
Effects of Carbohydrate Intake on Anopheles darlingi and Anopheles deaneorum Fitness under Lab-Reared Conditions

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Blood Digestion
2.3. Fecundity
2.4. Survival
2.5. Mosquito Infection
2.6. Mosquito Dissection
2.7. Statistical Analysis
3. Results
3.1. Blood Digestion
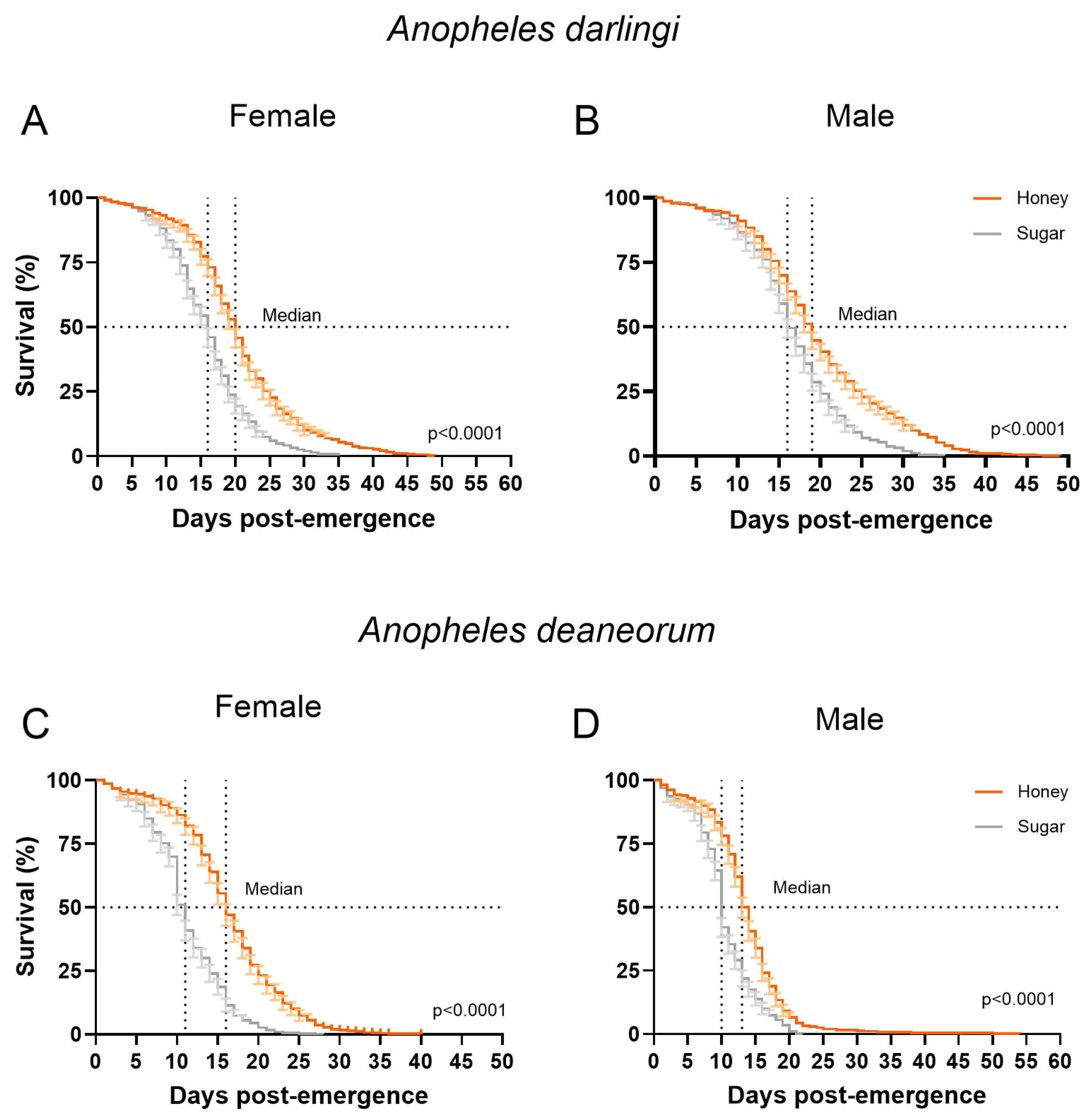
3.2. Survival and Fecundity
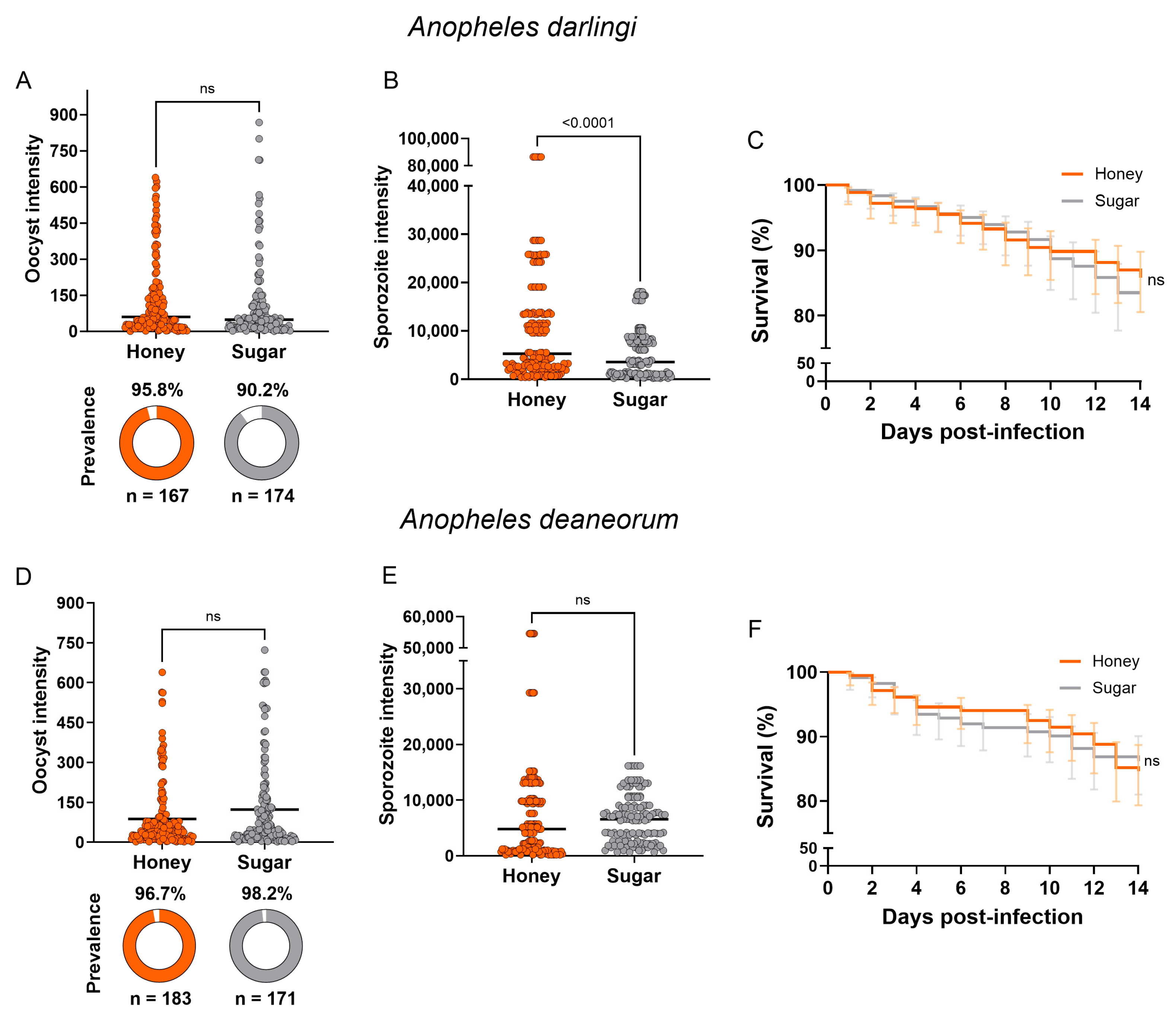
3.3. Mosquito Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tachibana, M.; Takakshima, E.; Morita, M.; Sattabongkot, J.; Ishino, T.; Culleton, R.; Torii, M.; Tsuboi, T. Plasmodium vivax transmission-blocking vaccines: Progress, challenges and innovation. Parasitol Int. 2022, 87, 102525. [Google Scholar] [CrossRef]
- Araujo, M.S.; Andrade, A.O.; Santos, N.A.C.; Pereira, D.B.; Costa, G.S.; Paulo, P.F.M.; Rios, C.T.; Moreno, M.; Pereira-da-Silva, L.H.; Medeiros, J.F. Brazil’s first free-mating laboratory colony of Nyssorhynchus darlingi. Rev. Soc. Med. Trop. 2019, 52, e20190159. [Google Scholar] [CrossRef]
- Araujo, S.A.; Santos, N.A.C.; Andrade, A.O.; Castro, B.C.; Bastos, A.S.; Resadore, F.; Pereira-da-Silva, L.H.; Medeiros, J.F. Description of an automatic copulation induction system used to establish a free-mating laboratory colony of Nyssorhynchus deaneorum from Brazil. Mem. Inst. Oswaldo Cruz. 2020, 115, e200070. [Google Scholar] [CrossRef]
- Vittor, A.Y.; Pan, W.; Gilman, R.H.; Tielsch, J.; Glass, G.; Shields, T.; Sánchez-Lozano, W.; Pinedo, V.V.; Salas-Cobos, E.; Flores, S.; et al. Linkin deforestation to malaria in the Amazon: Characterization of the breeding habitat of the principal malaria vector, Anopheles darlingi. Am. J. Trop. Med. Hyg. 2009, 81, 5–12. [Google Scholar]
- Rosa-Freitas, M.G. Anopheles (Nyssorhynchus) deaneorum: A new species in the Albitarsis complex (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz. 1989, 84, 535–543. [Google Scholar] [CrossRef]
