
The Effects of Temperature on the Development and Survival of Bathycoelia distincta (Hemiptera: Pentatomidae), a Significant Pest of Macadamia in South Africa

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Stock Colony and Rearing
2.2. Effect of Temperature on the Developmental Duration and Survival Rate
2.3. Statistical Analyses
2.4. Temperature-Dependent Models
2.5. Developmental Thresholds and Thermal Constant
3. Results
3.1. Effect of Temperature on the Developmental Duration and Survival Rate
3.2. Effect of Diet on the Developmental Duration and Survival Rate
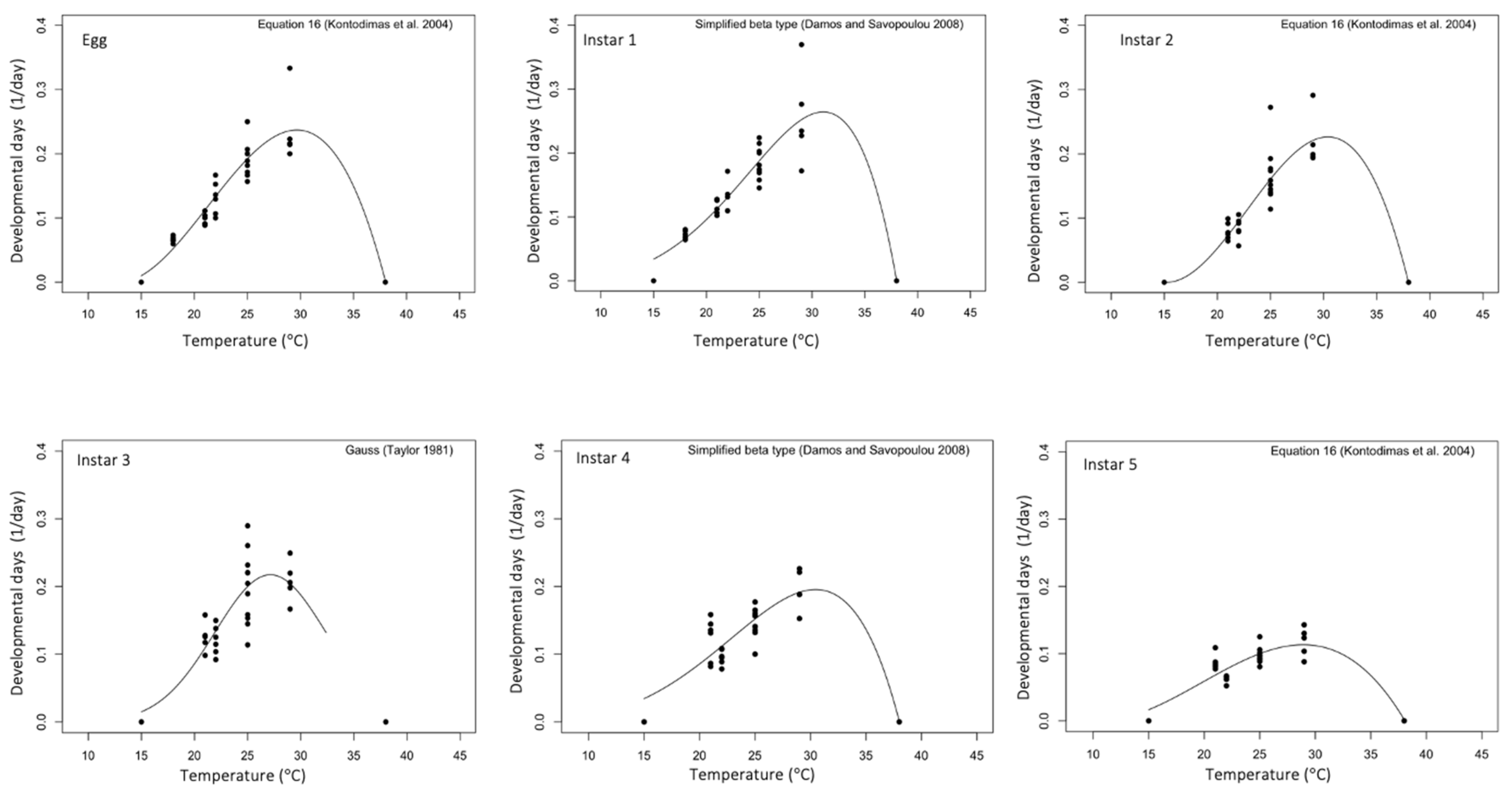
3.3. Temperature-Dependent Models
3.4. Developmental Thresholds and Thermal Constant
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, J.A.; Epstein, M.E. Factors affecting phenology in a temperate insect community. Am. Midl. Nat. 1987, 117, 103–118. [Google Scholar] [CrossRef]
- Herms, D.A. Using degree-days and plant phenology to predict pest activity. IPM (Integr. Pest Manag.) Midwest Landsc. 2004, 58, 49–59. [Google Scholar]
- Herrera, A.M.; Dahlsten, D.D.; Tomic-Carruthers, N.; Carruthers, R.I. Estimating temperature-dependent developmental rates of Diorhabda elongata (Coleoptera: Chrysomelidae), a biological control agent of Saltcedar (Tamarix spp.). Environ. Entomol. 2005, 34, 775–784. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 5, 440. [Google Scholar] [CrossRef] [PubMed]
- Musolin, D.L. Insects in a warmer world: Ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob. Change Biol. 2007, 18, 1565–1585. [Google Scholar] [CrossRef]
- Tougou, D.; Musolin, D.L.; Fujisaki, K. Some like it hot! Rapid climate change promotes changes in distribution ranges of Nezara viridula and Nezara antennata in Japan. Entomol. Exp. Appl. 2009, 3, 249–258. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 6405, 916–919. [Google Scholar] [CrossRef]
- Sharma, H.C. Climate change effects on insects: Implications for crop protection and food security. J. Crop Improv. 2014, 2, 229–259. [Google Scholar] [CrossRef]
- Tonnang, H.E.Z.; Sokame, B.M.; Abdel-Rahman, E.M.; Dubois, T. Measuring and modelling crop yield losses due to invasive insect pests under climate change. Curr. Opin. Insect Sci. 2022, 50, 100873. [Google Scholar] [CrossRef]
- Bale, J.S. Insects and low temperatures: From molecular biology to distributions and abundance. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 1423, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores: Rising temperature and insect herbivores. Glob. Change Biol. 2002, 1, 1–16. [Google Scholar] [CrossRef]
