
Geostatistical Analysis of the Spatial Variation of Chrysolina aeruginosa Larvae at Different Stages in Desert Ecosystems


Abstract
:Simple Summary
Abstract
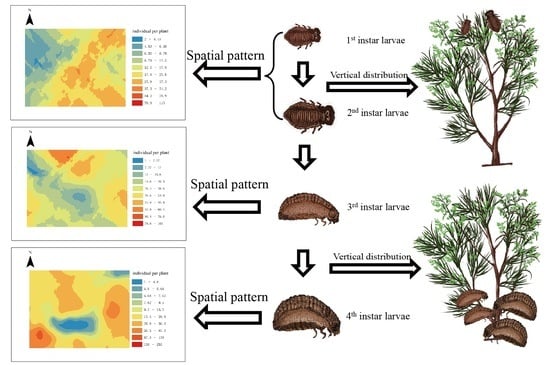
1. Introduction
2. Materials and Methods
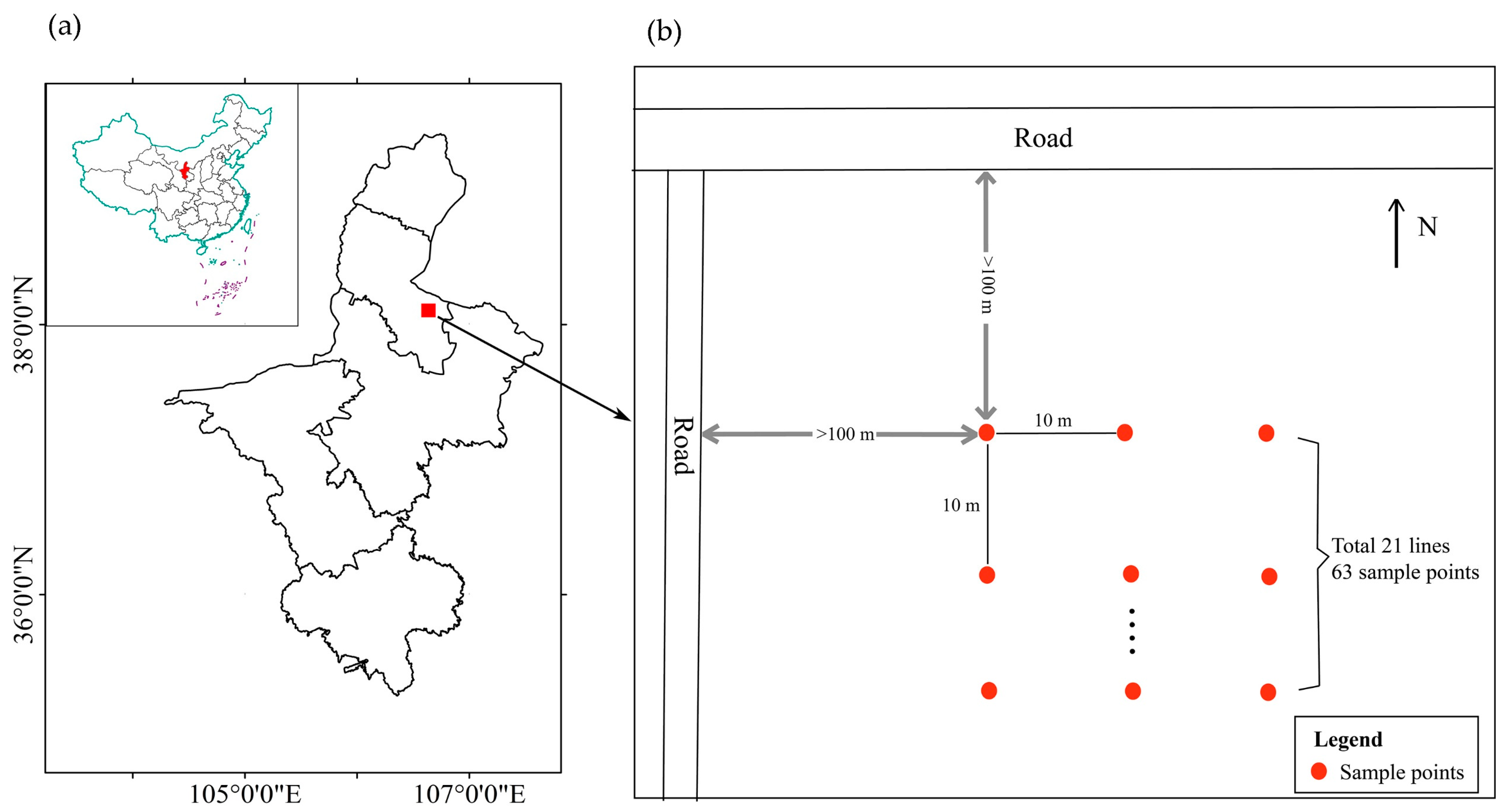
2.1. Study Area and Study Species
2.2. Experimental Procedures
2.2.1. Population Survey of Different Instar Larvae
2.2.2. Vertical Distribution Survey of Different Instar Larvae
2.2.3. Geostatistical Analysis
2.3. Statistical Analyses
3. Results
3.1. Basic Information of C. aeruginosa Larvae
3.2. The Vertical Distribution of C. aeruginosa Larvae
3.3. Correlation Analysis of the Number of C. aeruginosa Larvae and A. ordosica Characteristics
3.4. Geostatistical Analysis of C. aeruginosa Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rijal, J.P.; Brewster, C.C.; Bergh, J.C. Spatial Distribution of Grape Root Borer (Lepidoptera: Sesiidae) Infestations in Virginia Vineyards and Implications for Sampling. Environ. Entomol. 2014, 43, 716–728. [Google Scholar] [CrossRef] [PubMed]
- Maestre-Serrano, R.; Flórez-Rivadeneira, Z.; Castro-Camacho, J.M.; Soto-Arenilla, E.; Gómez-Camargo, D.; Pareja-Loaiza, P.; Ponce-Garcia, G.; Juache-Villagrana, A.E.; Flores, A.E. Spatial Distribution of Pyrethroid Resistance and Kdr Mutations in Aedes aegypti from La Guajira, Colombia. Insects 2023, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Ifoulis, A.A.; Savopoulou-Soultani, M. Use of Geostatistical Analysis to Characterize the Spatial Distribution of Lobesia botrana (Lepidoptera: Tortricidae) Larvae in Northern Greece. Environ. Entomol. 2006, 35, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.C.; Picanço, M.C.; Silva, R.S.; Gonring, A.H.; Galdino, T.V.; Guedes, R.N. Assessing the Spatial Distribution of Tuta absoluta (Lepidoptera: Gelechiidae) Eggs in Open-Field Tomato Cultivation through Geostatistical Analysis. Pest Manag. Sci. 2018, 74, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.S.; Sarmento, R.A.; Lima, C.H.O.; Pinto, C.B.; Silva, G.A.; Dos Santos, G.R.; Picanço, M.C. Geostatistical Assessment of Frankliniella schultzei (Thysanoptera: Thripidae) Spatial Distribution in Commercial Watermelon Crops. J. Econ. Entomol. 2020, 113, 489–495. [Google Scholar] [CrossRef]