- Klein, T.A.; Lima, J.B.P.; Tada, M.S.; Mil, R. Comparative Susceptibility of Anopheline Mosquitoes in Rondonia, Brazil to Infection by Plasmodium vivax. Am. J. Trop. Med. Hyg. 1991, 45, 463–470. [Google Scholar] [CrossRef]
- Benedict, M.Q.; Knols, B.G.; Bossin, H.C.; Howell, P.I.; Mialhe, E.; Caceres, C.; Robinson, A.S. Colonization and mass rearing: Learning from others. Malar. J. 2009, 8, 4–15. [Google Scholar] [CrossRef]
- Villarreal-Trevino, C.; Vasquez, G.M.; Lopez-Sifuentes, V.M.; Escobedo-Vargas, K.; Huayanay-Repetto, A.; Linton, Y.M.; Flores-Mendoza, C.; Lescano, A.G.; Stell, F.M. Establishment of a free-mating, long-standing and highly productive laboratory colony of Anopheles darlingi from the Peruvian Amazon. Malar. J. 2015, 14, 227. [Google Scholar] [CrossRef]
- Moreno, M.; Tong, C.; Guzmán, M.; Chuquiyauri, R.; Llanos-Cuentas, A.; Rodriguez, H.; Gamboa, D.; Meister, S.; Winzeler, E.A.; Maguina, P.; et al. Infection of laboratory-colonized Anopheles darlingi mosquitoes by Plasmodium vivax. Am. J. Trop. Med. Hyg. 2014, 90, 612–616. [Google Scholar] [CrossRef]
- Puchot, N.; Lecoq, M.T.; Carinci, R.; Duchemin, J.B.; Gendrin, M.; Bourgouin, C. Establishment of a colony of Anopheles darlingi from French Guiana for vector competence studies on malaria transmission. Front. Trop. Dis. 2022, 3, 949300. [Google Scholar] [CrossRef]
- Coluzzi, M. Maintenance of Laboratory colonies of Anopheles mosquitos. Bull. Org. Mond. Santé. 1964, 31, 441–443. [Google Scholar]
- Manda, H.; Gouagna, L.C.; Foster, W.A.; Jackson, R.R.; Beier, J.C.; Githure, J.I.; Hassanali, A. Effect of discriminative plat-sugar feeding on the survival and fecundity of Anopheles gambiae. Malar. J. 2007, 6, 113. [Google Scholar] [CrossRef]
