



Measurement of Fitness and Predatory Ability of Four Predatory Mite Species in Tibetan Plateau under Laboratory Conditions


Abstract
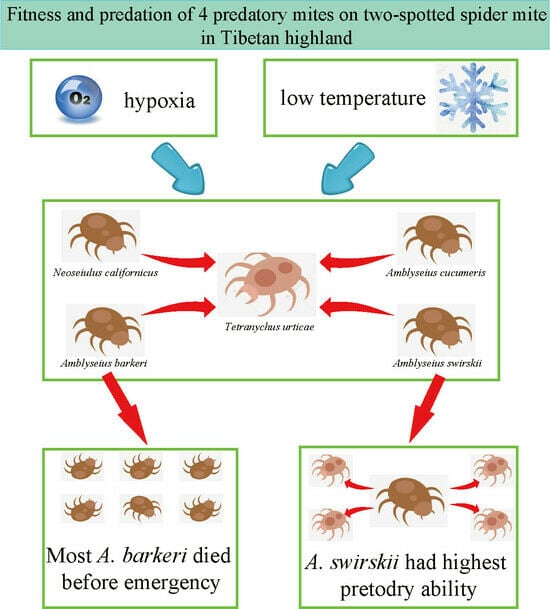
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Establishment of TSSMs and Predatory Mite Populations
2.2. Fitness Experiment for the Four Predatory Mite Species
2.3. Functional Response
2.4. Predator Interactions at Different Predatory Mite Densities
3. Results
3.1. Fitness of Four Predatory Mites at the Tibetan Plateau
3.2. Predatory Mite Fecundity
3.3. Survival Rates at the Tibetan Plateau
3.4. Longevity
3.5. Evaluation of the Predatory Capacity of Predatory Mites
3.6. Functional Responses at Different Developmental Stages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider Mites Web: A comprehensive database for the Tetranychidae. In Trends in Acarology; Springer: Dordrecht, The Netherlands, 2011; pp. 557–560. [Google Scholar]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Dermauw, W.; Grbic, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2013, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Ding, X.L.; Zhang, J.P.; Hong, X.Y. Tetranychus urticae (green form) on Gossypium hirsutum in China: Two records confirmed by aedeagus morphology and RFLP analysis. Syst. Appl. Acarol. 2013, 18, 239–244. [Google Scholar]
- Lu, W.; Wang, M.; Xu, Z.; Shen, G.; Wei, P.; Li, M.; Reid, W.; He, L. Adaptation of acaricide stress facilitates Tetranychus urticae expanding against Tetranychus cinnabarinus in China. Ecol. Evol. 2017, 7, 1233–1249. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.A.; Yang, J.; Luo, B.; Chen, H.Q.; Zhou, J.; Luo, Y.; Wang, Y.X. Regularity of outbreak and prevention of carmine spider mite in Tibet. Tibet J. Agric. Sci. 2011, 33, 27–29. (In Chinese) [Google Scholar]
- Xiang, D.; Huang, H.J.; Wang, Z.; Deqingzhuoga. Preliminary report on control effect of several predator mites on leaf mites of fruit trees. Tibet J. Agric. Sci. 2019, 2, 39–42. (In Chinese) [Google Scholar]
- Elmoghazy, M.M.E.; El-Seidy, E.M.A.; Romeith, A.H.M. Integrated control of the two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) on faba bean Vicia faba (L.) in an open field at Behaira Governorate, Egypt. Int. J. Environ. Sci. Eng. 2011, 2, 93–100. [Google Scholar]
- Escudero, L.A.; Ferragut, F. Life-history of Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) on four spider mite species as prey, with special reference to Tetranychus evansi (Acari: Tetranychidae). Biol. Control 2005, 32, 378–384. [Google Scholar] [CrossRef]
- Toldi, M.; Ferla, N.J.; Dameda, C.; Majelo, F. Biology of Neoseiulus californicus feeding on two-spotted spider mite. Biotemas 2013, 26, 105–111. [Google Scholar] [CrossRef]
- McMurtry, J.A.; Croft, B.A. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol. 1997, 42, 291–321. [Google Scholar] [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourassou, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef]
- Kwon, D.H.; Song, D.Y.; Kang, S.; Ahn, J.J.; Lee, J.H.; Choi, B.R.; Lee, S.W.; Kim, J.H.; Lee, S.H. Residual contact vial bioassay for the on-site detection of acaricide resistance in the two-spotted spider mite. J. Asia-Pac. Entomol. 2010, 13, 333–337. [Google Scholar] [CrossRef]