- Diaz, B.M.; Barrios, L.; Fereres, A. Interplant Movement and Spatial Distribution of Alate and Apterous Morphs of Nasonovia ribisnigri (Homoptera: Aphididae) on Lettuce. Bull. Entomol. Res. 2012, 102, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Mahla, M.; Swaminathan, R.; Babu, S.; Kumar, A.; Ahir, K.; Singh, B.; Chhangani, G. Assessment of Spatial Distribution of Plutella xylostella on Cabbage (Brassica Oleracea Var. Capitata). Indian J. Agric. Sci. 2022, 92, 190–194. [Google Scholar] [CrossRef]
- Matheron, G. Principles of Geostatistics. Econ. Geol. 1963, 58, 1246–1266. [Google Scholar] [CrossRef]
- Foresti, J.; Pereira, R.R.; Santana Jr, P.A.; das Neves, T.N.; da Silva, P.R.; Rosseto, J.; Novais Istchuk, A.; Ishizuka, T.K.; Harter, W.; Schwertner, M.H.; et al. Spatial–Temporal Distribution of Dalbulus maidis (Hemiptera: Cicadellidae) and Factors Affecting Its Abundance in Brazil Corn. Pest Manag. Sci. 2022, 78, 2196–2203. [Google Scholar] [CrossRef]
- Cocco, A.; Serra, G.; Lentini, A.; Deliperi, S.; Delrio, G. Spatial Distribution and Sequential Sampling Plans for Tuta absoluta (Lepidoptera: Gelechiidae) in Greenhouse Tomato Crops. Pest Manag. Sci. 2015, 71, 1311–1323. [Google Scholar] [CrossRef]
- Saleem, M.J.; Hafeez, F.; Arshad, M.; Atta, B.; Maan, N.A.; Ayub, M.A.; Zubair, M. Population Dynamics of Sucking Pests on Transgenic Bt Cotton in Relation with Abiotic Factors and Physio-Morphological Plant Characters. J. Entomol. Zool. Stud. 2018, 6, 163–166. [Google Scholar]
- Shi, J.; Luo, Y.-Q.; Song, J.-Y.; Wu, H.-W.; Wang, L.; Wang, G.Z. Traits of Masson Pine Affecting Attack of Pine Wood Nematode. J. Integr. Plant Biol. 2007, 49, 1763–1771. [Google Scholar] [CrossRef]
- Haysom, K.A.; Coulson, J.C. The Lepidoptera Fauna Associated with Calluna Vulgaris: Effects of Plant Architecture on Abundance and Diversity. Ecol. Entomol. 1998, 23, 377–385. [Google Scholar] [CrossRef]
- Contarini, M.; Rossini, L.; Di Sora, N.; de Lillo, E.; Speranza, S. Monitoring the Bud Mite Pest in a Hazelnut Orchard of Central Italy: Do Plant Height and Irrigation Influence the Infestation Level? Agronomy 2022, 12, 1982. [Google Scholar] [CrossRef]
- Yan, W.; Luo, Y.; Zong, S.; Bao, S.; Sun, Y.; Li, Y. Woodborers Abundance and the Relationship with Environmental Factors at Different Successional Stages of Artemisia ordosica (Asterales: Compositae). Sci. Silvae Sin. 2009, 45, 87–91. (In Chinese) [Google Scholar]
- Yan, W.; Zong, S.; Luo, Y.; Cao, C.; Li, Z.; Guo, Q. Application of Stepwise Regression Model in Predicting the Movement of Artemisia ordosica Boring Insects. J. Beijing For. Univ. 2009, 31, 140–144. (In Chinese) [Google Scholar]
