
Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio molitor Larvae Infected by Beauveria bassiana

Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
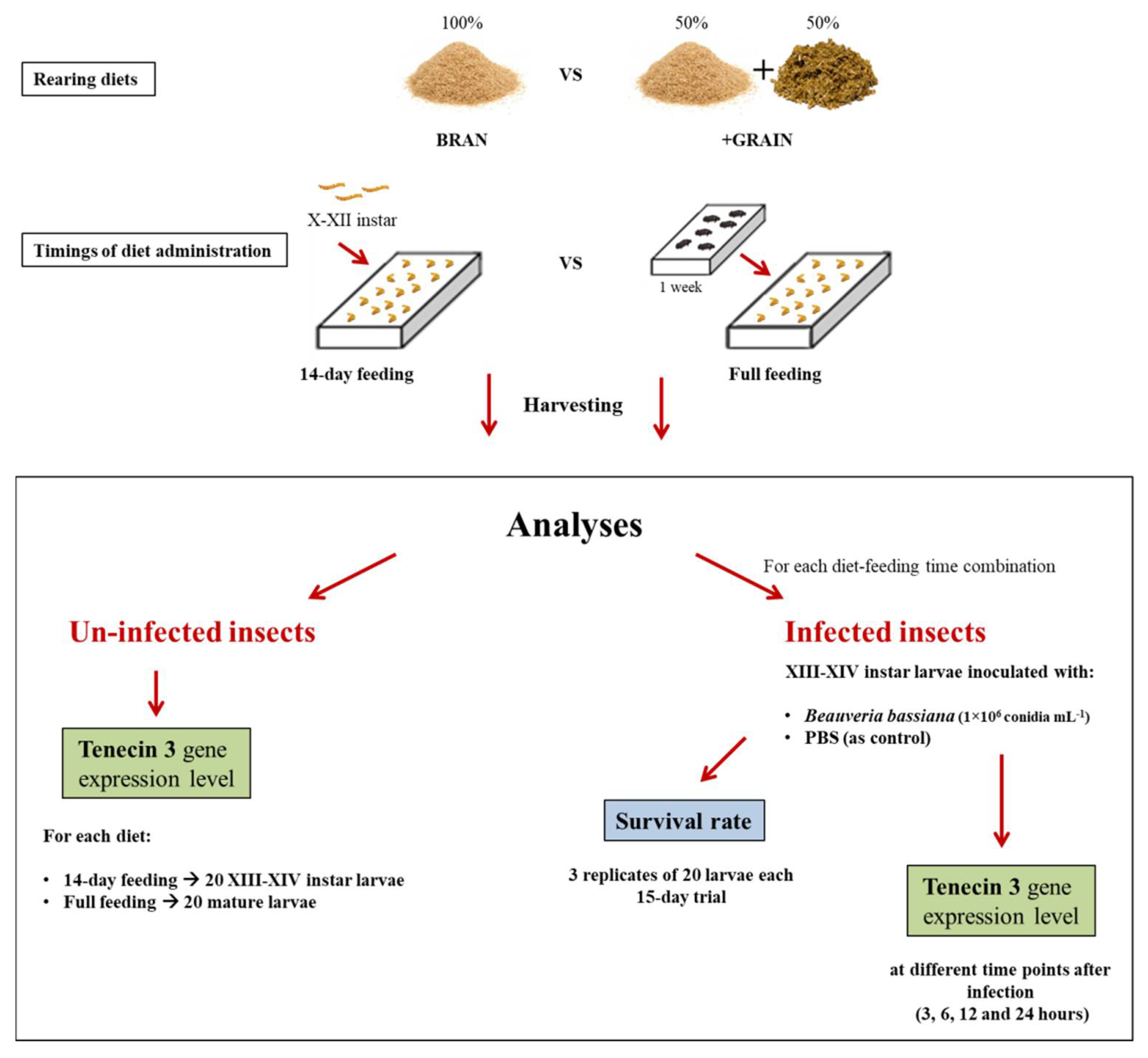
2.1. Diets and Feeding Times
2.2. Tenecin 3 Gene Expression Analysis in Uninfected Tenebrio Molitor Larvae
2.2.1. RNA Extraction
2.2.2. Quantitative Real-Time PCR
2.3. Tenebrio molitor Infection Assays with Beauveria bassiana
