Systematic Palaeontology
Order Neuroptera Linnaeus, 1758.
Family Berothidae Handlirsch, 1906.
Genus
Aggregataberotha Wang, Huang & Wang, 2022 [
10].
Type species.
Aggregataberotha punctate Wang, Huang & Wang, 2022 [
10].
Aggregataberotha paucipunctata Chen, Shi, Ren & Yang sp. nov.
urn:lsid:zoobank.org:act:FFB30C0B-33D1-40A1-A7B1-B954C41FB92E
Material. Holotype: CNU-NEU-MA2018096.
Etymology. The specific name is from the Latin paucipunctatus (meaning “spotless”), referring to the new species having pale and fewer wing spots than another species of Aggregataberotha.
Locality and horizon. Hukawng Valley, Kachin State, northern Myanmar; Lowermost Cenomanian, Upper Cretaceous.
Diagnosis. Forewing with darker pigmentation along wing margin, subcostal veinlets and longitudinal veins with dark intervals or dots, some small light dark speckles at the divergences of veins. Pterostigma pale. In forewings, the proximal branch of RA forked distally, another branch pectinately branched near wing margin; RP1 with light fork before margin branched; MP with deep fork before margin branched; one rp-m and one m-cua present before origin of MP; one cu-aa1 crossvein present after origin of CuP. In hindwings, one short and sigmoidal rp-m crossvein present, one short and straight mp-cu present.
Description. Body minute size. Body length ca. 3.2 mm. Right forewing length 2.9 mm, width 1.3 mm; left forewing length 2.9 mm, width 1.4 mm. Left hindwing length 2.3 mm, width 1.1 mm; right hindwing length 2.4 mm, width 1.1 mm.
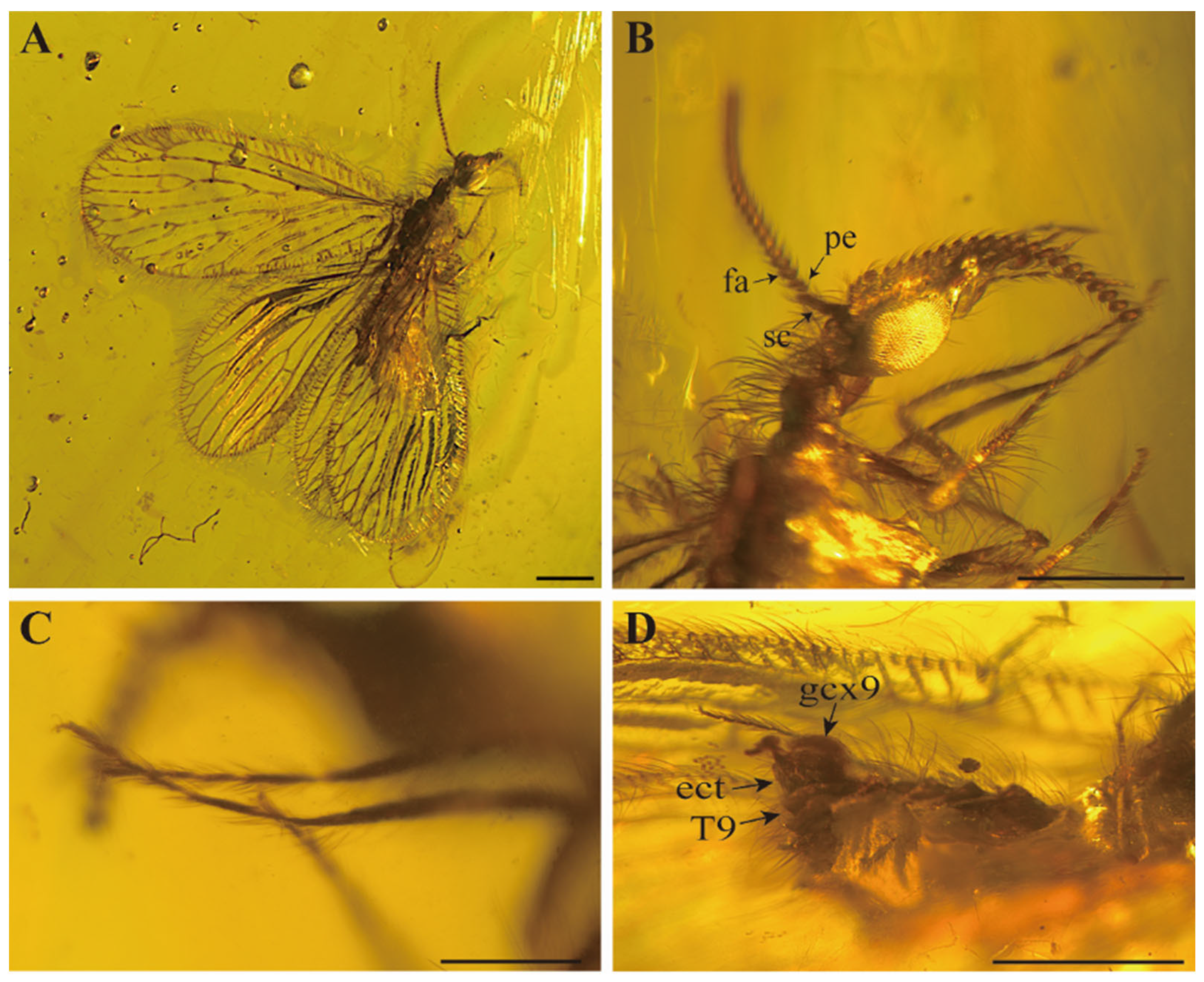
Head (
Figure 1B). Medium position of vertex moderately elevated, bearing some mid-length setae interspersed with few short setae. Oculus oval and well-developed, located on both sides of the head and strongly prominent. Frons elongated and with short setae. Palpus and palpi elongated and slender. Antennae with abundant short setae; scapes slender and medium length, about 2.5 times as long as wide; pedicels oblong; flagellum with about 19 articles and posterior part may be broken, flagellomeres bead-shape.