- Stoeckli, S.; Mody, K.; Dorn, S. Influence of Canopy Aspect and Height on Codling Moth (Lepidoptera: Tortricidae) Larval Infestation in Apple, and Relationship between Infestation and Fruit Size. J. Econ. Entomol. 2008, 101, 81–89. [Google Scholar] [CrossRef]
- Pekár, S. Horizontal and Vertical Distribution of Spiders (Araneae) in Sunflowers. J. Arachnol. 2005, 33, 197–204. [Google Scholar] [CrossRef]
- Wei, S.-H.; Zhu, M.-M.; Zhang, R.; Huang, W.-G.; Yu, Z. Effects of temperature on the development and reproduction of Chrysolina aeruginosa (Coleoptera: Chrysomelidae). Acta Entomol. Sin. 2013, 56, 1004–1009. (In Chinese) [Google Scholar]
- Wei, S.-H.; Zhu, M.-M.; Zhang, R.; Huang, W.-G.; Yu, Z. Morphology and Bionomics of Chrysolina aeruginosa Fald. Ningxia J. Agric. For. 2013, 4, 58–59. (In Chinese) [Google Scholar]
- Cui, Y.-Q.; Luo, Y.-Q.; Bai, L.; Zong, S.-X. Geostatistical Analysis of the Spatial Distribution of Three Boring Pests of Artemisia ordosica (Lepidoptera: Cossidae/Coleoptera: Buprestidae, Curculionidae). Entomol. Gen. 2013, 34, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wei, J.; Zhang, K.; Li, H.; Wei, S.; Pan, X.; Huang, W.; Zhu, M.; Zhang, R. Asymmetric Response of Different Functional Insect Groups to Low-Grazing Pressure in Eurasian Steppe in Ningxia. Ecol. Evol. 2018, 8, 11609–11618. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.L.; Kabir, M.H.; Nila, M.U.S.; Rubaiyat, A. Response of Grassland Net Primary Productivity to Dry and Wet Climatic Events in Four Grassland Types in Inner Mongolia. Plant-Environ. Interact. 2021, 2, 250–262. [Google Scholar] [CrossRef]
- Li, X.; Yu, M.-H.; Ding, G.-D.; He, Y.; Liu, W.; Wang, C.-Y. Soil Biocrusts Reduce Seed Germination and Contribute to the Decline in Artemisia ordosica Krasch. Shrub Populations in the Mu Us Sandy Land of North China. Glob. Ecol. Conserv. 2021, 26, e01467. [Google Scholar] [CrossRef]
- Zong, S.; Luo, Y.; Cui, Y.; Wang, J.; Yan, W.; Liu, A.; Kari, H. Damage Characteristics of Three Boring Pests in Artemisia ordosica. For. Stud. China 2009, 11, 24–27. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, L.-L.; Luo, Y.-Q.; Zong, S.-X. Scanning Electron Microscopy Analysis of the Cephalic Sensilla of Chrysolina aeruginosa Fald. (Coleoptera, Chrysomelidae). Microsc. Res. Tech. 2013, 76, 423–431. [Google Scholar] [CrossRef]
- Tian, C.; He, D.-H.; Li, Y.-J. The Occurrence and Control of Desert Forage Grass Insect Pests, Chrysolina aeruginosa (Falder). Plant Prot. China 1987, 13, 25–26. (In Chinese) [Google Scholar]