2.3.1. Inoculum Preparation
2.3.2. Survival Rate
2.3.3. Tenecin 3 Gene Expression Analysis in B. bassiana-Infected T. molitor Larvae
2.4. Statistical Analysis
3. Results
3.1. Tenebrio molitor Growing Performances
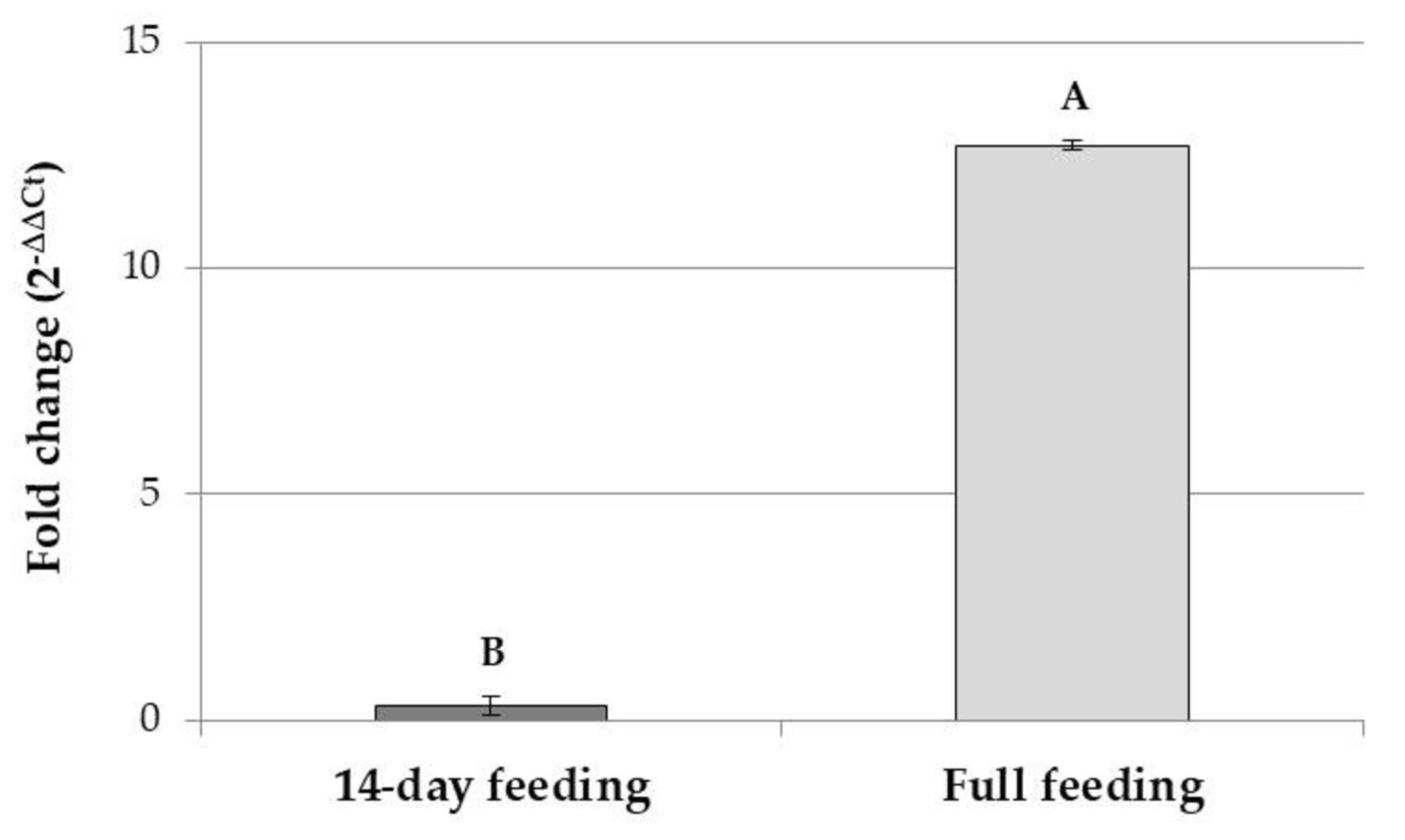
3.2. Tenecin 3 Gene Expression Analysis in Uninfected Tenebrio molitor Larvae
3.3. Tenebrio molitor Infection Assays with Beauveria bassiana
