


Local Habitat Complexity and Its Effects on Herbivores and Predators in Urban Agroecosystems

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study System
2.2. Vegetation and Ground Cover Surveys
2.3. Arthropod Community Surveys
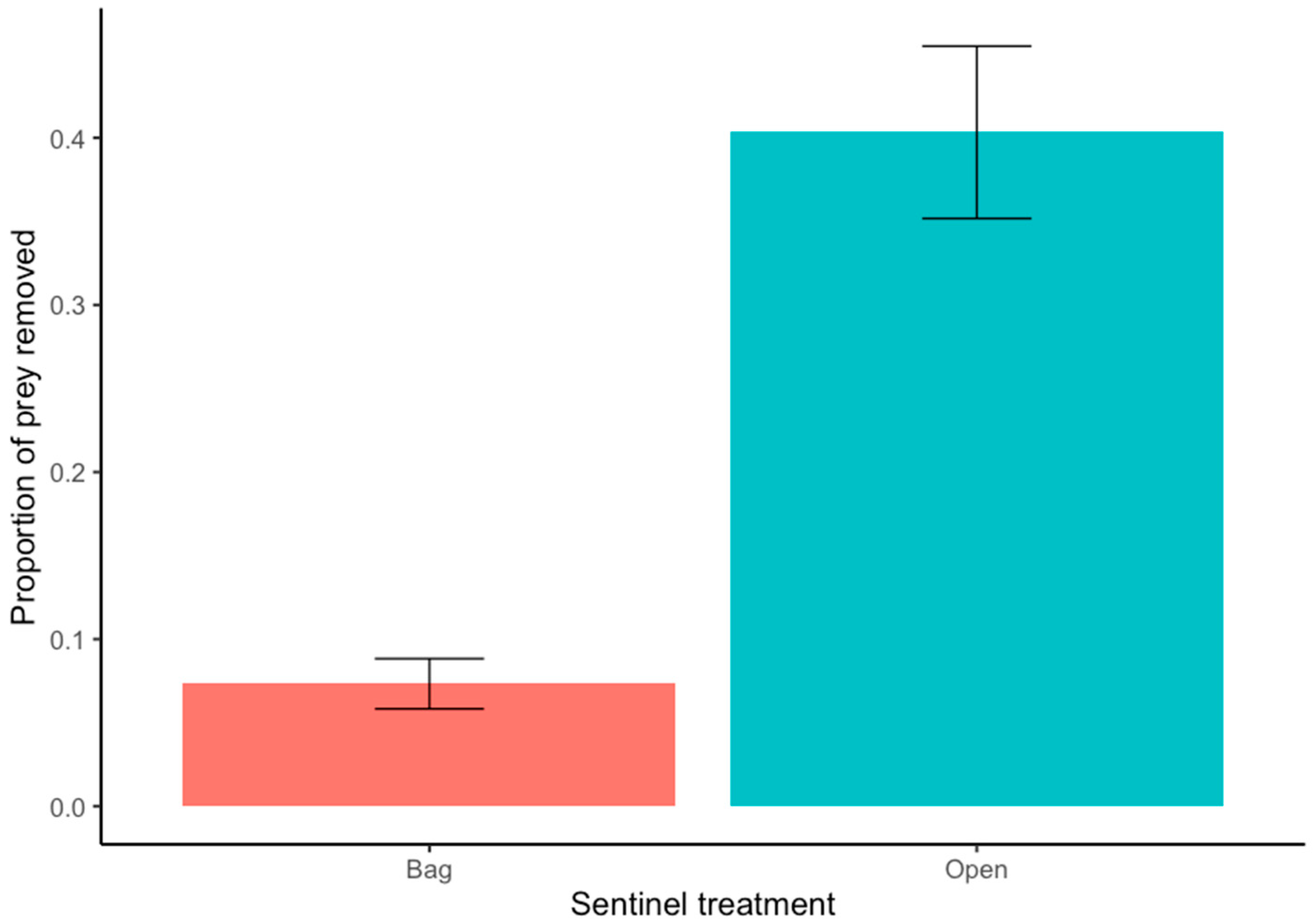
2.4. Sentinel Pest Removal Experiment
2.5. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat Complexity: Approaches and Future Directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Loke, L.H.L.; Chisholm, R.A. Measuring Habitat Complexity and Spatial Heterogeneity in Ecology. Ecol. Lett. 2022, 25, 2269–2288. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H. Patterns of Species Diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Badgley, C.; Smiley, T.M.; Terry, R.; Davis, E.B.; DeSantis, L.R.G.; Fox, D.L.; Hopkins, S.S.B.; Jezkova, T.; Matocq, M.D.; Matzke, N.; et al. Biodiversity and Topographic Complexity: Modern and Geohistorical Perspectives. Trends Ecol. Evol. 2017, 32, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Pianka, E.R. Convexity, Desert Lizards, and Spatial Heterogeneity. Ecology 1966, 47, 1055–1059. [Google Scholar] [CrossRef]
- Dean, R.L.; Connell, J.H. Marine Invertebrates in an Algal Succession. III. Mechanisms Linking Habitat Complexity with Diversity. J. Exp. Mar. Biol. Ecol. 1987, 109, 249–273. [Google Scholar] [CrossRef]
- Pierre, J.I.S.; Kovalenko, K.E. Effect of Habitat Complexity Attributes on Species Richness. Ecosphere 2014, 5, 1–10. [Google Scholar] [CrossRef]
- August, P.V. The Role of Habitat Complexity and Heterogeneity in Structuring Tropical Mammal Communities. Ecology 1983, 64, 1495–1507. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F.; Cassis, G.; Reid, C.A.M. Effects of Habitat Complexity on Forest Beetle Diversity: Do Functional Groups Respond Consistently? Divers. Distrib. 2005, 11, 73–82. [Google Scholar] [CrossRef]
- Uetz, G.W. The Influence of Variation in Litter Habitats on Spider Communities. Oecologia 1979, 40, 29–42. [Google Scholar] [CrossRef]
- Nooten, S.S.; Schultheiss, P.; Rowe, R.C.; Facey, S.L.; Cook, J.M. Habitat Complexity Affects Functional Traits and Diversity of Ant Assemblages in Urban Green Spaces (Hymenoptera: Formicidae). Myrmecol. News 2019, 29, 67–77. [Google Scholar] [CrossRef]
- Huffaker, C.B. Experimental Studies on Predation: Dispersion Factors and Predator-Prey Oscillations. Hilgardia 1958, 27, 343–383. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Intraguild Predation Diminished in Complex-Structured Vegetation: Implications for Prey Suppression. Ecology 2002, 83, 643–652. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; Kremen, C. Pest Control Experiments Show Benefits of Complexity at Landscape and Local Scales. Ecol. Appl. 2012, 22, 1936–1948. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, L.; Munir, S.; Bashir, N.H.; Wang, Y.; Yang, J.; Li, C. Crop Diversity and Pest Management in Sustainable Agriculture. J. Integr. Agric. 2019, 18, 1945–1952. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, C.C.; Williams, N.M. Cropland Heterogeneity Drives Frequency and Intensity of Pesticide Use. Environ. Res. Lett. 2021, 16, 074008. [Google Scholar] [CrossRef]