- Zhang, Z.-K.; Yang, C.-X.; Gao, L.-Y. Study on the Spatial Distribution Pattern of Chrysolina aeruginosa Fald. and its Sampling Technique. J. Northwest AF Univ. (Nat. Sci. Ed) 2007, 35, 99–104. (In Chinese) [Google Scholar]
- Duarte, F.; Calvo, M.; Borges, A.; Scatoni, I. Geostatistics Applied to the Study of the Spatial Distribution of Insects and Its Use in Integrated Pest Management. Rev. Agron. Noroeste Argent. 2015, 35, 9–20. [Google Scholar]
- Yang, X.; Qin, J.; Luo, Y.; Yang, Z.; Wei, J. Geostatistical Analysis of Spatial Distribution of Endoclita signifer Larvae on Eucalyptus. Am. J. Agric. For. 2018, 6, 226–236. [Google Scholar]
- Ribeiro, A.V.; Ramos, R.S.; de Araújo, T.A.; Soares, J.R.; Paes, J.d.S.; de Araújo, V.C.; Bastos, C.S.; Koch, R.L.; Picanço, M.C. Spatial Distribution and Colonization Pattern of Bemisia tabaci in Tropical Tomato Crops. Pest Manag. Sci. 2021, 77, 2087–2096. [Google Scholar] [CrossRef]
- Dangol, D.; Khanal, L.; Pandey, N.; Ghimire, A.; Kyes, R.C. Test of Ecogeographical Rules on Sparrows (Passer spp.) along the Elevation Gradient of the Himalaya in Central Nepal. Ecologies 2022, 3, 480–491. [Google Scholar] [CrossRef]
- Cao, S.-G.; Pang, Z.-H.; Yang, X.-H.; Yu, Y.-H.; Qiu, R.-Q. Preliminary Study on Spatial Distribution Pattern of Endoclyta signifer Walker Larva. Plant Dis. Pests 2011, 2, 28–72. [Google Scholar]
- Karimzadeh, R.; Hejazi, M.J.; Helali, H.; Iranipour, S.; Mohammadi, S.A. Analysis of the Spatio-Temporal Distribution of Eurygaster integriceps (Hemiptera: Scutelleridae) by Using Spatial Analysis by Distance Indices and Geostatistics. Environ. Entomol. 2011, 40, 1253–1265. [Google Scholar] [CrossRef]
- de Alves, M.C.; da Silva, F.M.; Moraes, J.C.; Pozza, E.A.; de Oliveira, M.S.; Souza, J.C.S.; Alves, L.S. Geostatistical Analysis of the Spatial Variation of the Berry Borer and Leaf Miner in a Coffee Agroecosystem. Precis. Agric. 2011, 12, 18–31. [Google Scholar] [CrossRef]
- Negrón, J.F. Within-Stand Distribution of Tree Mortality Caused by Mountain Pine Beetle, Dendroctonus ponderosae Hopkins. Insects 2020, 11, 112. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wu, J.; Shang, H.; Cheng, J. Landscape Connectivity Shapes the Spread Pattern of the Rice Water Weevil: A Case Study from Zhejiang, China. Environ. Manag. 2011, 47, 254–262. [Google Scholar] [CrossRef]