Thorax. Pronotum slender, about 1.2–1.5 times as long as wide, grouped with long setae. Mesonotum well-developed and subsquare shape. Metanotum shorter than mesonotum, subrectangle and dispersedly covered with cluster setae.
Legs (
Figure 1C). Femurs and tibias slightly expanded in the middle, covered with middle intermixed with short hairs; mesocoxa elongated, stout and tapered; relative length proportion of protarsomeres 1.8–0.8–0.6–0.5–0.7; relative length proportion of mesotarsomeres 4.1–1.4–0.9–0.8–1.3; tibial spurs absent, two curved and thin tarsal claws.
Female (
Figure 1D). Tergite and sternite both closely arranged, tile-like arrangement, moderately sclerotizated, covered with medium-length setae; tergite 7 trapezoid, strong bulge at the end; tergite 8 subrectangle, sternite 8 reduced to small subrectangle; tergite 9 ribbon-shaped, longer than tergite 8, extended to the ventral side; paired 9th gonocoxites, ectoproct obviously separated with tergite 9, subrectangular; hypocaudae absent.
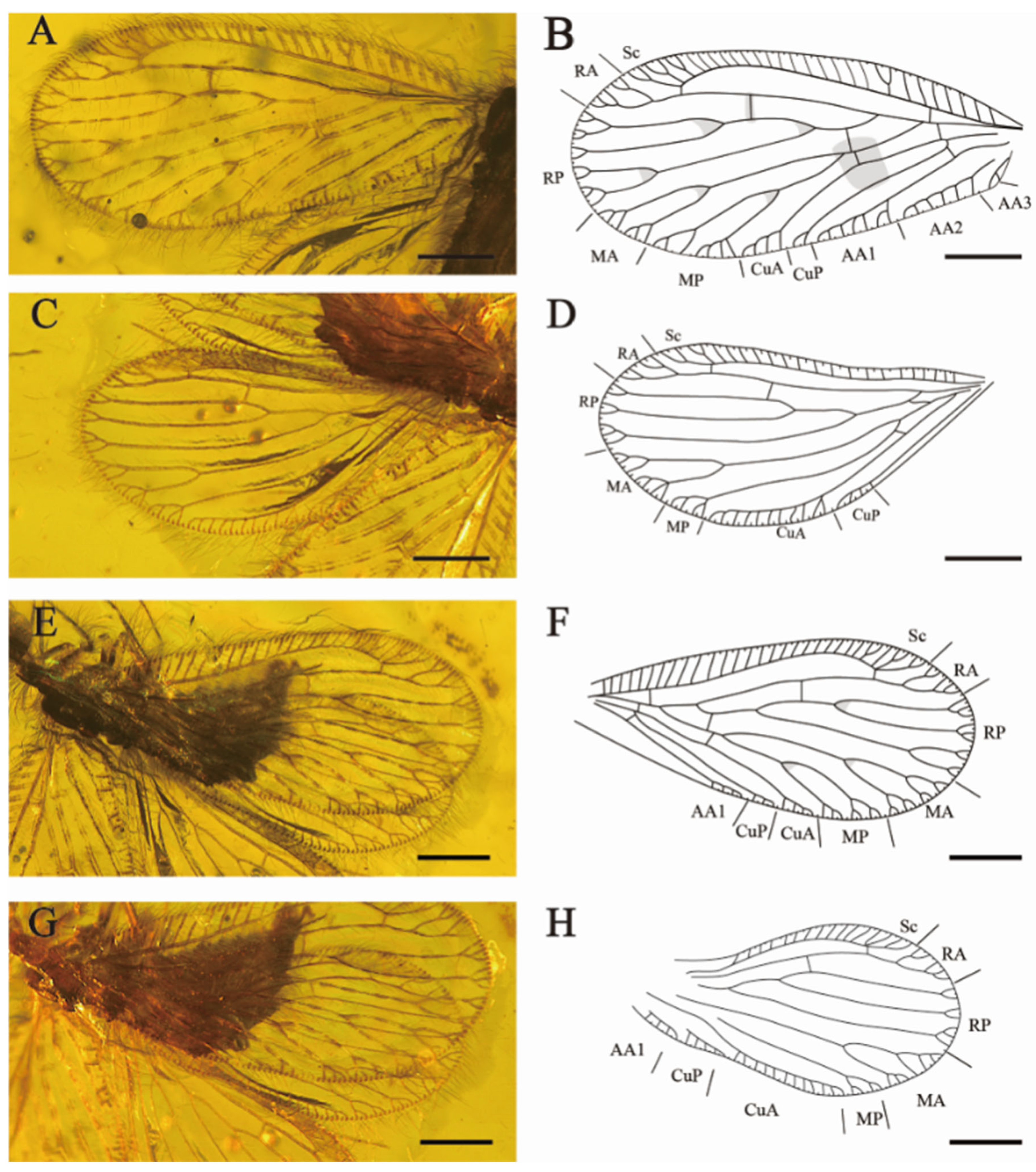
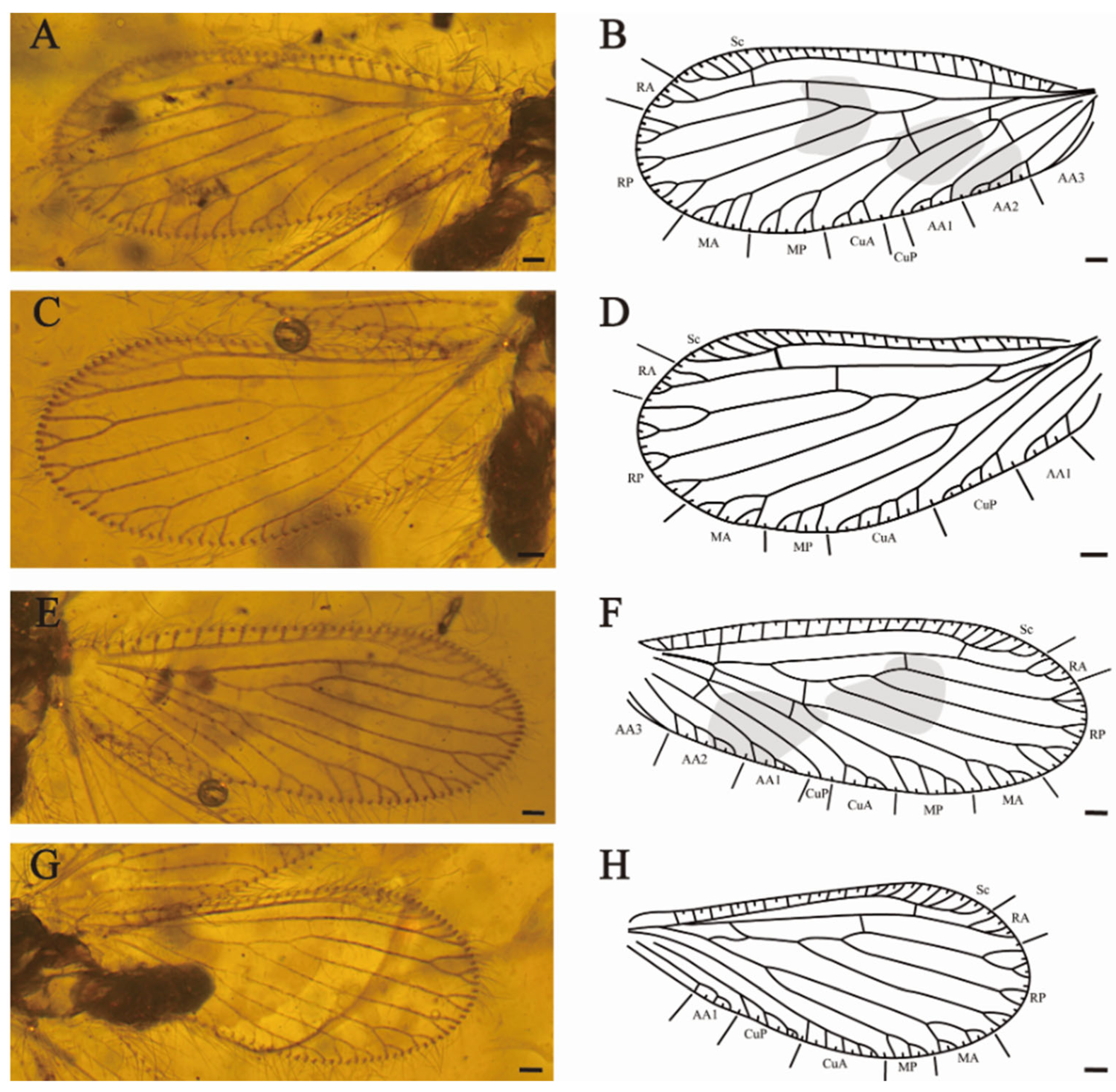
Forewings (
Figure 2A,B,E,F). Wing veins diverge with mottled markings. Most subcostal veinlets simple, three subcostal veinlets forked at distal, the rest simple. Sc and RA free distally, one sc-ra and one sc-r crossvein present; one branch of RA forked distally, another branch pectinately branched near wing margin. RP diverged from RA far from wing base, one ra-rp crossvein lies between the separation of RP1 and RP3, RP1 with light fork before margin forks, branches of RP2 and RP3 bifurcated or trifurcated branched. One rp-m crossvein present. Each branch of MA and MP with pectinate margin forks; MP deeply forked before the margin forks (left forewing, LFW). One rp-m present before the origin of MP. Two m-cu crossveins present. CuA with four short and simple pectinate branches near the wing margin, CuP dichotomous forked distally (LFW). One oblique cup-aa1 present. AA1 with five short pectinate branches and one branch forked, AA2 with six pectinate branches, AA3 simple (LFW). MA ending in two main branches twice dichotomously forked; MP deep fork, the proximal branch dichotomously forked, the distal branch pectinately branched; CuA and CuP pectinate branched near wing margin; AA1 simply pectinate branched (right forewing, RFW).