- Xu, D.D.; He, Y.Y.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. Status of pesticide resistance and associated mutations in the two-spotted spider mite, Tetranychus urticae, in China. Pestic. Biochem. Physiol. 2018, 150, 89–96. [Google Scholar] [CrossRef]
- Vassiliou, V.A.; Kitsis, P. Acaricide resistance in Tetranychus urticae (Acari: Tetranychidae) populations from Cyprus. J. Econ. Entomol. 2013, 106, 1848–1854. [Google Scholar] [CrossRef]
- Wang, X.G.; Xiang, X.; Yu, H.L.; Liu, S.H.; Yin, Y.; Cui, P.; Wu, Y.Q.; Yang, J.; Jiang, C.X.; Yang, Q.F. Monitoring and biochemical characterization of beta-cypermethrin resistance in Spodoptera exigua (Lepidoptera: Noctuidae) in Sichuan Province, China. Pestic. Biochem. Physiol. 2018, 146, 71–79. [Google Scholar] [CrossRef]
- Fan, R.Y.; Wang, B.H.; Zai, Q.; Wang, W.F.; Zhao, Y.Q.; Wei, Q.; Zhang, Y.L.; Cao, L. Composition and occurrence of wheat pests and green control technology in Tibet. Tibet J. Agric. Sci. 2019, 41, 133–137. (In Chinese) [Google Scholar]
- Deqingzhuoga; Xiang, D.; Nima, Y.Z.; Wang, Z.; Chen, H.Q. Study on the characteristics and green prevention and control technology of vegetable pests in plateau. Tibet J. Agric. Sci. 2019, 4, 81–84. (In Chinese) [Google Scholar]
- Zheng, D.; Zhao, D.S. Characteristics of natural environment of the Tibetan Plateau. Sci. Technol. Rev. 2017, 35, 13–22. (In Chinese) [Google Scholar]
- Qi, Y.; Zhang, T.; Wu, Y.; Yao, Z.; Qiu, X.; Pu, P.; Tang, X.; Fu, J.; Yang, W. A multilevel assessment of plasticity in response to high-altitude environment for Agama lizards. Front. Ecol. Evol. 2022, 10, 845072. [Google Scholar] [CrossRef]
- Trochet, A.; Dupoué, A.; Souchet, J.; Bertrand, R.; Deluen, M.; Murarasu, S.; Calvez, O.; Martinez-Silvestre, A.; Verdaguer-Foz, I.; Darnet, E.; et al. Variation of preferred body temperatures along an altitudinal gradient: A multi-species study. J. Therm. Biol. 2018, 77, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Men, S.; Jia, L.; Tang, X.; Storey, K.B.; Niu, Y.; Chen, Q. Comparative metabolomics analysis reveals high-altitude adaptations in a toad-headed viviparous lizard, Phrynocephalus vlangalii. Front. Zool. 2023, 20, 35. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, C.; Zhang, D.; Jiang, H.; Peng, S.; Liu, Y.; Zhao, K.; Wang, C.; Chen, L. Analysis of the erythropoietin of a Tibetan Plateau schizothoracine fish (Gymnocypris dobula) reveals enhanced cytoprotection function in hypoxic environments. BMC Evol. Biol. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.F.; Wang, L.; Qiu, J.P.; Liu, Z.C.; Geng, X.Q. Comparative metabolomics analysis of Callosobruchus chinensis larvae under hypoxia, hypoxia /hypercapnia and normoxia. Pest Manag. Sci. 2017, 73, 1267–1276. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Chen, B.; Zhao, D.J.; Kang, L. Functional modulation of mitochondrial cytochrome c oxidase underlies adaptation to high-altitude hypoxia in a Tibetan migratory locust. Proc. Royal Soc. B 2013, 280, 20122758. [Google Scholar] [CrossRef]