- Hortal, J.; Roura-Pascual, N.; Sanders, N.J.; Rahbek, C. Understanding (Insect) Species Distributions across Spatial Scales. Ecography 2010, 33, 51–53. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.-A.; Guisan, A.; et al. The Role of Biotic Interactions in Shaping Distributions and Realised Assemblages of Species: Implications for Species Distribution Modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Macfadyen, S.; Tay, W.T.; Hulthen, A.D.; Paull, C.; Kalyebi, A.; Jacomb, F.; Parry, H.; Sseruwagi, P.; Seguni, Z.; Omongo, C.A.; et al. Landscape Factors and How They Influence Whitefly Pests in Cassava Fields across East Africa. Landsc. Ecol. 2021, 36, 45–67. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; De Simone, W.; Biondi, M. Habitat Specificity, Host Plants and Areas of Endemism for the Genera-Group Blepharida s.l. in the Afrotropical Region (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Insects 2021, 12, 299. [Google Scholar] [CrossRef] [PubMed]
- Steffan-Dewenter, I.; Tscharntke, T. Insect Communities and Biotic Interactions on Fragmented Calcareous Grasslands—A Mini Review. Biol. Conserv. 2002, 104, 275–284. [Google Scholar] [CrossRef]
- Garcia, A.G.; Araujo, M.R.; Uramoto, K.; Walder, J.M.M.; Zucchi, R.A. Geostatistics and Geographic Information System to Analyze the Spatial Distribution of the Diversity of Anastrepha Species (Diptera: Tephritidae): The Effect of Forest Fragments in an Urban Area. Environ. Entomol. 2017, 46, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-Z.; Zhao, N.; Li, Y.-C.; Ma, Y.; Wang, X.; Xie, J.-C.; Ma, C.-L.; Chen, H.-B. Population dynamics of Chrysolina aeruginosa in Ningdong region of Ningxia and the related influencing factors. Chin. J. Ecol. 2014, 33, 346–351. (In Chinese) [Google Scholar]
- Vinatier, F.; Tixier, P.; Duyck, P.-F.; Lescourret, F. Factors and Mechanisms Explaining Spatial Heterogeneity: A Review of Methods for Insect Populations. Methods Ecol. Evol. 2011, 2, 11–22. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Bergeron, J.A.C.; Spence, J.R.; He, F. Landscape Effects of Disturbance, Habitat Heterogeneity and Spatial Autocorrelation for a Ground Beetle (Carabidae) Assemblage in Mature Boreal Forest. Ecography 2013, 36, 636–647. [Google Scholar] [CrossRef]
- Karimzadeh, R.; Iranipour, S. Spatial Distribution and Site-Specific Spraying of Main Sucking Pests of Elm Trees. Neotrop. Entomol. 2017, 46, 316–323. [Google Scholar] [CrossRef]
- Mitchell, M.G.E.; Hartley, E.; Tsuruda, M.; Gonzalez, A.; Bennett, E.M. Contrasting Responses of Soybean Aphids, Primary Parasitoids, and Hyperparasitoids to Forest Fragments and Agricultural Landscape Structure. Agric. Ecosyst. Environ. 2022, 326, 107752. [Google Scholar] [CrossRef]