Hindwings (
Figure 2C,D,G,H) elongated and round. Two subcostal veinlets forked near the apex of Sc. Sc and R free distally. One ra-rp crossvein present, RP with two bifurcated branches and one trifurcated branch. One short and sigmoidal rp-m crossvein present. MA dichotomous forked, each branch again pectinately branched near wing margin. MP forked before marginal dichotomously branched. One oblique mp-cua crossvein present beyond origin of MP. CuA with nine short, simple pectinate branches and one branch forked (LHW), with twelve simple pectinate branches (RHW), parallel to the posterior of wing margin. CuP with at least four simple branches. Anal vein with four simple and short pectinate branches.
Remark. Aggregataberotha paucipunctata sp. nov. belongs to Aggregataberotha based on the following characteristics: (1) mottled markings or dots along subcostal veinlets and longitudinal veins, (2) RP with three main branches, (3) most costal crossveins simple and humeral vein not recurrent, (4) one ra-rp present between origin of RP1 and RP3 in forewings but located at RP3 in hindwings, (5) gradate series absent both in fore- and hindwings, (6) stem of M joined with R near the basal one fifth of the wing length, and then divergent from RP, MA and MP both deep dichotomously branched, (7) paired gonocoxite 9 boat-like laterally and without hypocaudae, (8) ectoproct and tergite 9 distinctly unfused in female.
A. paucipunctata sp. nov. is obviously distinguished from A. punctate by the following characteristics of the venation of wings: in forewing, (1) pale pterostigma present, nearly transparent light black patch at the divergences of veins (vs. dark and distinct pterostigma, fuscous speckles present in A. punctate), (2) one cup-aa1 present beyond the origin of CuP (vs. cup-aa1 absent in A. punctate); in hindwings, (1) a sigmoidal m-cu located near the wing base, (2) one oblique and short mp-cua present beyond the origin of MP (vs. mp-cua absent in A. punctate).
Genus Sejunctaberotha Chen, Shi, Ren & Yang gen. nov.
urn:lsid:zoobank.org:act:3BCF5C46-FA17-4CE9-A1E8-ECA0C9697003
Type species: Sejunctaberotha transversa Chen, Shi, Ren & Yang gen. et sp. nov.
Species included: Sejunctaberotha sphaerica gen. et sp. nov.; Sejunctaberotha tenuis gen. et sp. nov.; Sejunctaberotha transversa gen. et sp. nov.
Etymology. The generic name is a combination of the Latin sejuncta (meaning “separated”) and Berotha, a generic name of the family Berothidae, referring to the abnormal vein character of Sc and R free distinctly both in fore- and hindwings of the new genus. Gender feminine.
Diagnosis. Body small to medium size. Antennae moniliform and antennae with the number of total segments less than 40; scapes not larger elongated. Both in forewings and hindwings, pterostigma pale; without recurrent humeral vein; subcostal veinlets all simple; Sc and RA free distally, connected by a crossvein; after the connection point of the crossvein, RA most with more than three simple marginal branches; only one ra-rp crossvein connected RA and RP3; RP with three major branches; without gradate veins. In forewings, each branch of RP with two to three short and simple branches near wing margin; MA originated from R at basal 1/5 of wing length, MA and MP separate at about the half length of the wing; one rp-m crossvein located before the origin of MP; MP slightly forked before margin branched; two m-cua crossveins present; CuA pectinately branched near wing margin, CuP simple; AA1 with less than four simple branches; AA2 with pectinate branches. Pterostigma pale. Female genitalia simple and all sternites without any modification; tergite 9 and ectoproct not fused; callus cerci, hypocauda and pseudohypocauda absent.
Remarks.
