fungal Infection 48 hpi | Control | Fungal Infection 24 hpi | Fungal Infection 48 hpi |
| C6:0 | ND | ND | ND | 0.09 ± 0.02 i | 0.51 ± 0.11 i | ND |
| C8:0 | ND | ND | ND | 0.12 ± 0.01 j | 1.19 ± 0.83 j | ND |
| C9:0 | ND | ND | 6.28 ± 1.41 | 0.45 ± 0.02 k | 1.24 ± 0.03 k | ND |
| C10:0 | ND | ND | 3.29 ± 0.59 | 0.13 ± 0.02 | ND | ND |
| C12:0 | 0.25 ± 0.13 a | 24.48 ± 1.12 a | 117.65 ± 36.11 a | ND | 2.29 ± 0.11 | ND |
| C13:0 | ND | 54.57 ± 1.23 b | 245.09 ± 71.08 b | ND | 2.83 ± 0.09 l | 20.46 ± 11.01 l |
| C14:1 | 0.49 ± 0.23 | ND | ND | 0.39 ± 0.06 | ND | ND |
| C14:0 | 1.06 ± 0.48 c | 215.08 ± 6.73 c | 1069.82 ± 302.58 c | 1.41 ± 0.06 m | 18.01 ± 1.08 m | 193.11 ± 39.11 m |
| C15:0 | ND | 60.41 ± 3.28 | 175.54 ± 113.09 | 0.49 ± 0.01 n | 3.46 ± 0.24 n | 28.12 ± 4.97 n |
| C16:1 | 24.46 ± 3.19 d | 76.69 ± 3.92 d | 150.13 ± 54.48 d | 23.84 ± 1.57 o | 17.89 ± 0.91 o | 295.08 ± 67.26 o |
| C16:0 | 9.73 ± 0.82 A,e | 609.55 ± 26.15 e | 2053.95 ± 661.04 e | 21.81 ± 0.85 A,p | 72.08 ± 2.99 p | 519.80 ± 100.70 p |
| C17:1 | 0.55 ± 0.08 B | ND | ND | 0.19 ± 0.02 B | ND | ND |
| C17:0 | ND | ND | ND | 0.77 ± 0.12 | ND | ND |
| C18:2 | ND | ND | ND | 2.19 ± 0.05 | ND | ND |
| C18:1 | 6.04 ± 0.49 C,f | 120.37 ± 2.73 f | 315.81 ± 34.52 f | 26.98 ± 1.61 C,q | 61.70 ± 1.69 q | 344.02 ± 107.75 q |
| C18:0 | 2.23 ± 0.27 D,g | 21.20 ± 0.95 g | 102.40 ± 20.68 g | 3.82 ± 0.05 D,r | 23.81 ± 1.32 r | 98.15 ± 22.49 r |
| C20:0 | ND | ND | ND | 0.85 ± 0.55 | ND | ND |
| C22:0 | ND | ND | ND | 2.88 ± 0.04 | ND | ND |
| C24:0 | ND | ND | ND | 1.71 ± 0.19 | ND | ND |
| C26:0 | ND | ND | ND | ND | ND | ND |
| Sum of FFAs | 44.82 ± 5.70 E,h | 1182.35 ± 46.12 h | 4239.99 ± 1295.59 h | 88.13 ± 5.29 E,s | 205.02 ± 9.44 s | 1498.75 ± 353.32 s |
| FFA Concentration (fg/Conidium ± SD) | ||
|---|---|---|
| FFA | Non-Germinated Spores | Germinated Spores |
| C6:0 | 149 ± 7 a | 83 ± 8 a |
| C7:0 | 21 ± 2 | 17 ± 10 |
| C8:0 | 70 ± 5 b | 27 ± 1 b |
| C9:0 | 188 ± 2 c | 49 ± 2 c |
| C10:0 | 53 ± 3 d | 9 ± 1 d |
| C11:0 | 33 ± 3 e | 6 ± 2 e |
| C12:0 | 1631 ± 71 f | 82 ± 6 f |
| C13:0 | 2695 ± 97 g | 169 ± 2 g |
| C14:1 | 47 ± 6 | ND |
| C14:0 | 14,735 ± 242 h | 2490 ± 62 h |
| C15:1 | 20 ± 6 | ND |
| C15:0 | 3507 ± 103 i | 1063 ± 51 i |
| C16:1 | 907 ± 36 j | 76 ± 6 j |
| C16:0 | 15,797 ± 172 k | 6928 ± 105 k |
| C17:1 | 1000 ± 44 l | 138 ± 8 l |
| C17:0 | 550 ± 38 m | 223 ± 10 m |
| C18:3 | 1112 ± 98 n | 90 ± 8 n |
| C18:2 | 2203 ± 144 o | 767 ± 148 o |
| C18:1 | 12,346 ± 439 p | 3571 ± 289 p |
| C18:0 | 2692 ± 108 q | 1913 ± 82 q |
| C19:1 | ND | 53 ± 8 |
| C19:0 | ND | 28 ± 1 |
| C20:4 | 4068 ± 123 | ND |
| C20:3 | ND | 531 ± 4 |
| C20:1 | ND | 236 ± 83 |
| C20:0 | ND | 81 ± 12 |
| C22:1 | ND | 77 ± 32 |
| C22:0 | ND | 130 ± 25 |
| C24:1 | ND | 485 ± 182 |
| C24:0 | ND | 267 ± 84 |
| Sum of FFAs | 63,827 ± 1643 r | 19,588 ± 1015 r |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boguś, M.I.; Kazek, M.; Drozdowski, M.; Kaczmarek, A.; Wrońska, A.K. The Entomopathogenic Fungus Conidiobolus coronatus Has Similar Effects on the Cuticular Free Fatty Acid Profile of Sensitive and Resistant Insects. Insects 2023, 14, 895. https://doi.org/10.3390/insects14110895
Boguś MI, Kazek M, Drozdowski M, Kaczmarek A, Wrońska AK. The Entomopathogenic Fungus Conidiobolus coronatus Has Similar Effects on the Cuticular Free Fatty Acid Profile of Sensitive and Resistant Insects. Insects. 2023; 14(11):895. https://doi.org/10.3390/insects14110895
Chicago/Turabian StyleBoguś, Mieczysława Irena, Michalina Kazek, Mikołaj Drozdowski, Agata Kaczmarek, and Anna Katarzyna Wrońska. 2023. "The Entomopathogenic Fungus Conidiobolus coronatus Has Similar Effects on the Cuticular Free Fatty Acid Profile of Sensitive and Resistant Insects" Insects 14, no. 11: 895. https://doi.org/10.3390/insects14110895
APA StyleBoguś, M. I., Kazek, M., Drozdowski, M., Kaczmarek, A., & Wrońska, A. K. (2023). The Entomopathogenic Fungus Conidiobolus coronatus Has Similar Effects on the Cuticular Free Fatty Acid Profile of Sensitive and Resistant Insects. Insects, 14(11), 895. https://doi.org/10.3390/insects14110895