- Meehan, T.D.; Werling, B.P.; Landis, D.A.; Gratton, C. Agricultural Landscape Simplification and Insecticide Use in the Midwestern United States. Proc. Natl. Acad. Sci. USA 2011, 108, 11500–11505. [Google Scholar] [CrossRef]
- Altieri, M.A.; Letourneau, D.K.; Davis, J.R.; Altieri, M.A.; Letourneau, D.K.; Davis, J.R. Developing Sustainable Agroecosystems. Bioscience 1983, 33, 45–49. [Google Scholar] [CrossRef]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.F.; Ferrer, A.; Peigné, J. Agroecological Practices for Sustainable Agriculture. A Review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef]
- Ong, T.W.Y.; Liao, W. Agroecological Transitions: A Mathematical Perspective on a Transdisciplinary Problem. Front. Sustain. Food Syst. 2020, 4, 91. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a Plant-Arthropod Association in Simple and Diverse Habitats: The Fauna of Collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Ambrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Escobar, S.; Galindo, V.; Gutiérrez, C.; López, S.D.; et al. Does Plant Diversity Benefit Agroecosystems? A Synthetic Review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E. Biodiversity and Ecosystem Function: The Consumer Connection. Oikos 2002, 99, 201–219. [Google Scholar] [CrossRef]
- Ives, A.R.; Cardinale, B.J.; Snyder, W.E. A Synthesis of Subdisciplines: Predator-Prey Interactions, and Biodiversity and Ecosystem Functioning. Ecol. Lett. 2005, 8, 102–116. [Google Scholar] [CrossRef]
- González-Chang, M.; Tiwari, S.; Sharma, S.; Wratten, S.D. Habitat Management for Pest Management: Limitations and Prospects. Ann. Entomol. Soc. Am. 2019, 112, 302–317. [Google Scholar] [CrossRef]
- Redlich, S.; Martin, E.A.; Steffan-Dewenter, I. Landscape-Level Crop Diversity Benefits Biological Pest Control. J. Appl. Ecol. 2018, 55, 2419–2428. [Google Scholar] [CrossRef]
- Russell, E.P. Enemies Hypothesis: A Review of the Effect of Vegetational Diversity on Predatory Insects and Parasitoids Diversity on Predatory Insects and Parasitoids. Environ. Entomol. 1989, 18, 590–599. [Google Scholar] [CrossRef]
- Langellotto, G.A.; Denno, R.F. Responses of Invertebrate Natural Enemies to Complex-Structured Habitats: A Meta-Analytical Synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef]
- Beaumelle, L.; Auriol, A.; Grasset, M.; Pavy, A.; Thiéry, D.; Rusch, A. Benefits of Increased Cover Crop Diversity for Predators and Biological Pest Control Depend on the Landscape Context. Ecol. Solut. Evid. 2021, 2, e12086. [Google Scholar] [CrossRef]
- Burkman, C.E.; Gardiner, M.M. Urban Greenspace Composition and Landscape Context Influence Natural Enemy Community Composition and Function. Biol. Control. 2014, 75, 58–67. [Google Scholar] [CrossRef]
- Randlkofer, B.; Obermaier, E.; Hilker, M.; Meiners, T. Vegetation Complexity—The Influence of Plant Species Diversity and Plant Structures on Plant Chemical Complexity and Arthropods. Basic Appl. Ecol. 2010, 11, 383–395. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Natural Enemy Responses and Pest Control: Importance of Local Vegetation. Biol. Control. 2010, 52, 160–166. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L.; Rasmann, S.; Castagneyrol, B.; Mooney, K.A. Plant Diversity Effects on Insect Herbivores and Their Natural Enemies: Current Thinking, Recent Findings, and Future Directions. Curr. Opin. Insect Sci. 2016, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lawton, J.H. Plant Architecture and the Diversity of Phytophagous Insects. Annu. Rev. Entomol. 1983, 28, 23–39. [Google Scholar] [CrossRef]