Sejunctaberotha gen. nov. is most related to three extinct genera, i.e.,
Sibelliberotha Azar & Nel, 2013,
Aggregataberotha and
Haploberotha Engel & Grimaldi, 2008, but is obviously different from those by the following combined characteristics: (1) subcostal veinlets all simple (
Aggregataberotha with more than two forked subcostal veinlets near the terminal of Sc); (2) vein of Sc and RA obviously free at apex both in forewings and hindwings (other fossil genera fused distantly both in fore- and hindwings or only free at hindwings); (3) one ra-rp crossvein connected RA and RP3 (one ra-rp crossvein lies between the separation of RP1 and RP3 in
Aggregataberotha and
Sibelliberotha); (4) each branch of RP with two or three short and simple branches near wing margin (
Aggregataberotha and
Haploberotha RP with fork before margin branched,
Sibelliberotha RP only with two short branches near wing margin); (5) in forewings, MP with light fork before margin branched (
Aggregataberotha MP with deep fork before margin branched,
Sibelliberotha MP with pectinate branches near wing margin); (6) one rp-m present before the origin of MP (one rp-m present after the orgin of MP and connect RP with MP in
Sibelliberotha and
Haploberotha); (6) in forewings, CuP simple (
Sibelliberotha and
Aggregataberotha with two to three short branches near wing margin,
Haploberotha with two forked branches) [
10,
28,
29,
30].
Sejunctaberotha sphaerica Chen, Shi, Ren & Yang gen. et sp. nov.
urn:lsid:zoobank.org:act:BE7AAEC9-B910-4169-94FD-0E1BF710E556
Material. Holotype: CNU-NEU-MA2018097.
Etymology. The specific epithet is from the Latin sphaerica (meaning “spherical”), referring to a pair of spherical bulges at the ends of the antennae.
Locality and horizon. Hukawng Valley, Kachin State, northern Myanmar; Lowermost Cenomanian, Upper Cretaceous.
Diagnosis. Vertex covered with scattered, short and fine setae. RA with three simple branches at distal; MA dichotomously branched at distal; elongated and sigmoidal crossvein linked RP and M in hindwings; ectoproct very broad and obviously bigger than tergite 9.
Description. Body length ca. 2.2 mm. Right forewing length 2.1 mm, width 0.9 mm; left forewing length 2.5 mm, width 0.7 mm. Left hindwing length 1.9 mm, width 0.7 mm; right hindwing length 1.6 mm, width 0.9 mm.
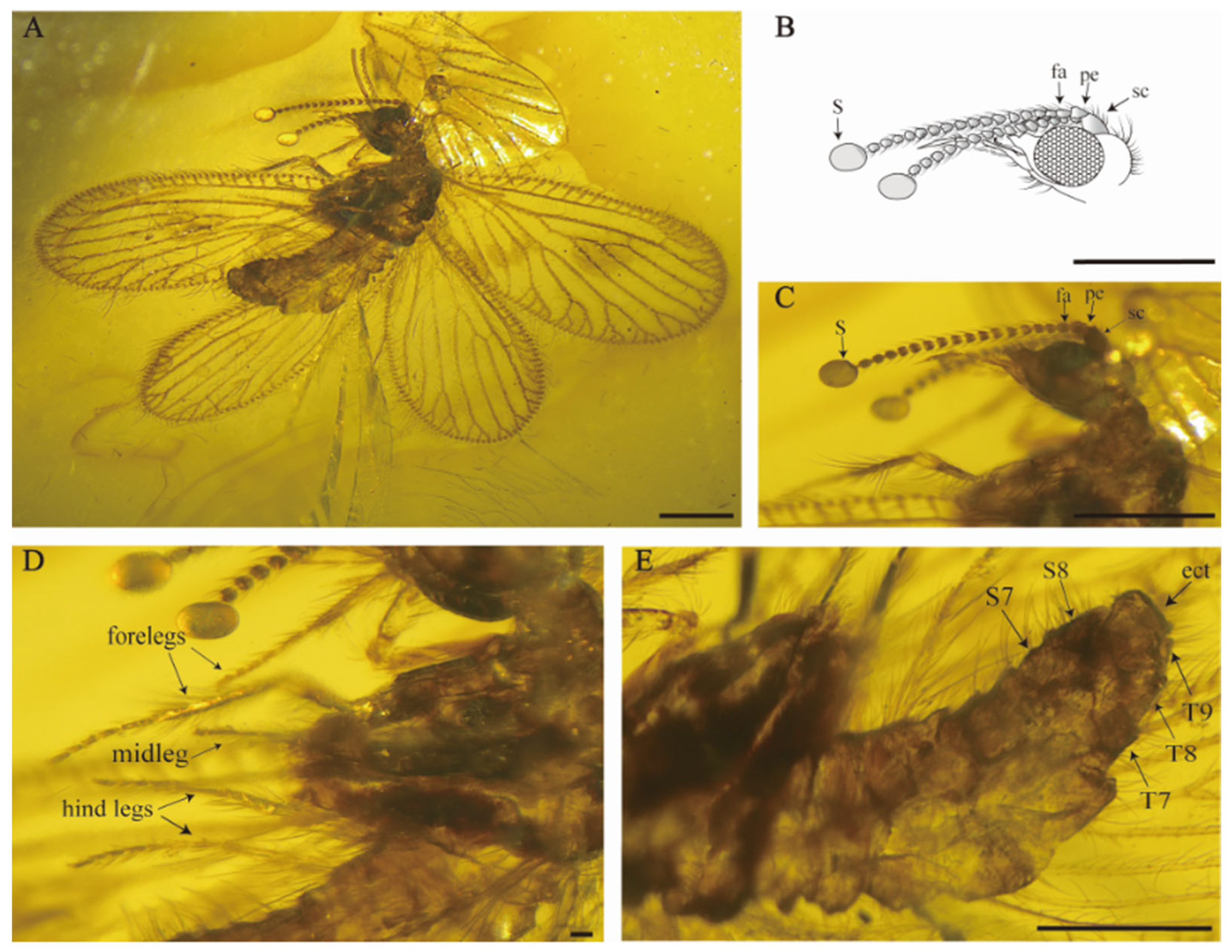
Head (
Figure 3B,C). Vertex moderately swelled, higher than oculus, with abundant long, fine setae; gena expanded; frons and mouthparts elongated, with medium interspersed with small setae. Compound eyes protruded in a hemispherical shape and well developed, antennae moniliform, shorter than body length, bearing ring of short and bent setae; scapus short and stout, about 1.5 times as long as wide; pedicel subsquare, length almost equal to width; flagellum with at least 14 articles, flagellomeres subquadrangular, a pair of spherical bulges present at the end of the antennae.