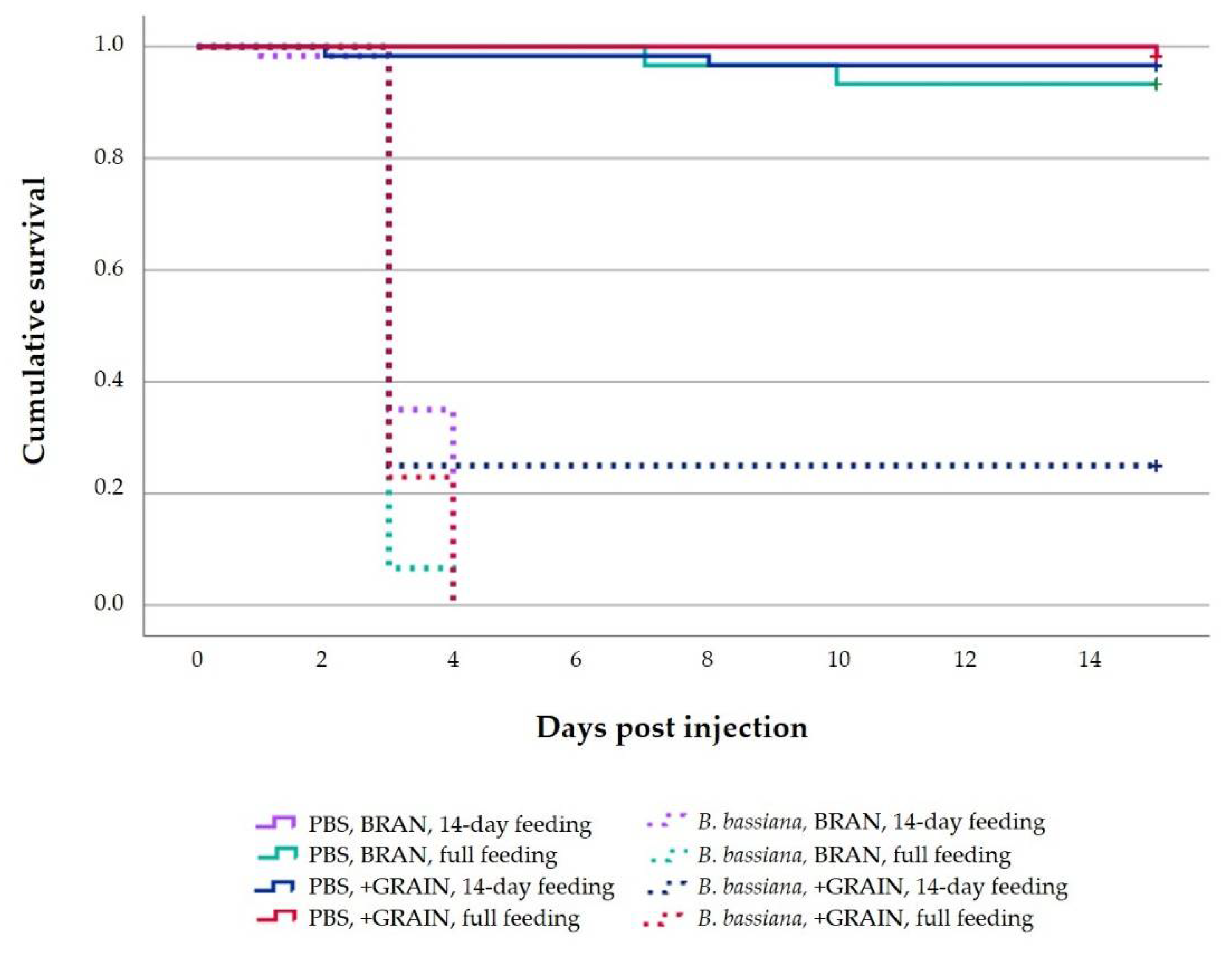
3.3.1. Survival Rate
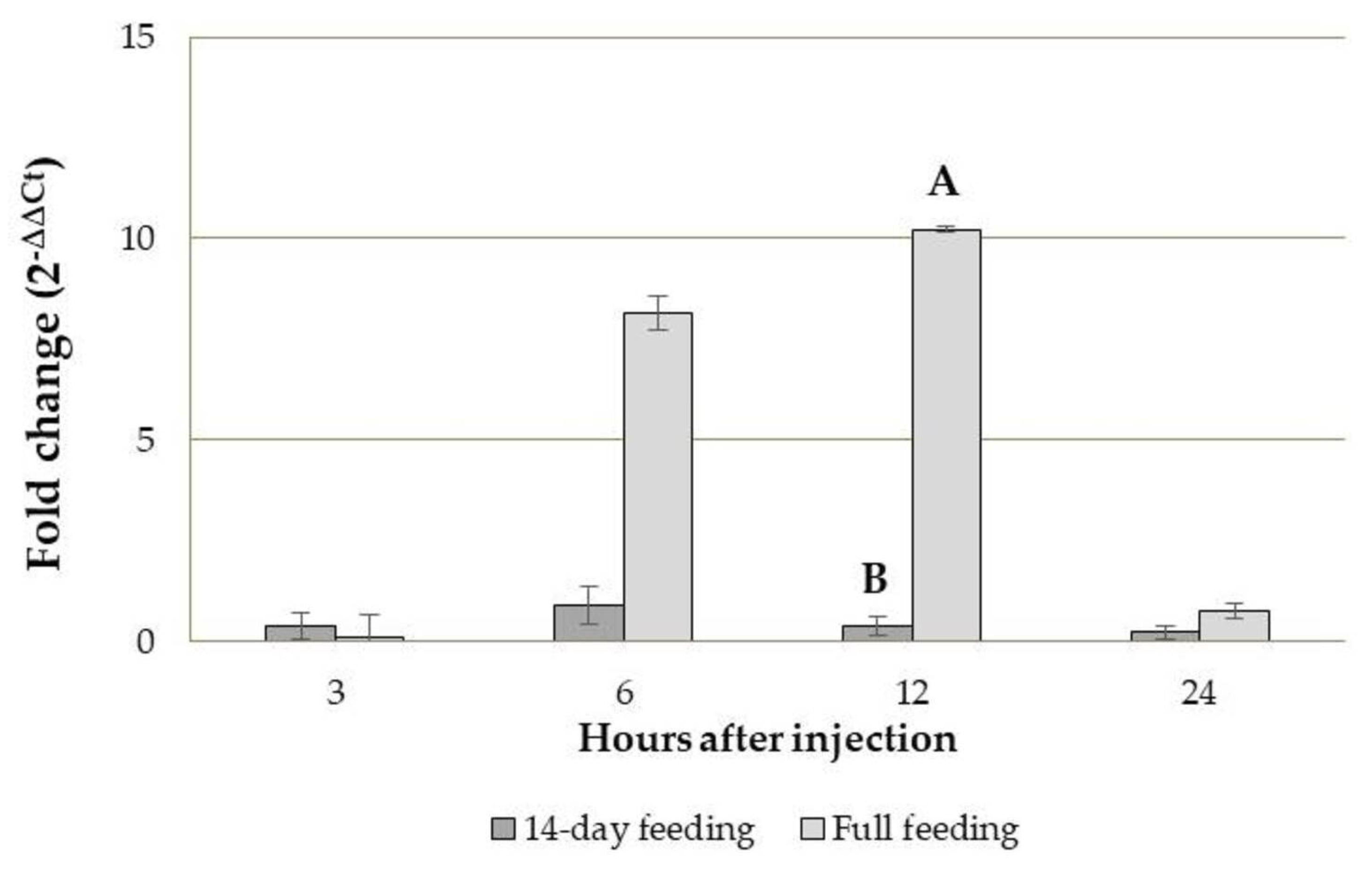
3.3.2. Tenecin 3 Gene Expression Analysis in B. bassiana-Infected T. molitor Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Huis, A. Prospects of insects as food and feed. Org. Agric. 2021, 11, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Bug Burger. Available online: https://www.bugburger.se/foretag/the-eating-insects-startups-here-is-the-list-of-entopreneurs-around-the-world/ (accessed on 27 February 2023).
- Barclays. I Can’t Believe It’s Not Meat. Available online: https://www.investmentbank.barclays.com/content/dam/barclaysmicrosites/ibpublic/documents/our-insights/FoodWaste/Leaflet%20Food%20Revolution.pdf (accessed on 27 February 2023).