- Obermaier, E.; Heisswolf, A.; Poethke, J.; Randlkofer, B.; Meiners, T. Plant Architecture and Vegetation Structure: Two Ways for Insect Herbivores to Escape Parasitism. Eur. J. Entomol. 2008, 105, 233–240. [Google Scholar] [CrossRef]
- Marquis, R.J.; Road, N.B.; Louis, S. Plant Morphology and Recruitment of the Third Trophic Level: Subtle and Little- Recognized Defenses? Oikos 1996, 75, 330–334. [Google Scholar] [CrossRef]
- Inbar, M.; Gerling, D. Plant-Mediated Interactions between Whiteflies, Herbivores, and Natural Enemies. Annu. Rev. Entomol. 2008, 53, 431–448. [Google Scholar] [CrossRef]
- Grevstad, F.S.; Klepetka, B.W. The Influence of Plant Architecture on the Foraging Efficiencies of a Suite of Ladybird Beetles Feeding on Aphids. Oecologia 1992, 92, 399–404. [Google Scholar] [CrossRef]
- Meiners, T.; Obermaier, E. Hide and Seek on Two Spatial Scales—Vegetation Structure Effects Herbivore Oviposition and Egg Parasitism. Basic Appl. Ecol. 2004, 5, 87–94. [Google Scholar] [CrossRef]
- Shrewsbury, P.M.; Raupp, M.J. Evaluation of Components of Vegetational Texture for Predicting Azalea Lace Bug, Stephanitis Pyrioides (Heteroptera: Tingidae), Abundance in Managed Landscapes. Environ. Entomol. 2000, 29, 919–926. [Google Scholar] [CrossRef]
- Parsons, S.E.; Frank, S.D. Urban Tree Pests and Natural Enemies Respond to Habitat at Different Spatial Scales. J. Urban Ecol. 2019, 5, 1–15. [Google Scholar] [CrossRef]
- Peng, M.H.; Hung, Y.C.; Liu, K.L.; Neoh, K.B. Landscape Configuration and Habitat Complexity Shape Arthropod Assemblage in Urban Parks. Sci. Rep. 2020, 10, 16043. [Google Scholar] [CrossRef] [PubMed]
- Lowe, E.C.; Threlfall, C.G.; Wilder, S.M.; Hochuli, D.F.; Lowe Lowe, E.C. Environmental Drivers of Spider Community Composition at Multiple Scales along an Urban Gradient. Biodivers Conserv 2018, 27, 829–852. [Google Scholar] [CrossRef]
- Randlkofer, B.; Obermaier, E.; Casas, J.; Meiners, T. Connectivity Counts: Disentangling Effects of Vegetation Structure Elements on the Searching Movement of a Parasitoid. Ecol. Entomol. 2010, 35, 446–455. [Google Scholar] [CrossRef]
- Kareiva, P.; Perry, R. Leaf Overlap and the Ability of Ladybird Beetles to Search among Plants. Ecol. Entomol. 1989, 14, 127–129. [Google Scholar] [CrossRef]
- Jimenez-Soto, E.; Morris, J.R.; Letourneau, D.K.; Philpott, S.M. Vegetation Connectivity Increases Ant Activity and Potential for Ant-Provided Biocontrol Services in a Tropical Agroforest. Biotropica 2019, 51, 50–61. [Google Scholar] [CrossRef]
- Lin, B.B.; Philpott, S.M.; Jha, S. The Future of Urban Agriculture and Biodiversity-Ecosystem Services: Challenges and next Steps. Basic Appl. Ecol. 2015, 16, 189–201. [Google Scholar] [CrossRef]
- Gardiner, M.M.; Burkman, C.E.; Prajzner, S.P. The Value of Urban Vacant Land to Support Arthropod Biodiversity and Ecosystem Services. Environ. Entomol. 2013, 42, 1123–1136. [Google Scholar] [CrossRef]
- Nighswander, G.P.; Sinclair, J.S.; Dale, A.G.; Qiu, J.; Iannone, B.V. Importance of Plant Diversity and Structure for Urban Garden Pest Resistance. Landsc. Urban Plan. 2021, 215, 104211. [Google Scholar] [CrossRef]