Body medium size. Pronotum subrectangle, one transverse furrow present, about 1.5 times as wide as long, medium-sized brown setae spread on the later edge. Mesonotum and metanotum with similar shape and color, slightly wider than long.
Legs (
Figure 3D) slender; femur slender, short setae covered both at ends while middle part located with mix of stout long and medium-length fine setae; tarsus densely distributed with short setae, with tarsus 5 segmented, the first tarsomere longest; without tibial spurs; tarsal claws slender and curved; arolium present. Forelegs are walking legs. Procoxa elongated, subcylindrical, entire surface covered with short setae, relative length of tarsomeres 2.7–1.7–1.3–1.1–1.9.
Female genitalia (
Figure 3E). Abdomen slightly swollen. Segments 1–8 well developed and subrectangle, sternite 7 in lateral view subquadrate, sternite 7 and sternite 8 both one piece, without modification; tergite 9 not elongated, subtriangular and distinctly smaller and shorter than tergite 8; ectoproct one piece, obviously bigger than tergite 9; tergite 9 is distinctly separate from ectoproct, without hypocaudae; tergite and ectoproct bearing scattered, mid-length fine hairs.
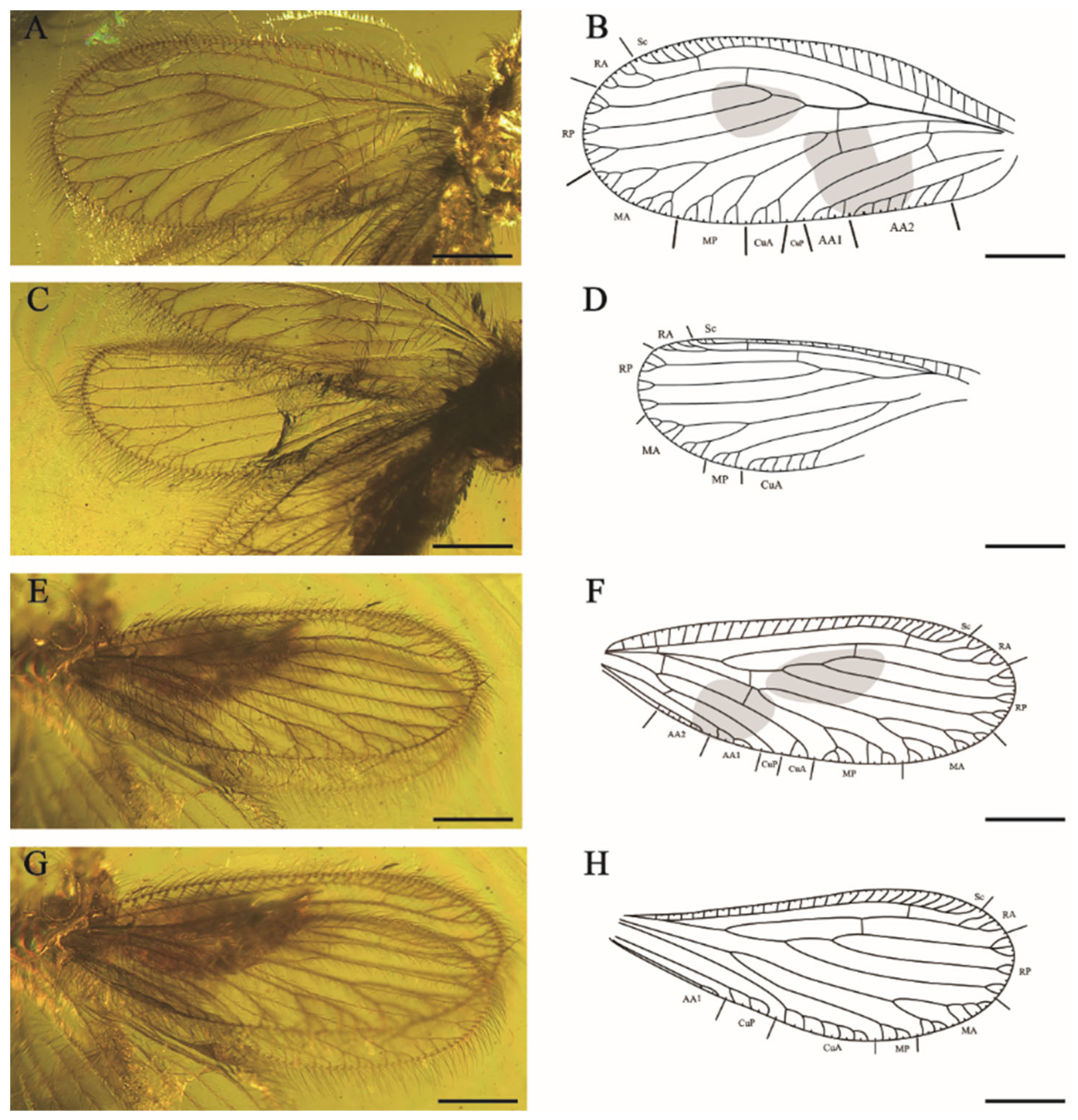
Forewings (
Figure 4A,B,E,F) elongated oblong with rounded apex, membrane hyaline. Dense medium-sized setae and prominent trichosors along the whole wing margin, fine setae scatteredly located on venation. Costal space narrow at base, first slightly expanded proximate the base of the wing at one quarter of the wing length, maximally expanded in the end of subcostal vein. Recurrent humeral vein absent. All subcostal veinlets simple. Sc and RA free at apex and connected with a crossvein. Two crossveins present between Sc and RA. RA with simple pectinate branches near wing margin. RP diverged from RA far apart from wing base, with three major branches, and one crossvein between RA and RP connecting RA and RP3, RP branches bifurcately or trifurcately branched at apex. MA amalgamated with stem of racial vein near 1 sc-r, one rp-m crossvein present before origin of MP, MA and MP both shallowly forked, with simple distal dichotomies. Two m-cua crossveins present beyond origin of CuP, CuA with four pectinate branches close to posterior wing margin, CuP simple. One cup-aa1 crossvein present near origin of the CuP, AA1 dichotomous branched near wing edge, AA2 with four pectinate branches, AA3 not detected (RFW); AA1 and AA2 with three to four short and pectinate branches, AA3 simple (LFW).
Hindwings (