- Savaris, M.; Lampert, S.; Hickmann, F.; Aurélio, M.S.; Schwertner, C.F.; Pereira, P.R. Rearing the green-belly stink bug, Diceraeus furcatus (Fabricius, 1775) (Hemiptera: Pentatomidae) in laboratory. Entomol. Commun. 2022, 4, ec04022. [Google Scholar] [CrossRef]
- Gabre, R.M.; Adham, F.K.; Chi, H. Life table of Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae). Acta Oecologica 2005, 3, 179–183. [Google Scholar] [CrossRef]
- Nietschke, B.S.; Magarey, R.D.; Borchert, D.M.; Calvin, D.D.; Jones, E. A developmental database to support insect phenology models. Crop Prot. 2007, 9, 1444–1448. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 2, 348–355. [Google Scholar] [CrossRef]
- Azrag, A.G.A.; Murungi, L.K.; Tonnang, H.E.Z.; Mwenda, D.; Rabin, R. Temperature-dependent models of development and survival of an insect pest of African tropical highlands, the coffee antestia bug Antestiopsis thunbergii (Hemiptera: Pentatomidae). J. Therm. Biol. 2017, 70, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Damos, P.T.; Savopoulou-Soultani, M. Temperature-dependent bionomics and modeling of Anarsia Lineatella (Lepidoptera: Gelechiidae) in the Laboratory. J. Econ. Entomol. 2008, 5, 1557–1567. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Milliken, G.A. Quantitative analysis of temperature, moisture, and diet factors affecting insect development. Ann. Entomol. Soc. Am. 1988, 4, 539–546. [Google Scholar] [CrossRef]
- Tochen, S.; Woltz, J.M.; Dalton, D.T.; Lee, J.C.; Wiman, N.G.; Walton, V.M. Humidity affects populations of Drosophila suzukii (Diptera: Drosophilidae) in blueberry. J. Appl. Entomol. 2016, 1–2, 47–57. [Google Scholar] [CrossRef]
- Chanthy, P.; Robert, M.; Robin, G.; Nigel, A. Influence of temperature and humidity regimes on the developmental stages of green vegetable bug, Nezara viridula (L.) (Hemiptera: Pentatomidae) from inland and coastal populations in Australia. Gen. Appl. Entomol. 2015, 43, 37–55. [Google Scholar]
- Kingsolver, J.G.; Huey, R. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 2, 251–268. [Google Scholar]
- Kaspari, M.; Alonso, L.; O’Donnellkwd, S. Three energy variables predict ant abundance at a geographical scale. Proc. R. Soc. London. Ser. B Biol. Sci. 2000, 1442, 485–489. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 18, 6668–6672. [Google Scholar] [CrossRef] [PubMed]
- Schoeman, P.S. Phytophagous stink bugs (Hemiptera: Pentatomidae; Coreidae) associated with macadamia in South Africa. Open J. Anim. Sci. 2013, 3, 179–183. [Google Scholar] [CrossRef]