- Rantala, M.J.; Viitaniemi, H.; Roff, D.A. Effects of inbreeding on potential and realized immune responses in Tenebrio molitor. Parasitology 2011, 138, 906–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. J. Insects Food Feed 2015, 1, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Maciel-Vergara, G.; Ros, V.I.D. Viruses of insects reared for food and feed. J. Invertebr. Pathol. 2017, 147, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J.; Van Oers, M.M.; Jensen, A.B.; Lecocq, A.; Maciel-Vergara, G.; Santacoloma, L.P.A.; Van Loon, J.J.A.; Hesketh, H. Towards a coordination of European activities to diagnose and manage insect diseases in production facilities. J. Insects Food Feed 2018, 4, 157–166. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.B.; Lecocq, A.; Eilenberg, J. Diseases in edible insect rearing systems. J. Insects Food Feed 2021, 7, 621–638. [Google Scholar] [CrossRef]
- Veldkamp, T.; Meijer, N.; Alleweldt, F.; Deruytter, D.; Van Campenhout, L.; Gasco, L.; Roos, N.; Smetana, S.; Fernandes, A.; van der Fels-Klerx, H.J. Overcoming technical and market barriers to enable sustainable large-scale production and consumption of insect proteins in Europe: A SUSINCHAIN Perspective. Insects 2022, 13, 281. [Google Scholar] [CrossRef]
- Tsakas, S.; Marmaras, V.J. Insect immunity and its signalling: An overview. Invertebr. Surviv. J. 2010, 7, 228–238. [Google Scholar]
- Wu, K.; Yang, B.; Huang, W.; Dobens, L.; Song, H.; Ling, E. Gut immunity in Lepidopteran insects. Dev. Comp. Immunol. 2016, 64, 65–74. [Google Scholar] [CrossRef]
- Strand, M.R. Insect hemocytes and their role in immunity. Insect Immunol. 2008, 32, 25–47. [Google Scholar]
- Gillespie, J.P.; Kanost, M.R.; Trenczek, T. Biological mediators of insect immunity. Annu. Rev. Entomol. 1997, 42, 611–643. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, O.; Theopold, U.; Strand, M. Innate immunity and its evasion and suppression by hymenopteran endoparasitoids. BioEssays 2001, 23, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Nappi, A.J.; Kohler, L.; Mastore, M. Signaling pathways implicated in the cellular innate immune responses of Drosophila. Invertebr. Surv. J. 2004, 1, 5–33. [Google Scholar]
- Lamprou, I.; Tsakas, S.; Theodorou, G.L.; Karakantza, M.; Lampropoulou, M.; Marmaras, V.J. Uptake of LPS/E. coli/latex beads via distinct signalling pathways in medfly hemocytes: The role of MAP kinases activation and protein secretion. Biochim. Biophys. Acta 2005, 1744, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mavrouli, M.D.; Tsakas, S.; Theodorou, G.L.; Lampropoulou, M.; Marmaras, V.J. MAP kinases mediate phagocytosis and melanization via prophenoloxidase activation in medfly hemocytes. Biochim. Biophys. Acta 2005, 1744, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Sideri, M.; Tsakas, S.; Markoutsa, E.; Lampropoulou, M.; Marmaras, V.J. Innate immunity in insects: Surface-associated dopa decarboxylase-dependent pathways regulate phagocytosis, nodulation and melanization in medfly haemocytes. Immunology 2008, 123, 528–537. [Google Scholar] [CrossRef]
- Vigneron, A.; Jehan, C.; Rigaud, T.; Moret, Y. Immune defenses of a beneficial pest: The mealworm beetle, Tenebrio molitor. Front. Physiol. 2019, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Hillyer, J.F.; Strand, M.R. Mosquito hemocyte-mediated immune responses. Curr. Opin. Insect Sci. 2014, 3, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Bracho, C. Epidemiology of Chagas disease in Mexico: An update. Trends Parasitol. 2001, 17, 372–376. [Google Scholar] [CrossRef]
- Noireau, F.; Carbajal-De-La-Fuente, A.L.; Lopes, C.M.; Diotaiuti, L. Some considerations about the ecology of Triatominae. An. Acad. Bras. Cienc. 2005, 77, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Bello-Bedoy, R.; Peiro-Nuño, H.; Córdoba-Aguilar, A.; Flores-López, C.A.; Romero-Figueroa, G.; Arteaga, M.C.; Gutiérrez-Cabrera, A.E.; De la Rosa-Conroy, L. Ontogenetic changes in wild chagasic bugs (Dipetalogaster maximus): Exploring morphological adaptations in pre-adult and adult stages. Rev. Mex. Biodivers. 2019, 90, e902664. [Google Scholar] [CrossRef]