- Liu, T.S.; Zhu, X.; He, D.; You, M.; You, S. Oxygen stress on age-stage, two-sex life tables and transcriptomic response of diamondback moth (Plutella xylostella). Environ. Entomol. 2023, 52, 527–537. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, H.; Luo, Q.; Yang, Y.; Zhang, G.; Zhou, Z.; Naeem, M.; An, J. De novo transcriptomic and metabolomic analyses reveal the ecological adaptation of high-altitude Bombus pyrosoma. Insects 2020, 11, 631. [Google Scholar] [CrossRef]
- Ramalho, Q.; Vale, M.M.; Manes, S.; Diniz, P.; Malecha, A.; Prevedello, J.A. Evidence of stronger range shift response to ongoing climate change by ectotherms and high-latitude species. Biol. Conser. 2023, 279, 109911. [Google Scholar] [CrossRef]
- Ovalle, T.M.; Vásquez-Ordóñez, A.A.; Jimenez, J.; Parsa, S.; Cuellar, W.J.; Lopez-Lavalle, L.A.B. A simple PCR-based method for the rapid and accurate identification of spider mites (Tetranychidae) on cassava. Sci. Rep. 2020, 10, 19496. [Google Scholar] [CrossRef]
- Sun, B.; Zhang, Y.K.; Xue, X.F.; Li, Y.X.; Hong, X.Y. Effects of Wolbachia infection in Tetranychus urticae (Acari: Tetranychidae) on predation by Neoseiulus cucumeris (Acari: Phytoseiidae). Syst. Appl. Acarol. 2015, 20, 591–602. [Google Scholar]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Watt, K.E.F. A mathematical model for the effect of densities of attacked and attacking species on the number attacked. Can. Entomol. 1959, 91, 129–144. [Google Scholar] [CrossRef]
- Atkinson, D.; Sibly, R.M. Why are organisms usually bigger in colder environments? Making sense of a life history puzzle. Trends Ecol. Evol. 1997, 12, 235–239. [Google Scholar] [CrossRef]
- Dahlhoff, E.P.; Dahlhoff, V.C.; Grainger, C.A.; Zavala, N.A.; Otepola-Bello, D.; Sargent, B.A.; Roberts, K.T.; Heidl, S.J.; Smiley, J.T.; Rank, N.E. Getting chased up the mountain: High elevation may limit performance and fitness characters in a montane insect. Funct. Ecol. 2019, 33, 809–818. [Google Scholar] [CrossRef]
- Gaston, K.J.; Chown, S.L.; Evans, K.L. Ecogeographical rules: Elements of a synthesis. J. Biogeogr. 2008, 35, 483–500. [Google Scholar] [CrossRef]
- Greenslade, A.F.C.; Chapman, J.W.; Reynolds, D.R. High-altitude migration of Psylloidea (Hemiptera) over England. Entomol. Gaz. 2021, 72, 189–198. [Google Scholar] [CrossRef]
- McCulloch, G.A.; Waters, J.M. Does wing reduction influence the relationship between altitude and insect body size? A case study using New Zealand’s diverse stonefly fauna. Ecol. Evol. 2018, 8, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Ilias, A.; Vontas, J.; Tsagkarakou, V.A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Guo, Y.; Lv, J.; Jiang, X.; Wang, B.; Gao, Y.; Wang, E.; Xu, X. Intraguild predation between Amblyseius swirskii and two native Chinese predatory mite species and their development on intraguild prey. Sci Rep. 2016, 6, 22992. [Google Scholar] [CrossRef]
- Midthassel, A.; Leather, S.R.; Wright, D.J.; Baxter, I.H. Compatibility of Amblyseius swirskii with Beauveria bassiana: Two potentially complimentary biocontrol agents. BioControl 2016, 61, 437–447. [Google Scholar] [CrossRef]