- Geng, S.; Jung, C. Temperature-dependent development of immature Phyllonorycter ringoniella (Lepidoptera: Gracillariidae) and its stage transition models. J. Econ. Entomol. 2018, 4, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Dingha, B.N.; Jackai, L.E. Laboratory rearing of the brown marmorated stink bug (Hemiptera: Pentatomidae) and the impact of single and combination of food substrates on development and survival. Can. Entomol. 2017, 1, 104–117. [Google Scholar] [CrossRef]
- Hope, R.M. Rmisc: Ryan Miscellaneous. R Package Version 1.5.1. 2022. Available online: https://CRAN.R-project.org/package=Rmisc (accessed on 23 July 2023).
- Ahlmann-Eltze, C.; Patil, I. Ggsignif: R package for displaying significance brackets for ‘ggplot2’. 2021. Available online: https://www.researchgate.net/publication/350527292_ggsignif_R_Package_for_Displaying_Significance_Brackets_for_'ggplot2' (accessed on 10 June 2022).
- Patil, I. Visualizations with statistical details: The ggstatsplot approach. J. Open Source Softw. 2021, 61, 3167. [Google Scholar] [CrossRef]
- Rebaudo, F.; Struelens, Q.; Dangles, O. Modelling temperature-dependent development rate and phenology in arthropods: The DEVRATE Package for R ed. Samantha Price. Methods Ecol. Evol. 2018, 4, 1144–1150. [Google Scholar] [CrossRef]
- Taylor, F. Ecology and evolution of physiological time in insects. Am. Nat. 1981, 1, 1–23. [Google Scholar] [CrossRef]
- Harcourt, D.G.; Yee, J.M. Polynomial algorithm for predicting the duration of insect life stages. Environ. Entomol. 1982, 3, 581–584. [Google Scholar] [CrossRef]
- Kontodimas, D.C.; Eliopoulos, P.A.; Stathas, G.J.; Economou, L.P. Comparative Temperature-dependent development of Nephus Includens (Kirsch) and Nephus Bisignatus (Boheman) (Coleoptera: Coccinellidae) preying on Planococcus Citri (Risso) (Homoptera: Pseudococcidae): Evaluation of a linear and various nonlinear models using specific criteria. Environ. Entomol. 2004, 1, 1–11. [Google Scholar] [CrossRef]
- Cullen, E.M.; Zalom, F.G. Phenology-based field monitoring for consperse stink bug (Hemiptera: Pentatomidae) in processing tomatoes. Environ. Entomol. 2000, 3, 560–567. [Google Scholar] [CrossRef]
- Daane, K.M.; Da Silva, P.G.; Stahl, J.M.; Scaccini, D.; Wang, X. Comparative life history parameters of three stink bug pest species. Rondon. Environ. Entomol. 2022, 2, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.G.; Daane, K.M. Life history parameters of Chinavia Hilaris (Hemiptera: Pentatomidae), a stink bug injurious to pistachios in California. J. Econ. Entomol. 2014, 1, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Scriber, J.M.; Slansky, F., Jr. The nutritional ecology of immature insects. Annu. Rev. Entomol. 1981, 1, 183–211. [Google Scholar] [CrossRef]