- Oberholtzer, L.; Dimitri, C.; Pressman, A.A. Urban Agriculture in the United States: Characteristics, Challenges, and Technical Assistance Needs. J. Ext. 2014, 52, 28. [Google Scholar] [CrossRef]
- Cabral, I.; Keim, J.; Engelmann, R.; Kraemer, R.; Siebert, J.; Bonn, A. Ecosystem Services of Allotment and Community Gardens: A Leipzig, Germany Case Study. Urban For. Urban Green. 2017, 23, 44–53. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Minor, E.S. Herbivores and Natural Enemies of Brassica Crops in Urban Agriculture. Urban Ecosyst. 2018, 21, 519–529. [Google Scholar] [CrossRef]
- Philpott, S.M.; Bichier, P. Local and Landscape Drivers of Predation Services in Urban Gardens. Ecol. Appl. 2017, 27, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Egerer, M.; Liere, H.; Lucatero, A.; Philpott, S.M. Plant Damage in Urban Agroecosystems Varies with Local and Landscape Factors. Ecosphere 2020, 11, e03074. [Google Scholar] [CrossRef]
- Griffith, G.E.; Omernik, J.M.; Smith, D.W.; Cook, T.D.; Tallyn, E.; Moseley, K.; Johnson, C.B. Ecoregions of California; U.S. Geological Survey: Seattle, WA, USA, 2016.
- Philpott, S.M.; Lucatero, A.; Bichier, P.; Egerer, M.H.; Jha, S.; Lin, B.; Liere, H. Natural Enemy–Herbivore Networks along Local Management and Landscape Gradients in Urban Agroecosystems. Ecol. Appl. 2020, 30, e02201. [Google Scholar] [CrossRef] [PubMed]
- Philpott, S.M.; Albuquerque, S.; Bichier, P.; Cohen, H.; Egerer, M.H.; Kirk, C.; Will, K.W. Local and Landscape Drivers of Carabid Activity, Species Richness, and Traits in Urban Gardens in Coastal California. Insects 2019, 10, 112. [Google Scholar] [CrossRef]
- Iowa State University Department of Entomology BugGuide. Available online: https://bugguide.net/ (accessed on 12 December 2021).
- Borror, D.J.; White, R.E. A Field Guide to Insects: America North of Mexico, 2nd ed.; Houghton Mifflin Harcourt: New York, NY, USA, 1970; Volume 19. [Google Scholar]
- Marshall, S.A. Insects: Their Natural History and Diversity: With a Photographic Guide to Insects of Eastern North America, 2nd ed.; Firefly Books: Buffalo, NY, USA, 2006. [Google Scholar]
- Philpott, S.M.; Bichier, P.; Jha, S.; Lin, B.B.; Liere, H. Landscape, Habitat, and Microhabitat Influences on Predator Abundance and Composition and Prey Removal in Urban Agroecosystems. Biol. Control. 2024; in press. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 December 2023).
- Hartig, F. Package ‘DHARMa’: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6. 2022. Available online: https://cran.r-project.org/web/packages/DHARMa/DHARMa.pdf (accessed on 1 December 2023).
- Fox, J.; Friendly, G.G.; Graves, S.; Heiberger, R.; Monette, G.; Nilsson, H.; Ripley, B.; Weisberg, S.; Zeleis, A. Companion to Applied Regression. The Car Package. R Found. Stat. Comput. 2007, 1109, 1431. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Dai, B.; Scheipl, F.; Grothendieck, G.; Green, P. Package ‘lme4’: Linear Mixed-Effects Models using ‘Eigen’ and S4. R Package Version 1.1-35.1. 2023. Available online: https://cran.r-project.org/web/packages/lme4/lme4.pdf (accessed on 1 December 2023).
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.5.2. 2012. Available online: http://CRAN.R-project.org/package-MuMin (accessed on 1 December 2023).