- Vrzal, E.M.; Allan, S.A.; Hahn, D.A. Amino acids in nectar enhance longevity of female Culex quinquefasciatus mosquitoes. J. Insect Physiol. 2010, 46, 1659–1664. [Google Scholar] [CrossRef]
- Gary, R.E.; Foster, W.A. Effects of available sugar on the reproductive fitness and vectorial capacity of the malaria vector Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2001, 38, 22–28. [Google Scholar] [CrossRef]
- Foster, W.A. Mosquito sugar feeding and reproductive energetics. Annu. Rev. Entomol. 1995, 40, 443–474. [Google Scholar] [CrossRef]
- Carvajal-Lago, L.; Ruiz-López, M.J.; Figuerola, J.; Martínez-de la Puente, J. Implications of diet on mosquito life history traits and pathogen transmission. Environ. Res. 2021, 195, 110893. [Google Scholar] [CrossRef]
- Barker, C.M.; Reisen, W.K. Epidemiology of Vector-Borne Diseases. In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: New York, NY, USA, 2019; pp. 33–49. [Google Scholar]
- Gary, R.E., Jr.; Cannonn, J.W., III; Foster, W.A. Effect of sugar on male Anopheles gambiae mating performance, as modified by temperature, space, and body size. Parasit. Vectors. 2009, 2, 19. [Google Scholar] [CrossRef]
- Horosko, S., 3rd; Lima, J.B.; Brandolini, M.B. Establishment of a free-mating colony of Anopheles albitarsis from Brazil. J. Am. Mosq. Control Assoc. 1997, 13, 95–96. [Google Scholar]
- Shute, G.T. A method of maintaining colonies of East African strains of Anopheles gambiae. Ann. Trop. Med. Parasitol. 1956, 50, 92–94. [Google Scholar] [CrossRef]
- Grieco, J.P.; Achee, N.L.; Briceno, I.; King, R.; Andre, R.; Roberts, D.; Rejmankova, E. Comparison of life table attributes from newly established colonies of Anopheles albimanus and Anopheles vestitipennis in northern Belize. J. Vector Ecol. 2003, 28, 200–207. [Google Scholar]