- Manniello, M.D.; Moretta, A.; Salvia, R.; Scieuzo, C.; Lucchetti, D.; Vogel, H.; Sgambato, A.; Falabella, P. Insect antimicrobial peptides: Potential weapons to counteract the antibiotic resistance. Cell. Mol. Life Sci. 2021, 78, 4259–4282. [Google Scholar] [CrossRef]
- Wilson, K.; Reeson, A.F. Density-dependent prophylaxis: Evidence from Lepidoptera-baculovirus interactions? Ecol. Entomol. 1998, 23, 100–101. [Google Scholar] [CrossRef] [Green Version]
- Prokkola, J.; Roff, D.; Karkkainen, T.; Krams, I.; Rantala, M.J. Genetic and phenotypic relationships between immune defense, melanism and life-history traits at different temperatures and sexes in Tenebrio molitor. Heredity 2013, 111, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.W.S.; Elliot, S.L. Temperature and population density: Interactional effects of environmental factors on phenotypic plasticity, immune defenses, and disease resistance in an insect pest. Ecol. Evol. 2016, 6, 3672–3683. [Google Scholar] [CrossRef] [Green Version]
- Laughton, A.M.; O’connor, C.O.; Knell, R.J. Responses to a warming world: Integrating life history, immune investment, and pathogen resistance in a model insect species. Ecol. Evol. 2017, 7, 9699–9710. [Google Scholar] [CrossRef]
- Cammack, J.; Tomberlin, J. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Catalán, T.P.; Barceló, M.; Niemeyer, H.M.; Kalergis, A.M.; Bozinovic, F. Pathogen-and diet-dependent foraging, nutritional and immune ecology in mealworms. Evol. Ecol. Res. 2011, 13, 711–723. [Google Scholar]
- Siva-Jothy, M.T.; Thompson, J.J. Short term nutrient deprivation affects immune function. Physiol. Entomol. 2002, 27, 206–212. [Google Scholar] [CrossRef]
- De Smet, J.; Wynants, E.; Cos, P.; Van Campenhout, L. Microbial community dynamics during rearing of black soldier fly larvae (Hermetia illucens) and impact on exploitation potential. Appl. Environ. Microbiol. 2018, 84, e02722-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krams, I.; Kecko, S.; Kangassalo, K.; Moore, F.R.; Jankevics, E.; Inashkina, I.; Krama, T.; Lietuvietis, V.; Meija, L.; Rantala, M.J. Effects of food quality on trade-offs among growth, immunity and survival in the greater wax moth Galleria mellonella. Insect Sci. 2015, 22, 431–439. [Google Scholar] [CrossRef]
- Bruno, D.; Bonelli, M.; De Filippis, F.; Di Lelio, I.; Tettamanti, G.; Casartelli, M.; Ercolini, D.; Caccia, S. The intestinal microbiota of Hermetia illucens larvae is affected by diet and shows a diverse composition in the different midgut regions. Appl. Environ. Microbiol. 2019, 85, e01864-18. [Google Scholar] [CrossRef] [Green Version]
- Candian, V.; Savio, C.; Meneguz, M.; Gasco, L.; Tedeschi, R. Effect of the rearing diet on gene expression of antimicrobial peptides in Hermetia illucens (Diptera: Stratiomyidae). Insect Sci. 2022. [Google Scholar] [CrossRef]
- Candian, V.; Meneguz, M.; Tedeschi, R. Immune responses of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae) reared on catering waste. Life 2023, 13, 213. [Google Scholar] [CrossRef]
- Smits, S.A.; Leach, J.; Sonnenburg, E.D.; Gonzalez, C.G.; Lichtman, J.S.; Reid, G.; Knight, R.; Manjurano, A.; Changalucha, J.; Elias, J.E.; et al. Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science 2017, 357, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.; Müller, A.; Heckel, D.G.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef]
- Chae, J.H.; Kurokawa, K.; So, Y.I.; Hwang, H.O.; Kim, M.S.; Park, J.W. Purification and characterization of Tenecin 4, a new anti Gram-negative bacterial peptide, from the beetle Tenebrio molitor. Dev. Comp. Immunol. 2012, 36, 540–546. [Google Scholar] [CrossRef]