Figure 4C,D,G,H) elongated oval, obviously smaller and shorter than forewings. Costal space narrow at wing base, then typically expanded in pterostigmal region. Subcostal veinlets simple without fork. Sc and RA not fused distally, connected by one sc-r crossvein. RA with three distal simple branches, one ra-rp crossvein connected the RA and RP3, RP with three distal forked branches. One short and sigmoidal crossvein linked RP and M; MA dichotomously branched at distal, each branch bifurcated (LHW), the proximal branch trichotomously forked and the distal branch twice forked (RHW); MP with three simple distal branches (LHW), MP bifurcated (RHW); CuA with six to seven pectinate branches almost parallel to posterior wing edge, CuP with at least four pectinate branches. AA1 with three pectinate branches near wing margin.
Sejunctaberotha tenuis Chen, Shi, Ren & Yang gen. et sp. nov.
urn:lsid:zoobank.org:act:7B54281A-9155-4FB2-805C-82F5616B85C3
Material. Holotype: CNU-NEU-MA2018098.
Etymology. The specific epithet is from the Latin tenuis (meaning “thin”), referring to the dorsal plate of the prothorax being narrower and slightly wider than the length.
Locality and horizon. Hukawng. Valley, Kachin State, northern Myanmar; Lowermost Cenomanian, Upper Cretaceous.
Diagnosis. Protothorax slender, longer than wide, with middle length bristle; ectoproct very broad and obviously bigger than tergite 9 and 9th gonocoxites. Vertex and pronotum bearing with clustered long hairs. RA and MA both dichotomously branched in forewings. No sigmoidal or straight crossvein between basal of RP and M in hindwings. Entire wing margin with dense and medium-length hairs.
Description. Body length ca. 1.2 mm. Right forewing length 2.8 mm, width 1.0 mm; left forewing length 2.7 mm, width 1.2 mm. Left hindwing length 2.3 mm, width 1.0 mm; right hindwing length 2.6 mm, width 1.0 mm.
Head (
Figure 5A,B). Oculus oval shaped. Vertex slightly prominent, bearing dense, long setae. Frons and mouthparts elongated, shorter than the other species of this new genus. Antenna with short scape, ca. 1.5 times as long as wide, covered with short setae; pedicel subsquare, about 3 times big as the first flagellomere; the first flagellomere flattened shape, the rest of flagellomere globular shape, with 36 segments of flagellomere, the last flagellomere subtriangle.
Body medium size. Pronotum short and slender, bearing dense, long and curved setae. Mesonotum and metanotum both transverse strip shape, notum slightly broader than the metanotum, covered with intensive long setae.