- Timinao, L.; Vinit, R.; Katusele, M.; Schofield, L.; Burkot, T.R.; Karl, S. Optimization of the feeding rate of Anopheles farauti s.s. colony mosquitoes in direct membrane feeding assays. Parasit. Vectors 2021, 14, 356. [Google Scholar] [CrossRef]
- Moreno, M.; Tong-Rios, C.; Orjuela-Sanchez, P.; Carrasco-Escobar, G.; Campo, B.; Gamboa, D.; Winzeler, E.A.; Vinetz, J.M. Continuous supply of Plasmodium vivax sporozoites from colonized Anopheles darlingi in the Peruvian Amazon. ACS Infect. Dis. 2018, 4, 541–548. [Google Scholar] [CrossRef]
- Rios-Velásquez, C.M.; Martins-Campos, K.M.; Simões, R.C.; Izzo, T.; Santos, E.V.; Pessoa, F.A.C.; Lima, J.B.P.; Monteiro, W.M.; Secundino, N.F.C.; Lacerda, M.V.G.; et al. Experimental Plasmodium vivax infection of key Anopheles species from the Brazilian Amazon. Malar. J. 2013, 12, 460. [Google Scholar] [CrossRef]
- Vavra, J.; Undeen, A.H. Nosema algerae n. sp. (Cnidospora, Microsporida) a pathogen in a laboratory colony of Anopheles stephensi Liston (Diptera, Culicidae). J. Protozool. 1970, 17, 240–249. [Google Scholar] [CrossRef]
- Kim, S.J.; Choochote, W.; Jitpakdi, A.; Junkum, A.; Park, S.J.; Min, G.S. Establishment of a self-mating mosquito colony of Anopheles sinensis from Korea. Entomol. Res. 2003, 33, 267–271. [Google Scholar] [CrossRef]
- Dreyer, S.M.; Leiva, D.; Magaña, M.; Pott, M.; Kay, J.; Cruz, A.; Achee, N.L.; Grieco, J.P.; Vaughan, J.A. Fipronil and ivermectin treatment of cattle reduced the survival and ovarian development of field-collected Anopheles albimanus in a pilot trial conducted in nothern Belize. Malar. J. 2019, 18, 296. [Google Scholar] [CrossRef]
- Detinova, T.S.; Bertram, D.S. Age-Grouping Methods in Diptera of Medical Importance, with Special Reference to Some Vectors of Malaria. World Health Organization. Available online: https://iris.who.int/handle/10665/41724(accessed on 3 January 2024).