- Avissar, R. Potential Effects of Vegetation on the Urban Thermal Environment. Atmos. Environ. 1996, 30, 437–448. [Google Scholar] [CrossRef]
- Gómez-Navarro, C.; Pataki, D.E.; Pardyjak, E.R.; Bowling, D.R. Effects of Vegetation on the Spatial and Temporal Variation of Microclimate in the Urbanized Salt Lake Valley. Agric. For. Meteorol. 2021, 296, 108211. [Google Scholar] [CrossRef]
- Andow, D.A. Vegetational Diversity and Arthropod Population Response. Annu. Rev. Entomol. 1991, 36, 561–586. [Google Scholar] [CrossRef]
- Thompson, K.; Austin, K.C.; Smith, R.M.; Warren, P.H.; Angold, P.G.; Gaston, K.J. Urban Domestic Gardens (I): Putting Small-Scale Plant Diversity in Context. J. Veg. Sci. 2003, 14, 71–78. [Google Scholar] [CrossRef]
- Taylor, J.R.; Lovell, S.T.; Wortman, S.E.; Chan, M. Ecosystem Services and Tradeoffs in the Home Food Gardens of African American, Chinese-Origin and Mexican-Origin Households in Chicago, IL. Renew. Agric. Food Syst. 2017, 32, 69–86. [Google Scholar] [CrossRef]
- Mata, L.; Threlfall, C.G.; Williams, N.S.G.; Hahs, A.K.; Malipatil, M.; Stork, N.E.; Livesley, S.J. Conserving Herbivorous and Predatory Insects in Urban Green Spaces. Sci. Rep. 2017, 7, 40970. [Google Scholar] [CrossRef] [PubMed]
- Schlinkert, H.; Westphal, C.; Clough, Y.; László, Z.; Ludwig, M.; Tscharntke, T. Plant Size as Determinant of Species Richness of Herbivores, Natural Enemies and Pollinators across 21 Brassicaceae Species. PLoS ONE 2015, 10, e0135928. [Google Scholar] [CrossRef] [PubMed]
- Shrewsbury, P.M.; Raupp, M.J. Do Top-down or Bottom-up Forces Determine Stephanitis Pyrioides Abundance in Urban Landscapes? Ecol. Appl. 2006, 16, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global Patterns of Guild Composition and Functional Diversity of Spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [PubMed]
- Diehl, E.; Mader, V.L.; Wolters, V.; Birkhofer, K. Management Intensity and Vegetation Complexity Affect Web-Building Spiders and Their Prey. Oecologia 2013, 173, 579–589. [Google Scholar] [CrossRef]
- Delgado de la Flor, Y.A.; Perry, K.I.; Turo, K.J.; Parker, D.M.; Thompson, J.L.; Gardiner, M.M. Local and Landscape-Scale Environmental Filters Drive the Functional Diversity and Taxonomic Composition of Spiders across Urban Greenspaces. J. Appl. Ecol. 2020, 57, 1570–1580. [Google Scholar] [CrossRef]
- Uno, S.; Cotton, J.; Philpott, S.M. Diversity, Abundance, and Species Composition of Ants in Urban Green Spaces. Urban Ecosyst. 2010, 13, 425–441. [Google Scholar] [CrossRef]