- Govindan, B.N.; Hutchison, W.D. Influence of temperature on age-stage, two-sex life tables for a minnesota-acclimated population of the brown marmorated stink bug (Halyomorpha Halys). Insects 2020, 2, 108. [Google Scholar] [CrossRef]
- McPherson, J.E.; McPherson, R.M. Stink Bugs of Economic Importance in America North of Mexico; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Taylor, C.M.; Coffey, P.L.; DeLay, B.D.; Dively, G.P. The importance of gut symbionts in the development of the brown marmorated stink bug, Halyomorpha halys (Stål). PLoS ONE 2014, 3, e90312. [Google Scholar] [CrossRef]
- Lockwood, J.A.; Story, R.N. Adaptive functions of nymphal aggregation in the southern green stink bug, Nezara viridula (L.) (Hemiptera: Pentatomidae). Environ. Entomol. 1986, 3, 739–749. [Google Scholar] [CrossRef]
- Muluvhahothe, M.M.; Joubert, E.; Foord, S.H. Thermal tolerance responses of the two-spotted stink bug, Bathycoelia Distincta (Hemiptera: Pentatomidae), vary with life stage and the sex of adults. J. Therm. Biol. 2023, 111, 103395. [Google Scholar] [CrossRef]
- Stuart-Fox, D.; Newton, E.; Clusella-Trullas, S. Thermal consequences of colour and near-infrared reflectance. Philos. Trans. R. Soc. B Biol. Sci. 2017, 1724, 20160345. [Google Scholar] [CrossRef] [PubMed]
- Funayama, K. A new rearing method using carrots as food for the brown-marmorated stink bug, Halyomorpha Halys (Stål) (Heteroptera: Pentatomidae). Appl. Entomol. Zool. 2016, 3, 415–418. [Google Scholar] [CrossRef]
- Fourie, A.; Venter, S.N.; Slippers, B.; Fourie, G. A detection assay to identify alternative food sources of the two-spotted stink bug, Bathycoelia distincta (Hemiptera: Pentatomidae). J. Econ. Entomol. 2022, 2, 519–525. [Google Scholar] [CrossRef]
- Schoeman, P.S. Relative seasonal occurrence of economically significant heteropterans (Pentatomidae and Coreidae) on macadamias in South Africa: Implications for management. Afr. Entomol. 2018, 2, 543–549. [Google Scholar] [CrossRef]
- Sonnekus, B.; Slippers, B.; Hurley, B.P.; Joubert, E.; Stiller, M.; Fourie, G. Diversity and molecular barcoding of stink bugs (Hemiptera: Pentatomidae) associated with macadamia in South Africa. Insects 2022, 7, 601. [Google Scholar] [CrossRef]
- Dhillon, M.K.; Hasan, F. Temperature-dependent development of diapausing larvae of Chilo partellus (Swinhoe) (Lepidoptera: Crambidae). J. Therm. Biol. 2017, 69, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.A.; Ganjisaffar, F.; Palumbo, J.C.; Perring, T.M. Effects of temperatures on immature development and survival of the invasive stink bug Bagrada hilaris (Hemiptera: Pentatomidae). J. Econ. Entomol. 2017, 6, 2497–2503. [Google Scholar] [CrossRef]