Femur and tarsus slender, covered with short hairs at both ends while long, mixed short hairs located in the middle part; tarsus with five segments covered with short setae; tibial spurs absent; tarsal claws slender and curved, simple; arolium present. First tarsomere of the foreleg 1.5 times as long as second tarsomere, other tarsus of midleg is 2.5 times as long as second tarsus; metafemur subcylindrical, middle part of tarsus slightly swelled.
Female genitalia simple (
Figure 5C). Tergite and sternite 1–8 both well developed and subrectangle, without modification; sternite gradually decreases from sternite 6, sternite 8 smallest and reduced to a small subtriangle; ventral side of tergite 9 bent caudally and slightly extended, thinner than tergite 8; ectoproct and 9th gonocoxites both semicircle; tergite 9 free from ectoproct, without hypocaudae.
Forewings (
Figure 6A,B,E,F) oval and rounded apex. All subcostal veinlets simple. RA forked and all branches marginally branched. Sc and R free apically, one sc-r and one sc-ra crossvein present. One ra-rp crossvein connecting RA and RP3, RP1 and RP3 simple pectinate branches and RP2 bifurcately branched at the end. One rp-m and m-cua crossvein present before origin of MP. MA and MP both lightly fork with trifurcate branch near wing margin. Two m-cua crossveins present. CuA with three short pectinate branches, CuP simple. One cup-aa1 crossvein present. AA1 with three pectinate branches; AA2 with five to seven simple pectinate branches.
Hindwings (
Figure 6C,D,G,H) elongated and round. All subcostal veinlets simple. Sc and R free at apex, connected with a crossvein. One ra-rp crossvein present; RA simple trifurcately branched at apex; RP1 and RP2 bifurcately branched; RP3 trifurcately branched. MA shallow branched with some pectinate branches near wing margin; MP trifurcately branched. CuA with six short pectinate branches near the wing edge, CuP with three or five simple pectinate branches. AA1 with a distal fork.
Sejunctaberotha transversa Chen, Shi, Ren & Yang gen. et sp. nov.
urn:lsid:zoobank.org:act:B9493266-1509-4FED-9DDF-549A596848D8
Material. Holotype: CNU-NEU-MA2018100.
Etymology. The specific epithet is from the Latin transversus (meaning “transverse”), referring to the dorsal plate of the prothorax stout, transverse rectangle, clearly wider than long.
Locality and horizon. Hukawng Valley, Kachin State, northern Myanmar; Lowermost Cenomanian, Upper Cretaceous.
Diagnosis. Antennae relatively long, with about 29 flagellomeres; protothorax short and stout, wider than length; in forewing, MA pectinately branched near the wing margin in forewing while other species are bifurcately forked and pectinately branched at apex. Protergum stout and transverse, wider than long.
Description. Body length ca. 1.6 mm. Right forewing length 2.3 mm, width 0.8 mm; left forewing length 2.2 mm, width 0.9 mm. Left hindwing length 1.7 mm, width 0.9 mm; right hindwing length 1.9 mm, width 0.8 mm.
Head (
Figure 7B). Oculus strongly prominent in hemispherical shape. Frons and mouthparts slightly elongated. Vertex elevated above the oculus, hemisphere, scattered mid-length bristles. Antennae surround with dense bristle; scape slightly elongated, about 2.5 times as long as wide; pedicel subrectangle; flagellomere subtrapezoidal, with ca. 29 flagellomeres, last flagellomere triangle, smaller than previous flagellomeres.
Pronotum transversal square, covered with scattered medium-length setae, one transversal furrow present. Mesothorax and metathorax both damaged.
Forelegs are walking legs. Femur and tarsus slender, short setae covered both at ends while middle part located with mixed of stout, long and medium-length, fine setae; short setae densely distributed in tarsomere, with tarsus 5 segmented, the first tarsomere longest, the remaining tarsal segments nearly equal in length; without tibial spurs; tarsal claws slender and curved; arolium present (
Figure 7C,D).
Abdomen poorly preserved.
Forewings (
Figure 8A,B,E,F) ovoid and rounded apex. Humeral vein simple. Sc and R free distally, connected by a crossvein, another sc-r crossvein located before origin of RP. One ra-rp3 present. RP1 and RP3 with three short pectinate branches, RP2 one bifurcated, forked branch. One rp-m crossvein present before the origin of MP; MA pectinately branched near wing margin; MP light bifurcated fork before margin branch (LFW), or with four pectinate branches (RFW). Two m-cua crossveins present; CuA trichotomous branched distally; CuP simple. One cu-aa1 crossvein present near the origin of CuP, AA1 and AA2 with three to four short pectinate branches near wing margin; AA3 simple.
Hindwings (
Figure 8C,D,G,H). Costal space narrow at wing base, maximum width and rounded at apex. All subcostal veinlets simple. Sc and R broadly separated, connected by one crossvein. RP diverged from RA far from wing base; RP with three major branches, each branch forked at apex. One short and sigmoidal rp-m crossvein present before origin of R1 and MP. MA twice forked distally; MP pectinately branched apically. CuA almost parallel to the posterior wing margin, with five pectinate branches; CuP pectinately branched near the wing margin. AA1 with three short pectinate branches close to the wing verge, AA2 and AA3 not observed.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