- Clarke, K.M.; Fisher, B.L.; Lebuhn, G. The Influence of Urban Park Characteristics on Ant (Hymenoptera, Formicidae) Communities. Urban Ecosyst. 2008, 11, 317–334. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F. Effects of Habitat Complexity on Ant Assemblages. Ecography 2004, 27, 157–164. [Google Scholar] [CrossRef]
- Ossola, A.; Nash, M.A.; Christie, F.J.; Hahs, A.K.; Livesley, S.J. Urban Habitat Complexity Affects Species Richness but Not Environmental Filtering of Morphologically-Diverse Ants. PeerJ 2015, 2015, e1356. [Google Scholar] [CrossRef] [PubMed]
- Norton, B.A.; Thomson, L.J.; Williams, N.S.G.; McDonnell, M.J. The Effect of Urban Ground Covers on Arthropods: An Experiment. Urban Ecosyst. 2014, 17, 77–99. [Google Scholar] [CrossRef]
- Detrain, C.; Verheggen, F.J.; Diez, L.; Wathelet, B.; Haubruge, E. Aphid-Ant Mutualism: How Honeydew Sugars Influence the Behaviour of Ant Scouts. Physiol. Entomol. 2010, 35, 168–174. [Google Scholar] [CrossRef]
- Grover, C.D.; Dayton, K.C.; Menke, S.B.; Holway, D.A. Effects of Aphids on Foliar Foraging by Argentine Ants and the Resulting Effects on Other Arthropods. Ecol. Entomol. 2008, 33, 101–106. [Google Scholar] [CrossRef]
- Santolamazza-Carbone, S.; Velasco, P.; Soengas, P.; Cartea, M.E. Bottom-up and Top-down Herbivore Regulation Mediated by Glucosinolates in Brassica oleracea Var. Acephala. Oecologia 2014, 174, 893–907. [Google Scholar] [CrossRef]
- Kring, J.B. Flight Behavior of Aphids. Annu. Rev. Entomol. 1972, 17, 461–492. [Google Scholar] [CrossRef]
- Byrne, D.N. Migration and Dispersal by the Sweet Potato Whitefly, Bemisia Tabaci. Agric. For. Meteorol. 1999, 97, 309–316. [Google Scholar] [CrossRef]
- Banks, J.E. The Scale of Landscape Fragmentation Affects Herbivore Response to Vegetation Heterogeneity. Oecologia 1998, 117, 239–246. [Google Scholar] [CrossRef]
- Hodge, M.A. The Implications of Intraguild Predation for the Role of Spiders in Biological Control. J. Arachnol. 1999, 27, 351–361. [Google Scholar]
- Saqib, H.S.A.; Liang, P.; You, M.; Gurr, G.M. Molecular Gut Content Analysis Indicates the Inter- and Intra-Guild Predation Patterns of Spiders in Conventionally Managed Vegetable Fields. Ecol. Evol. 2021, 11, 9543–9552. [Google Scholar] [CrossRef] [PubMed]