- Khanamani, M.; Fathipour, Y.; Talebi, A.A.; Mehrabadi, M. Linking pollen quality and performance of Neoseiulus californicus (Acari: Phytoseiidae) in two-spotted spider mite management programmes. Pest Manag. Sci. 2017, 73, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Maroufpoor, M.; Ghoosta, Y.; Pourmirza, A.A. Life table parameters of Neoseiulus californicus (Acari: Phytoseiidae), on the European red mite, Panonychus ulmi (Acari: Tetranychidae) in laboratory condition. Persian J. Acarol. 2013, 2, 265–276. [Google Scholar]
- Rahmani, H.; Fathipour, Y.; Kamali, K. Life history and population growth parameters of Neoseiulus californicus (Acari: Phytoseiidae) fed on Thrips tabaci (Thysanoptera: Thripidae) in laboratory conditions. Syst. Appl. Acarol. 2009, 14, 91–100. [Google Scholar] [CrossRef]
- Ghazy, N.A.; Suzuki, T.; Amano, H. Development and reproduction of Neoseiulus californicus (Acari: Phytoseiidae) and Tetranychus urticae (Acari: Tetranychidae) under simulated natural temperature. Environ. Entomol. 2018, 47, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Marafeli, P.P.; Reis, P.R.; Silveira, E.C.; Souza-Pimentel, G.C.; de Toledo, M.A. Life history of Neoseiulus californicus (McGregor, 1954) (Acari: Phytoseiidae) fed with castor bean (Ricinus communis L.) pollen in laboratory conditions. Braz. J. Biol. 2014, 74, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Greco, N.M.; Sánchez, N.E.; Liljesthröm, G.G. Neoseiulus Californicus (Acari: Phytoseiidae) as a potential control agent of Tetranychus urticae (Acari: Tetranychidae): Effect of pest/predator ratio on pest abundance on strawberry. Exp. Appl. Acarol. 2005, 37, 57–66. [Google Scholar] [CrossRef]
- Olaniyi, O.G.; Rhodes, E.M.; Chase, C.A.; Liburd, O.E. The effect of summer cover crops and strawberry cultivars on the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae) and the predatory mite, Neoseiulus californicus (Acari: Phytoseidae) in organic strawberry production systems in Florida. J. Econ. Entomol. 2021, 114, 2135–2146. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Nguyen, D.T.; Audenaert, J.; Verhoeven, R.; Gobin, B.; Tirry, L.; Clercq, P.D. Prey consumption by phytoseiid spider mite predators as affected by diurnal temperature variations. BioControl 2015, 60, 595–603. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, D.; Wang, Z.; Xu, L.; Wang, Y.; Zhang, H.; Yang, K. Measurement of Fitness and Predatory Ability of Four Predatory Mite Species in Tibetan Plateau under Laboratory Conditions. Insects 2024, 15, 119. https://doi.org/10.3390/insects15020119
Xiang D, Wang Z, Xu L, Wang Y, Zhang H, Yang K. Measurement of Fitness and Predatory Ability of Four Predatory Mite Species in Tibetan Plateau under Laboratory Conditions. Insects. 2024; 15(2):119. https://doi.org/10.3390/insects15020119
Chicago/Turabian StyleXiang, Dong, Zhen Wang, Long Xu, Yunchao Wang, Huanhuan Zhang, and Kun Yang. 2024. "Measurement of Fitness and Predatory Ability of Four Predatory Mite Species in Tibetan Plateau under Laboratory Conditions" Insects 15, no. 2: 119. https://doi.org/10.3390/insects15020119
APA StyleXiang, D., Wang, Z., Xu, L., Wang, Y., Zhang, H., & Yang, K. (2024). Measurement of Fitness and Predatory Ability of Four Predatory Mite Species in Tibetan Plateau under Laboratory Conditions. Insects, 15(2), 119. https://doi.org/10.3390/insects15020119