- Manzoor, U.; Haseeb, M.; Chander, S.; Moraeit, M. Determination of thermal constant and development threshold of red cotton bug, Dysdercus Cingulatus. Natl. Acad. Sci. Lett. 2020, 4, 10. [Google Scholar] [CrossRef]
- Scaccini, D.; Duso, C.; Pozzebon, A. Lethal effects of high temperatures on brown marmorated stink bug adults before and after overwintering. Insects 2019, 10, 355. [Google Scholar] [CrossRef]
- Ciancio, J.J.; Turnbull, K.F.; Gariepy, T.D.; Sinclair, B.J. Cold tolerance, water balance, energetics, gas exchange, and diapause in overwintering brown marmorated stink bugs. J. Insect Physiol. 2021, 128, 104171. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.P.; Streito, J.C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stinkbug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]

{kind=link}
| Models | Equation | Reference |
|---|---|---|
| Taylor-81 | Taylor 1981 [31] | |
| HartcourtYee-82 | Harcourt and Yee 1982 [32] | |
| Kontodimas-04 | Kontodimas et al. 2004 [33] | |
| Damos-08 | Damos and Savopoulou-Soultani 2008 [17] |
| Developmental Stage | ||||||||
|---|---|---|---|---|---|---|---|---|
| Temperature, Relative Humidity | N | Egg | Instar 1 | Instar 2 | Instar 3 | Instar 4 | Instar 5 | Egg to Adult |
| 18 °C, 29% | 241 | 15.0 ± 0.1 a (222) | 14.2 ± 0.3 a (146) | - | - | - | - | - |
| 21 °C, 91% | 225 | 10.4 ± 0.1 b (192) | 8.5 ± 0.1 b (158) | 12.6 ± 0.3 a (101) | 8.1 ± 0.3 a (83) | 9.1 ± 0.4 ab (73) | 11.8 ± 0.6 a (46) | 60.3 ± 0.6 a |
| 22 °C, 30% | 232 | 7.8 ± 0.1 c (232) | 7.8 ± 0.1 b (199) | 12.2 ± 0.3 a (99) | 8.3 ± 0.5 a (90) | 10.6 ± 0.4 a (79) | 15.7 ± 0.6 b (60) | 62.5 ± 0.6 a |
| 25 °Chigh RH, 64% | 241 | 5.4 ± 0.1 d (232) | 5.3 ± 0.1 c (214) | 5.6 ± 0.1 b (158) | 5.2 ± 0.1 b (128) | 6.4 ± 0.2 bc (111) | 10.4 ± 0.3 ac (82) | 38.4 ± 0.3 b |
| 25 °Clow RH, 25% | 248 | 5.0 ± 0.0 d (243) | 5.9 ± 0.1 c (204) | 7.2 ± 0.1 ab (167) | 5.3 ± 0.2 b (142) | 7.4 ± 0.2 abc (121) | 10.7 ± 0.2 ac (93) | 41.6 ± 0.2 b |
| 29 °C, 23% | 209 | 4.4 ± 0.1 d (194) | 4.1 ± 0.1 c (157) | 4.6 ± 0.1 b (98) | 4.9 ± 0.1 b (94) | 5.2 ± 0.2 c (90) | 8.7 ± 0.2 c (77) | 32.0 ± 0.4 c |
| Macadamia nut and sweetcorn diet comparison | ||||||||
| 25 °C, 71% (Macadamia nut) | 194 | 6.2 ± 0.0 a (194) | 5.2 ± 0.0 a (187) | 7.2 ± 0.1 a (156) | 6.3 ± 0.1 a (150) | 7.0 ± 0.1 a (143) | 10.6 ± 0.1 a (126) | 43.3 ± 0.2 a |
| 25 °C, 64% (Sweetcorn) | 241 | 5.4 ± 0.1 a (232) | 5.3 ± 0.1 a (214) | 5.6 ± 0.1 b (158) | 5.2 ± 0.1 a (128) | 6.4 ± 0.2 b (111) | 10.4 ± 0.3 a (81) | 38.5 ± 0.3 a |