- Gallé, R.; Tölgyesi, C.; Császár, P.; Bátori, Z.; Gallé-Szpisjak, N.; Kaur, H.; Maák, I.; Torma, A.; Batáry, P. Landscape Structure Is a Major Driver of Plant and Arthropod Diversity in Natural European Forest Fragments. Ecosphere 2022, 13, e3905. [Google Scholar] [CrossRef]
- Sciarretta, A.; Trematerra, P. Geostatistical Tools for the Study of Insect Spatial Distribution: Practical Implications in the Integrated Management of Orchard and Vineyard Pests. Plant Prot. Sci. 2014, 50, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.E.; Prevedello, J.A.; Delciellos, A.C.; Vieira, M.V. Edge Effects and Geometric Constraints: A Landscape-Level Empirical Test. J. Anim. Ecol. 2016, 85, 97–105. [Google Scholar] [CrossRef]
- Altamirano, A.; Valladares, G.; Kuzmanich, N.; Salvo, A. Galling Insects in a Fragmented Forest: Incidence of Habitat Loss, Edge Effects and Plant Availability. J. Insect Conserv. 2016, 20, 119–127. [Google Scholar] [CrossRef]
- Mulcahy, M.M.; Wilson, B.E.; Reagan, T.E. Spatial Distribution of Lepidopteran Stem Borers in Louisiana Rice Fields. Environ. Entomol. 2022, 51, 405–412. [Google Scholar] [CrossRef]
- Mulcahy, M.M.; Wilson, B.E.; Reagan, T.E. Spatial Distribution of Lissorhoptrus oryzophilus (Coleoptera: Curculionidae) in Rice. Environ. Entomol. 2022, 51, 108–117. [Google Scholar] [CrossRef]
- Ferguson, A.W.; Klukowski, Z.; Walczak, B.; Clark, S.J.; Mugglestone, M.A.; Perry, J.N.; Williams, I.H. Spatial Distribution of Pest Insects in Oilseed Rape: Implications for Integrated Pest Management. Agric. Ecosyst. Environ. 2003, 95, 509–521. [Google Scholar] [CrossRef]
- Villaseñor, N.R.; Driscoll, D.A.; Escobar, M.A.; Gibbons, P.; Lindenmayer, D.B. Urbanization Impacts on Mammals Across Urban-Forest Edges and a Predictive Model of Edge Effects. PLoS ONE 2014, 9, e97036. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, Y.-C.; Zhang, D.-Z. Population dispersion of Chrysolina aeruginosa based on mark-recapture method. J. Environ. Entomol. 2016, 38, 912–917. (In Chinese) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | df | Chi-Square (χ2) | Estimate ± Standard Error | p |
|---|---|---|---|---|
| Intercept | 1 | 152.847 | 1.124 ± 0.090 | <0.0001 |
| Distribution height | 1 | 9.715 | 0.002 ± 0.001 | 0.0018 |
| Plants height | 1 | 12.647 | 0.004 ± 0.001 | 0.0004 |
| Crown width | 1 | 20.243 | 0.003 ± 0.001 | <0.0001 |