- Keshavarz, M.; Jo, Y.H.; Park, K.B.; Ko, H.J.; Edosa, T.T.; Lee, Y.S.; Han, Y.S. TmDorX2 positively regulates antimicrobial peptides in Tenebrio molitor gut, fat body, and hemocytes in response to bacterial and fungal infection. Sci. Rep. 2019, 9, 16878. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Wu, G.X.; Zhang, Z. Upregulation of coleoptericin transcription in Tenebrio molitor parasitized by Scleroderma guani. J. Asia Pac. Entomol. 2014, 17, 339–342. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- El Deen, S.N.; Lamaj, F. Productivity and larval growth of Tenebrio molitor reared on differently composed diets of similar nutritional composition. J. Insects Food Feed 2021, 7, 1207–1217. [Google Scholar] [CrossRef]
- Melis, R.; Braca, A.; Sanna, R.; Spada, S.; Mulas, G.; Fadda, M.L.; Sassu, M.M.; Serra, G.; Anedda, R. Metabolic response of yellow mealworm larvae to two alternative rearing substrates. Metabolomics 2019, 15, 113. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Chen, G.; Qiao, L.; Li, J.; Liu, B.; Liu, Z.; Li, M.; Liu, X. Growth performance and nutritional profile of mealworms reared on corn stover, soybean meal, and distillers’ grains. Eur. Food Res. Technol. 2019, 245, 2631–2640. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth potential of yellow mealworm reared on industrial residues. Agriculture 2020, 10, 599. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, S.; Paci, G.; Fratini, F.; Dal Bosco, A.; Tuccinardi, T.; Mancini, S. Former foodstuff in mealworm farming: Effects on fatty acids profile, lipid metabolism and antioxidant molecules. LWT 2021, 147, 111644. [Google Scholar] [CrossRef]
- Montalbán, A.; Sánchez, C.J.; Hernández, F.; Schiavone, A.; Madrid, J.; Martınez-Miró, S. Effects of agro-industrial byproduct-based diets on the growth performance, digestibility, nutritional and microbiota composition of mealworm (Tenebrio molitor L.). Insects 2022, 13, 323. [Google Scholar] [CrossRef]
- Xiros, C.; Christakopoulos, P. Biotechnological potential of brewers spent grain and its recent applications. Waste Biomass Valoriz. 2012, 3, 213–232. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, H.G.; Lee, K.Y.; Yoon, H.J.; Kim, N.J. Effects of Brewer’s spent grain (BSG) on larval growth of mealworms, Tenebrio molitor (Coleoptera: Tenebrionidae). Int. J. Indust. Entomol. 2016, 32, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Jucker, C.; Leonardi, M.G.; Rigamonti, I.; Lupi, D.; Savoldelli, S. Brewery’s waste streams as a valuable substrate for black soldier fly Hermetia illucens (Diptera: Stratiomyidae). J. Entomol. Acarol. Res. 2019, 51, 8876. [Google Scholar] [CrossRef]
- Wilson, J.K.; Ruiz, L.; Davidowitz, G. Dietary protein and carbohydrates affect immune function and performance in a specialist herbivore insect (Manduca sexta). Physiol. Biochem. Zool. 2019, 92, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Abisgold, J.D.; Simpson, S.J. The physiology of compensation by locusts for changes in dietary protein. J. Exp. Biol. 1987, 129, 329–346. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Maistrou, S.; Paris, V.; Jensen, A.B.; Rolff, J.; Meyling, N.V.; Zanchi, C. A constitutively expressed antifungal peptide protects Tenebrio molitor during a natural infection by the entomopathogenic fungus Beauveria bassiana. Dev. Comp. Immunol. 2018, 86, 26–33. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Baldwin, H.; Ffrench-Constant, R.H.; Reynolds, S.E. Developmental modulation of immunity: Changes within the feeding period of the fifth larval stage in the defence reactions of Manduca sexta to infection by Photorhabdus. J. Insect Physiol. 2008, 54, 309–318. [Google Scholar] [CrossRef]
- Tian, L.; Guo, E.E.; Diao, Y.P.; Zhou, S.; Peng, Q.; Cao, Y.; Ling, E.J.; Li, S. Genome-wide regulation of innate immunity by juvenile hormone and 20-hydroxyecdysone in the Bombyx fat body. BMC Genom. 2010, 11, 549. [Google Scholar] [CrossRef] [Green Version]