- Birkhofer, K.; Fließbach, A.; Wise, D.H.; Scheu, S. Generalist Predators in Organically and Conventionally Managed Grass-Clover Fields: Implications for Conservation Biological Control. Ann. Appl. Biol. 2008, 153, 271–280. [Google Scholar] [CrossRef]
- Uiterwaal, S.F.; Squires, A.J.; Grappone, B.A.; Dillard, B.; Castaneda, A.; Kim, S.L.; DeLong, J.P. Intraguild Predation Is Increased in Areas of Low Prey Diversity in a Generalist Predator Community. J. Anim. Ecol. 2023, 92, 901–912. [Google Scholar] [CrossRef]
- Holway, D.A. Competitive Mechanisms Underlying the Displacement of Native Ants by the Invasive Argentine Ant. Ecology 1999, 80, 238–251. [Google Scholar] [CrossRef]
- Human, D.M.; Gordon, K.G. Exploitation and Interference Competition between the Invasive Argentine Ant, I.Inepithema Humile, and Native Ant Species; Springer: Berlin/Heidelberg, Germany, 1996; Volume 105. [Google Scholar]
- Rowles, A.D.; O’Dowd, D.J. Interference Competition by Argentine Ants Displaces Native Ants: Implications for Biotic Resistance to Invasion. Biol. Invasions 2007, 9, 73–85. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Schielzeth, H.; Barnes, A.D.; Barry, K.E.; Bonn, A.; Brose, U.; Bruelheide, H.; Buchmann, N.; Buscot, F.; Ebeling, A.; et al. A Multitrophic Perspective on Biodiversity–Ecosystem Functioning Research. In Advances in Ecological Research; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 61, pp. 1–54. ISBN 9780081029121. [Google Scholar]
- Murphy, S.M.; Richards, L.A.; Wimp, G.M. Editorial: Arthropod Interactions and Responses to Disturbance in a Changing World. Front. Ecol. Evol. 2020, 8, 93. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Response Variable | Model Type | No. Models | R2 * | Predictor Variables | No. Models with Variable | Estimate | z- or t- ** Value | p-Value |
|---|---|---|---|---|---|---|---|---|
| Herbivore abundance | Average | 4 | <0.001–0.05 | Cover | 1 | −0.018 | 1.66 | 0.096 |
| Diversity | 1 | −0.091 | 1.36 | 0.173 | ||||
| Connectivity | 1 | −11.3 | 0.929 | 0.353 | ||||
| Herbivore richness | Average | 2 | <0.001–0.01 | Cover | 1 | 0.005 | 0.932 | 0.351 |
| Predator abundance (VS) | Best | 1 | <0.001 | (Intercept) | NA | 2.00 | 11.8 | <0.001 |
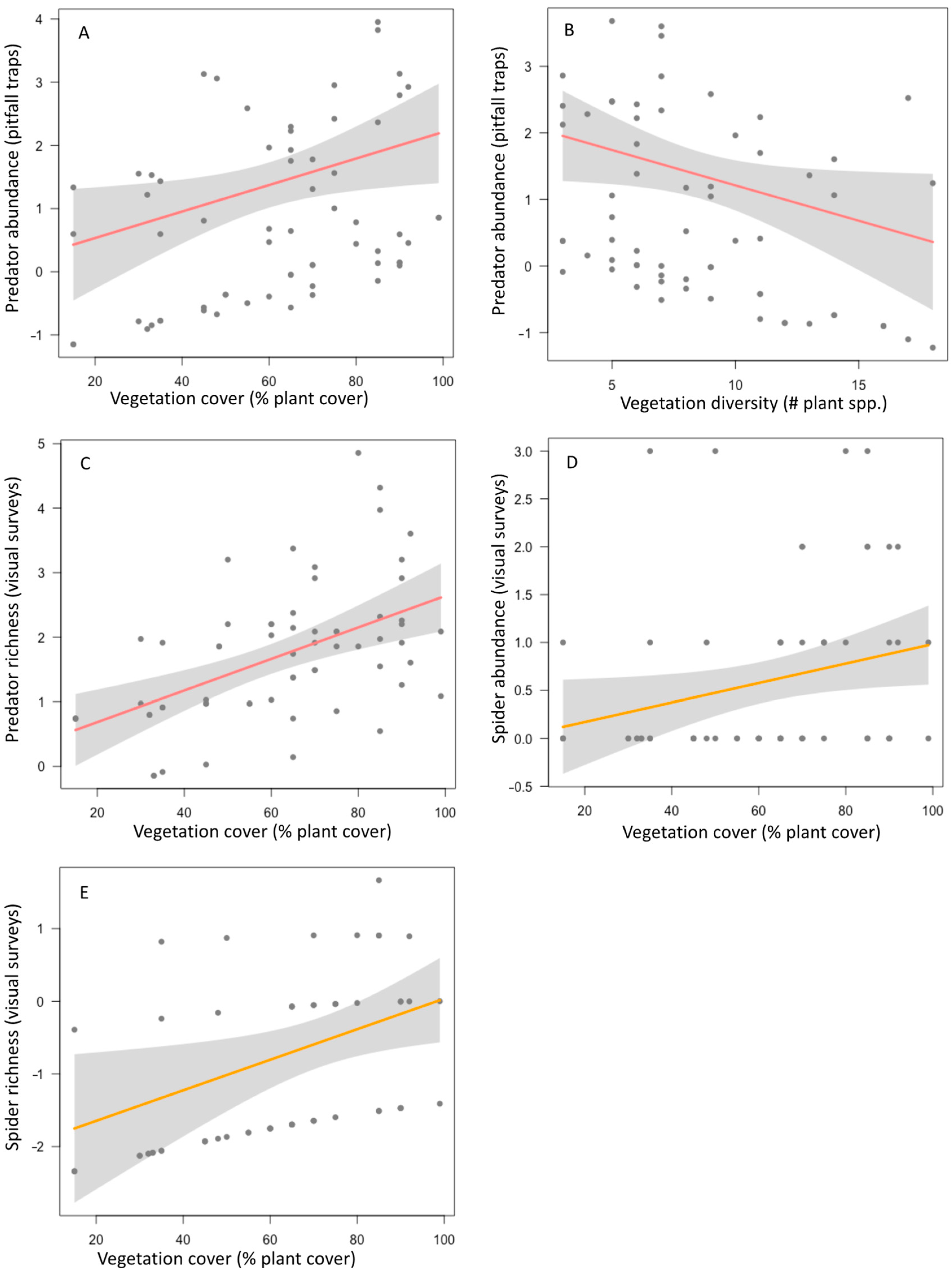
| Predator abundance (PT) | Average | 3 | <0.001–0.072 | Cover | 2 | 0.02 | 2.032 | 0.042 |
| Diversity | 1 | −0.106 | 1.983 | 0.047 | ||||
| Predator richness (VS) | Average | 2 | 0.172–0.204 | Cover | 2 | 0.022 | 3.49 | <0.001 |
| Diversity | 1 | −0.058 | 1.69 | 0.092 | ||||