| Developmental Stage | ||||||
|---|---|---|---|---|---|---|
| Temperature, Relative Humidity | Egg | Instar 1 | Instar 2 | Instar 3 | Instar 4 | Instar 5 |
| 18 °C, 29% | 88 ± 0.0 a | 69 ± 0.1 a | 0.0 ± 0.0 a | - | - | - |
| 21 °C, 91% | 95 ± 0.1 a | 76 ± 0.1 a | 66 ± 0.1 bc | 80 ± 0.1 a | 87 ± 0.0 a | 72 ± 0.1 a |
| 22 °C, 30% | 100 ± 0.0 a | 86 ± 0.0 a | 51 ± 0.1 b | 88 ± 0.1 a | 88 ± 0.0 a | 76 ± 0.1 a |
| 25 °Chigh RH, 64% | 96 ± 0.0 a | 92 ± 0.0 a | 73 ± 0.1 bc | 77 ± 0.1 a | 86 ± 0.0 a | 77 ± 0.1 a |
| 25 °Clow RH, 25% | 98 ± 0.0 a | 84 ± 0.1 a | 82 ± 0.1 c | 83 ± 0.0 a | 83 ± 0.0 a | 81 ± 0.1 a |
| 29 °C, 23% | 93 ± 0.0 a | 81 ± 0.0 a | 64 ± 0.0 bc | 95 ± 0.0 a | 97 ± 0.0 a | 84 ± 0.1 a |
| Macadamia nut and sweetcorn diet comparison | ||||||
| 25 °C, 71% (Macadamia nut) | 95 ± 0.0 a | 96 ± 0.0 a | 83 ± 0.1 a | 96 ± 0.0 a | 95 ± 0.0 a | 88 ± 0.1 a |
| 25 °C, 64% (Sweetcorn) | 96 ± 0.0 a | 92 ± 0.0 a | 73 ± 0.1 a | 77 ± 0.1 a | 86 ± 0.0 a | 77 ± 0.1 a |
| Developmental Stage | |||||||
|---|---|---|---|---|---|---|---|
| Model | Parameters | Egg | Instar 1 | Instar 2 | Instar 3 | Instar 4 | Instar 5 |
| Taylor-81 | Rm | 0.23 ± 0.01 *** | 0.23 ± 0.01 *** | 0.21 ± 0.01 *** | 0.21 ± 0.01 *** | 0.17 ± 0.01 *** | 0.11 ± 0.00 *** |
| Tm | 27.9 ± 0.58 *** | 28.4 ± 0.73 *** | 28.4 ± 0.68 *** | 27.1 ± 0.63 *** | 28.0 ± 0.78 *** | 27.5 ± 0.45 *** | |
| To | −5.60 ± 0.48 *** | 6.1 ± 0.59 *** | −4.84 ± 0.58 *** | 5.24 ± 0.72 *** | 6.13 ± 0.79 *** | 6.36 ± 0.55 *** | |
| R2 | 0.04 | 0.005 | 0.006 | 0.02 | 0.007 | 0.03 | |
| AIC | −143.1 | −131.4 | −114.8 | −105.8 | −118.5 | −148.0 | |
| HarcourtYee-82 | a0 | 0.83 ± 0.41. | 9.96 ± 0.05 * | 0.011 ± 0.046 * | 0.011 ± 0.01 | 0.038 ± 0.04 | 0.0044 ± 0.025 |
| a1 | −0.13 ± 0.04 ** | −0.01 ± 0.00 ** | −0.018 ± 0.00 ** | −0.0047 ± 0.00 | −0.0073 ± 0.00 | −0.0021 ± 0.00 | |
| a2 | 0.0073 ± 0.00 *** | 0.00079 ± 0.00 *** | 0.00088 ± 0.00 *** | 0.00037 ± 0.00 | 0.00043 ± 0.00 * | 0.00017 ± 0.00 | |
| a3 | −0.00011 ± 0.00 *** | −0.000013 ± 0.00 *** | −0.00001 ± 0.00 *** | −0.0000056 ± 0.00 * | −0.0000069 ± 0.00 ** | −0.000031 ± 0.00 * | |
| R2 | 0.02 | 0.02 | 0.04 | 0.01 | 0.003 | 0.02 | |
| AIC | −145.3 | −137.3 | −114.8 | −103.7 | −123.0 | −152.5 | |
| Kontodimas-04 | aa | 0.00001 ± 0.00 *** | 0.0000097 ± 0.00 *** | 0.000012 ± 0.00 *** | 0.0000084 ± 0.00 *** | 0.0000069 ± 0.00 *** | 0.0000036 ± 0.00 *** |
| Tmin | 0.011 ± 0.06 *** | 0.013 ± 0.07 *** | 0.015 ± 0.07 *** | 0.011 ± 0.01 *** | 0.012 ± 0.01 *** | 0.011 ± 0.01 *** | |