- Booth, K.; Cambron, L.; Fisher, N.; Greenlee, K.J. Immune defense varies within an instar in the tobacco hornworm, Manduca sexta. Physiol. Biochem. Zool. 2015, 88, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Benesova, J.; Dobes, P.; Hyrsl, P. Developmental changes in phenol-oxidizing activity in the greater wax moth Galleria mellonella. Bull. Insectol. 2009, 62, 237–243. [Google Scholar]
- Lu, J.-F.; Hu, J.; Fu, W.-J. Levels of encapsulation and melanization in two larval instars of Ostrinia furnacalis Guenee (Lep., Pyralidae) during simulation of parasitization by Macrocentrus cingulum Brischke (Hym., Braconidae). J. Appl. Entomol. 2006, 130, 290–296. [Google Scholar] [CrossRef]
- Woods, H.A. Patterns and mechanisms of growth of fifth-instar Manduca sexta caterpillars following exposure to low-or high-protein food during early instars. Physiol. Biochem. Zool. 1999, 72, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Pendland, J.C.; Hung, S.Y.; Boucias, D.G. Evasion of host defence by in vivo-produced protoplast-like cellsof the insectmycopathogen Beauveria bassiana. J. Bacteriol. 1993, 175, 5962–5969. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.; Park, S.; Choi, J.; Jeong, G.; Lee, S.B.; Choi, Y.; Lee, S.J. The intestinal bacterial community in the food waste-reducing larvae of Hermetia illucens. Curr. Microbiol. 2011, 62, 1390–1399. [Google Scholar] [CrossRef]
- Jang, S.; Kikuchi, Y. Impact of the insect gut microbiota on ecology, evolution, and industry. Curr. Opin. Insect Sci. 2020, 41, 33–39. [Google Scholar] [CrossRef]
- Lu, H.L.; Leger, R.S. Insect immunity to entomopathogenic fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar] [CrossRef]
- Yang, Y.T.; Lee, M.R.; Lee, S.J.; Kim, S.; Nai, Y.S.; Kim, J.S. Tenebrio molitor Gram-negative-binding protein 3 (TmGNBP3) is essential for inducing downstream antifungal Tenecin 1 gene expression against infection with Beauveria bassiana JEF-007. Insect Sci. 2017, 25, 969–977. [Google Scholar] [CrossRef]
- Haine, E.R.; Pollitt, L.C.; Moret, Y.; Siva-Jothy, M.T.; Rolff, J. Temporal patterns in immune responses to a range of microbial insults (Tenebrio molitor). J. Insect Physiol. 2008, 54, 1090–1097. [Google Scholar] [CrossRef]
- Lemaitre, B.; Reichhart, J.M.; Hoffmann, J.A. Drosophila host defense: Differential induction of antimicrobial peptide genes after infection by various classes of microorganisms. Proc. Natl. Acad. Sci. USA 1997, 94, 14614–14619. [Google Scholar] [CrossRef] [Green Version]
- Korner, P.; Schmid-Hempel, P. In vivo dynamics of an immune response in the bumble bee Bombus terrestris. J. Invertebr. Pathol. 2004, 87, 59–66. [Google Scholar] [CrossRef]
- Lavine, M.D.; Chen, G.; Strand, M.R. Immune challenge differentially affects transcript abundance of three antimicrobial peptides in hemocytes from the moth Pseudoplusia includens. Insect Biochem. Mol. Biol. 2005, 35, 1335–1346. [Google Scholar] [CrossRef]
- Moret, Y.; Siva-Jothy, M.T. Adaptive innate immunity? Responsive-mode prophylaxis in the mealworm beetle, Tenebrio molitor. Proc. R. Soc. Lond. B 2003, 273, 251–256. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candian, V.; Tedeschi, R. Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio molitor Larvae Infected by Beauveria bassiana. Insects 2023, 14, 359. https://doi.org/10.3390/insects14040359
Candian V, Tedeschi R. Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio molitor Larvae Infected by Beauveria bassiana. Insects. 2023; 14(4):359. https://doi.org/10.3390/insects14040359
Chicago/Turabian StyleCandian, Valentina, and Rosemarie Tedeschi. 2023. "Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio molitor Larvae Infected by Beauveria bassiana" Insects 14, no. 4: 359. https://doi.org/10.3390/insects14040359