- Bailey, L. The action of the proventriculus of the worker honeybee, Apis mellifera L. J. Expl. Biol. 1952, 29, 310–327. [Google Scholar] [CrossRef]
- Gooding, R.H.; Cheung, A.C.; Rolseth, B.M. The digestive processes of haematophagous insects. III. Inhibition of trypsin by honey and possible functions of the esophageal diverticula of mosquitoes (Diptera). Can. Ent. 1973, 105, 433–436. [Google Scholar] [CrossRef]
- Gooding, R.H. In vitro properties of proteinases in the midgut of adult Aedes aegypti L. and Culex fatigans (Wiedemann). Comp. Biochem. Physiol. 1966, 17, 115–127. [Google Scholar] [CrossRef]
- Pajot, F.X.; Le Pont, F.; Molez, J.F. Données sur l’alimentation non sanguine chez Anopheles (Nyssorhynchus) darlingi Root, 1926 (Diptera, Culicidae) en Guyane française. Ent. Méd. Parasitol. 1975, 3, 131–134. [Google Scholar]
- Okuda, K.; Caroci, A.; Ribolla, P.; Marinotti, O.; Bianchi, A.G.; Bijovsky, A.T. Morphological and enzymatic analysis on the midgut of Anopheles darlingi during blood digestion. J. Insect Physiol. 2005, 51, 769–776. [Google Scholar] [CrossRef]
- Nayar, J.K.; Sauerman, D.M., Jr. The effects of nutrition on survival and fecundity in Florida mosquitoes. J. Med. Ent. 1975, 12, 220–225. [Google Scholar] [CrossRef]
- Gary, R.E.; Foster, W.A. Anopheles gambiae feeding and survival on honeydew and extra-floral nectar of peridomestic plants. Med. Vet. Entomol. 2004, 18, 102–107. [Google Scholar] [CrossRef]
- Andersson, I.H. The effect of sugar melas and body size on fecundity and longevity of female Aedes communis (Diptera: Culicidae). Physiol. Entomol. 1992, 17, 203–207. [Google Scholar] [CrossRef]
- Okech, B.A.; Gouagna, L.C.; Kabiru, E.W.; Beier, J.C.; Yan, G.; Githure, J.I. Influence of age and previous diet of Anopheles gambiae on the infectivity of natural Plasmodium falciparum gametocytes from human volunteers. J. Insect Sci. 2004, 4, 33. [Google Scholar] [CrossRef]
- Almire, F.; Terhzaz, S.; Terry, S.; McFarlane, M.; Gestuveo, R.J.; Szemiel, A.M.; Varjak, M.; McDonal, A.; Kohl, A.; Pondeville, E. Sugar feeding protects against arboviral infection by enhancing gut immunity in the mosquito vector Aedes aegypti. PLoS Pathog. 2021, 17, e1009870. [Google Scholar] [CrossRef]
- Koella, J.C.; Sorensen, F.L. Effect of adult nutrition on the melanization immune response of malaria vector Anopheles stephensi. Med. Vet. Entomol. 2002, 16, 316–320. [Google Scholar] [CrossRef]
- Nyasembe, V.O.; Tea, P.E.A.; Sawa, P.; Tumlinson, J.H. Plasmodium falciparum infection increases Anopheles gambiae attraction to nectar sources and sugar uptake. Curr. Biol. 2014, 24, 217–221. [Google Scholar] [CrossRef]
- Rivero, A.; Ferguson, H.M. The energetic budget of Anopheles stephensi infected with Plasmodium chabaudi: Is energy depletion a mechanism for virulence? Proc. Biol. Sci. 2003, 270, 1365–1371. [Google Scholar] [CrossRef]
- Basseri, H.R.; Doosti, S.; Akbarzadeh, K.; Nateghpour, M.; Whitten, M.M.A.; Ladoni, H. Competency of Anopheles stephensi mysorensis strain for Plasmodium vivax and the role of inhibitory carbohydrates to block its sporogonic cycle. Malar. J. 2008, 7, 131. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Mosquito Species | Sella Stage | 24 h | 48 h | 72 h | 96 h | ||||