| Ground diameter | 1 | 58.0876 | −0.010 ± 0.001 | <0.0001 |
| Distribution height × Ground diameter | 1 | 7.261 | 0.0001 ± 0.00002 | 0.007 |
| Age | Models | β0 | β1 | R2 | RMSE | ME |
|---|---|---|---|---|---|---|
| 1st + 2nd instar larvae | Spherical | 0.851 | 0.233 | 0.110 | 0.959 | 1.822 |
| Exponential | 0.880 | 0.210 | 0.120 | 0.958 | 1.819 | |
| Gaussian | 0.874 | 0.200 | 0.131 | 0.943 | 1.867 | |
| 3rd instar larvae | Spherical | 1.143 | 0.072 | 0.025 | 0.929 | 1.569 |
| Exponential | 1.194 | 0.089 | 0.020 | 0.908 | 1.564 | |
| Gaussian | 1.113 | 0.044 | 0.030 | 0.902 | 1.733 | |
| 4th instar larvae | Spherical | 0.778 | 0.050 | 0.032 | 1.347 | 1.179 |
| Exponential | 0.777 | 0.036 | 0.021 | 1.357 | 1.194 | |
| Gaussian | 0.766 | 0.051 | 0.027 | 1.354 | 1.218 | |
| Average | Spherical | 1.052 | −0.001 | 0.013 | 0.612 | 0.582 |
| Exponential | 1.052 | −0.001 | 0.013 | 0.613 | 0.598 | |
| Gaussian | 1.057 | 0.005 | 0.005 | 0.596 | 1.023 |
| Age | Model | Range | C0 | C + C0 | LSD | Distribution |
|---|---|---|---|---|---|---|
| 1st, 2nd instar larvae | Gaussian | 51.772 | 0.932 | 1.031 | 0.096 | Aggregate distribution |
| 3rd instar larvae | Gaussian | 117.952 | 0.670 | 0.868 | 0.228 | Aggregate distribution |
| 4th instar larvae | Spherical | 131.278 | 0.365 | 1.178 | 0.690 | Aggregate distribution |
| Average | Spherical | 52.500 | 0.100 | 0.260 | 0.615 | Aggregate distribution |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Chen, L.; Yang, Y.; Zhao, F.; Zhou, C.; Zhang, D. Geostatistical Analysis of the Spatial Variation of Chrysolina aeruginosa Larvae at Different Stages in Desert Ecosystems. Insects 2023, 14, 379. https://doi.org/10.3390/insects14040379
He Z, Chen L, Yang Y, Zhao F, Zhou C, Zhang D. Geostatistical Analysis of the Spatial Variation of Chrysolina aeruginosa Larvae at Different Stages in Desert Ecosystems. Insects. 2023; 14(4):379. https://doi.org/10.3390/insects14040379
Chicago/Turabian StyleHe, Zeshuai, Liangyue Chen, Ying Yang, Fuqiang Zhao, Chunmei Zhou, and Dazhi Zhang. 2023. "Geostatistical Analysis of the Spatial Variation of Chrysolina aeruginosa Larvae at Different Stages in Desert Ecosystems" Insects 14, no. 4: 379. https://doi.org/10.3390/insects14040379
APA StyleHe, Z., Chen, L., Yang, Y., Zhao, F., Zhou, C., & Zhang, D. (2023). Geostatistical Analysis of the Spatial Variation of Chrysolina aeruginosa Larvae at Different Stages in Desert Ecosystems. Insects, 14(4), 379. https://doi.org/10.3390/insects14040379