| Predator richness (PT) | Average | 5 | <0.001–0.048 | Cover | 2 | 0.009 | 1.14 | 0.255 |
| Diversity | 2 | −0.063 | 1.37 | 0.172 | ||||
| Connectivity | 1 | 4.16 | 0.734 | 0.463 | ||||
| Ant abundance (VS) | Average | 2 | <0.001–0.009 | Diversity | 1 | 0.039 | 0.727 | 0.467 |
| Ant abundance (PT) | Average | 5 | <0.001–0.076 | Cover | 2 | 0.021 | 1.87 | 0.062 |
| Diversity | 2 | −0.111 | 1.67 | 0.094 | ||||
| Connectivity | 1 | 8.01 | 0.909 | 0.363 | ||||
| Ant richness (VS) | Average | 2 | 0.039–0.072 | Cover | 2 | 0.009 | 1.28 | 0.201 |
| Diversity | 1 | −0.045 | 1.07 | 0.285 | ||||
| Ant richness (PT) | Average | 3 | <0.001–0.016 | Diversity | 1 | −0.042 | 1.07 | 0.286 |
| Connectivity | 1 | 4.67 | 0.831 | 0.406 | ||||
| Spider abundance (VS) | Average | 2 | 0.072–0.075 | Cover | 2 | 0.01 | 2.02 | 0.044 |
| Diversity | 1 | −0.017 | 0.511 | 0.61 | ||||
| Spider abundance (PT) | Average | 5 | <0.001–0.158 | Cover | 3 | 0.033 | 1.78 | 0.075 |
| Diversity | 1 | −0.136 | 1.47 | 0.143 | ||||
| Connectivity | 2 | 15.6 | 1.19 | 0.232 | ||||
| Spider richness (VS) | Average | 5 | 0.092–0.01 | Cover | 2 | 0.022 | 2.34 | 0.02 |
| Connectivity | 1 | −5.06 | 0.616 | 0.538 | ||||
| Spider richness (PT) | Average | 3 | <0.001–0.111 | Cover | 2 | 0.023 | 1.65 | 0.099 |
| Diversity | 1 | −0.149 | 1.83 | 0.066 | ||||
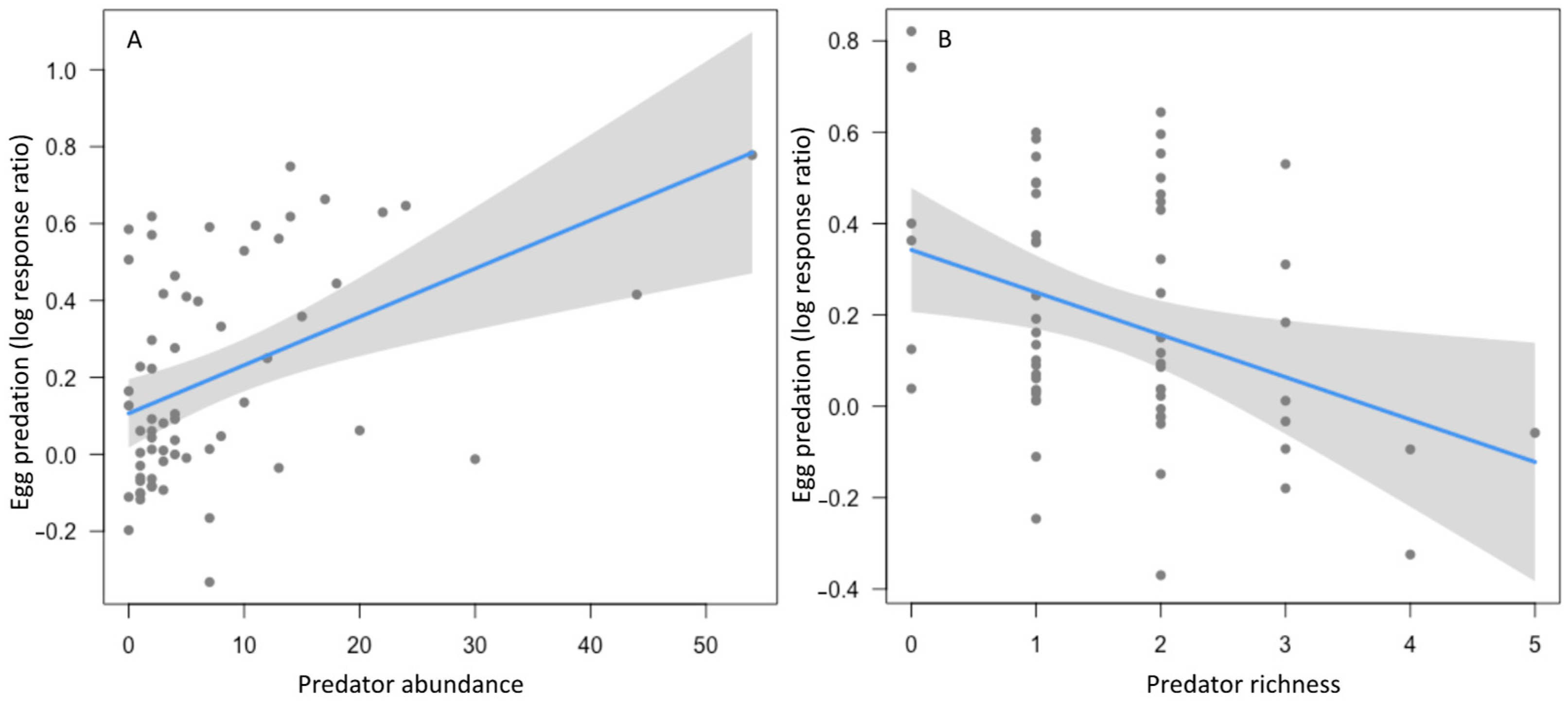
| Egg predation | Average | 4 | 0.12–0.214 | Cover | 2 | 0.003 | 1.52 | 0.128 |
| Diversity | 1 | −0.009 | 0.855 | 0.393 | ||||
| Predator abundance | 4 | 0.012 | 3.25 | 0.001 | ||||
| Predator richness | 3 | −0.084 | 2.1 | 0.036 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucatero, A.; Jha, S.; Philpott, S.M. Local Habitat Complexity and Its Effects on Herbivores and Predators in Urban Agroecosystems. Insects 2024, 15, 41. https://doi.org/10.3390/insects15010041
Lucatero A, Jha S, Philpott SM. Local Habitat Complexity and Its Effects on Herbivores and Predators in Urban Agroecosystems. Insects. 2024; 15(1):41. https://doi.org/10.3390/insects15010041
Chicago/Turabian StyleLucatero, Azucena, Shalene Jha, and Stacy M. Philpott. 2024. "Local Habitat Complexity and Its Effects on Herbivores and Predators in Urban Agroecosystems" Insects 15, no. 1: 41. https://doi.org/10.3390/insects15010041
APA StyleLucatero, A., Jha, S., & Philpott, S. M. (2024). Local Habitat Complexity and Its Effects on Herbivores and Predators in Urban Agroecosystems. Insects, 15(1), 41. https://doi.org/10.3390/insects15010041