| Tmax | 0.038 ± 0.04 *** | 0.038 ± 0.05 *** | 0.038 ± 0.05 *** | 0.037 ± 0.06 *** | 0.038 ± 0.06 *** | 0.038 ± 0.06 *** | |
| R2 | 0.02 | 0.02 | 0.04 | 0.005 | 0.003 | 0.02 | |
| AIC | −147.0 | −138.3 | −116.2 | −105.2 | −125.0 | −154.2 | |
| Damos-08 | aa | 0.003 ± 0.00 *** | 0.0024 ± 0.00 *** | 0.001 ± 0.00 * | 0.0042 ± 0.00 * | 0.0029 ± 0.00 ** | 0.002 ± 0.00 ** |
| bb | 4.01 ± 0.03 *** | 3.80 ± 0.03 *** | 3.79 ± 0.03 *** | 3.79 ± 0.06 *** | 3.80 ± 0.04 *** | 3.80 ± 0.05 *** | |
| cc | 3.98 ± 0.15 *** | 4.46 ± 0.27 *** | 5.13 ± 0.40 *** | 3.83 ± 0.40 *** | 4.02 ± 0.33 *** | 3.61 ± 0.31 *** | |
| R2 | 0.03 | 0.02 | 0.07 | 0.003 | 0.02 | 0.01 | |
| AIC | −145.6 | −141.9 | −114.2 | −102.8 | −125.9 | −153.9 | |
| Developmental Stage | ||||||||
|---|---|---|---|---|---|---|---|---|
| Model | Parameter (°C) | Egg | Instar 1 | Instar 2 | Instar 3 | Instar 4 | Instar 5 | Egg to Adult |
| Taylor-81 | Tmin | 10.9 | 10.0 | 13.7 | 11.2 | 9.4 | 8.1 | 11.0 |
| HarcourtYee-82 | NE | NE | 16.1 | 15.3 | 12.9 | 14.1 | 14.1 | |
| Kontodimas-04 | 13.9 | 13.6 | 16.1 | 12.8 | 12.9 | 11.6 | 13.5 | |
| Damos-08 | 7.9 | 7.9 | 9.6 | 6.2 | 6.8 | 5.6 | 7.2 | |
| Taylor-81 | Topt | 27.9 | 28.4 | 28.4 | 27.1 | 28.0 | 27.5 | 27.9 |
| HarcourtYee-82 | 29.7 | 29.9 | 30.4 | 28.9 | 29.3 | 28.7 | 29.5 | |
| Kontodimas-04 | 29.6 | 29.9 | 30.4 | 29.2 | 29.3 | 29.8 | 29.5 | |
| Damos-08 | 30.9 | 31.1 | 31.8 | 30.1 | 30.5 | 29.8 | 30.6 | |
| Taylor-81 | Tmax | NE | 46.9 | 43.1 | 43.1 | 46.7 | 46.8 | 44.9 |
| HarcourtYee-82 | 38.1 | 38.1 | 38.0 | 37.9 | 38.1 | 38.1 | 38.0 | |
| Kontodimas-04 | 38.1 | 38.2 | 38.0 | 37.9 | 38.1 | 38.1 | 38.0 | |
| Damos-08 | 38.0 | 38.0 | 38.0 | 37.9 | 38.0 | 38.0 | 38.0 | |
| Linear | DD | 62.9 | 58.9 | 99.8 | 134.1 | 225.7 | 201.3 | 783.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muluvhahothe, M.M.; Joubert, E.; Foord, S.H. The Effects of Temperature on the Development and Survival of Bathycoelia distincta (Hemiptera: Pentatomidae), a Significant Pest of Macadamia in South Africa. Insects 2024, 15, 152. https://doi.org/10.3390/insects15030152
Muluvhahothe MM, Joubert E, Foord SH. The Effects of Temperature on the Development and Survival of Bathycoelia distincta (Hemiptera: Pentatomidae), a Significant Pest of Macadamia in South Africa. Insects. 2024; 15(3):152. https://doi.org/10.3390/insects15030152
Chicago/Turabian StyleMuluvhahothe, Mulalo M., Elsje Joubert, and Stefan H. Foord. 2024. "The Effects of Temperature on the Development and Survival of Bathycoelia distincta (Hemiptera: Pentatomidae), a Significant Pest of Macadamia in South Africa" Insects 15, no. 3: 152. https://doi.org/10.3390/insects15030152