|---|---|---|---|---|---|---|---|---|---|
| Honey (±SD) | Sugar (±SD) | Honey (±SD) | Sugar (±SD) | Honey (±SD) | Sugar (±SD) | Honey (±SD) | Sugar (±SD) | ||
| Anopheles darlingi | Stage 2 | 3.7 (6.4) | 3.5 (6.0) | - | - | - | - | - | - |
| Stage 3 | 63.0 (30.0) | 57.9 (41.1) | 0 (0) | 2.1 (3.6) | - | - | - | - | |
| Stage 4 | 33.3 (35.1) | 38.6 (44.7) | 4.5 (3.9) | 6.3 (10.9) | 0 (0) | 2.4 (4.1) | - | - | |
| Stage 5 | - | - | 65.2 (9.4) * | 27.0 (9.5) * | 8.9 (8.4) | 2.4 (4.1) | 6.1 (10.5) | 0 (0) | |
| Stage 6 | - | - | 30.4 (12.6) | 62.5 (18.8) | 15.8 (8.0) | 14.7 (14.3) | 8.6 (8.3) | 5.6 (4.9) | |
| Stage 7 | - | - | 0 (0) | 2.1 (3.6) | 75.3 (15.2) | 80.6 (7.9) | 85.4 (13.8) | 94.4 (4.9) | |
| Anopheles deaneorum | Stage 2 | 8.6 (6.2) | 1.7 (2.9) | - | - | - | - | - | - |
| Stage 3 | 64.0 (24.6) | 62.2 (26.6) | - | - | - | - | - | - | |
| Stage 4 | 25.8 (21.2) | 36.1 (28.0) | 7.8 (13.6) | 1.9 (3.2) | - | - | - | - | |
| Stage 5 | 1.7 (2.9) | 0 (0) | 53.3 (16.2) | 56.6 (23.8) | 3.9 (3.4) | 1.8 (3.0) | - | - | |
| Stage 6 | - | - | 42.5 (19.3) | 28.7 (31.6) | 13.4 (6.9) | 11.3 (9.9) | 3.7 (6.4) | 4.1 (7.2) | |
| Stage 7 | - | - | 7.3 (2.8) | 11.1 (9.6) | 82.7 (7.0) | 86.9 (11.4) | 96.3 (6.4) | 95.8 (7.2) | |
| Species/Parameters | Honey (±SD) | Sugar (±SD) | p-Value |
|---|---|---|---|
| Anopheles darlingi | |||
| Engorgement rate (%). | 87.4 ± 16.0 | 93.1 ± 8.8 | 0.559 |
| Eggs/female | 27.5 ± 10.2 | 16.4 ± 5.2 | 0.118 |
| Hatching rate (%) | 84.1 ± 10.2 | 75.9 ± 18.9 | 0.481 |
| Anopheles deaneorum | |||
| Engorgement rate (%) | 71.7 ± 19.3 | 70 ± 22.3 | 0.909 |
| Eggs/female | 15.1 ± 10.6 | 21.5 ± 15.5 | 0.519 |
| Hatching rate (%) | 64.7 ± 11.1 | 64.8 ± 11.9 | 0.985 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, N.A.C.; Martins, M.M.; Andrade, A.O.; Bastos, A.S.; Pontual, J.D.C.; Araújo, J.E.; Rocha, M.L.; Medeiros, J.F.; Araujo, M.S. Effects of Carbohydrate Intake on Anopheles darlingi and Anopheles deaneorum Fitness under Lab-Reared Conditions. Insects 2024, 15, 240. https://doi.org/10.3390/insects15040240
Santos NAC, Martins MM, Andrade AO, Bastos AS, Pontual JDC, Araújo JE, Rocha ML, Medeiros JF, Araujo MS. Effects of Carbohydrate Intake on Anopheles darlingi and Anopheles deaneorum Fitness under Lab-Reared Conditions. Insects. 2024; 15(4):240. https://doi.org/10.3390/insects15040240
Chicago/Turabian StyleSantos, Najara A. C., Mirilene M. Martins, Alice O. Andrade, Alessandra S. Bastos, José D. C. Pontual, Jéssica E. Araújo, Marina L. Rocha, Jansen F. Medeiros, and Maisa S. Araujo. 2024. "Effects of Carbohydrate Intake on Anopheles darlingi and Anopheles deaneorum Fitness under Lab-Reared Conditions" Insects 15, no. 4: 240. https://doi.org/10.3390/insects15040240
APA StyleSantos, N. A. C., Martins, M. M., Andrade, A. O., Bastos, A. S., Pontual, J. D. C., Araújo, J. E., Rocha, M. L., Medeiros, J. F., & Araujo, M. S. (2024). Effects of Carbohydrate Intake on Anopheles darlingi and Anopheles deaneorum Fitness under Lab-Reared Conditions. Insects, 15(4), 240. https://doi.org/10.3390/insects15040240