
Prevalence in Potato of ‘Candidatus Arsenophonus Phytopathogenicus’ and ‘Candidatus Phytoplasma Solani’ and Their Transmission via Adult Pentastiridius leporinus

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
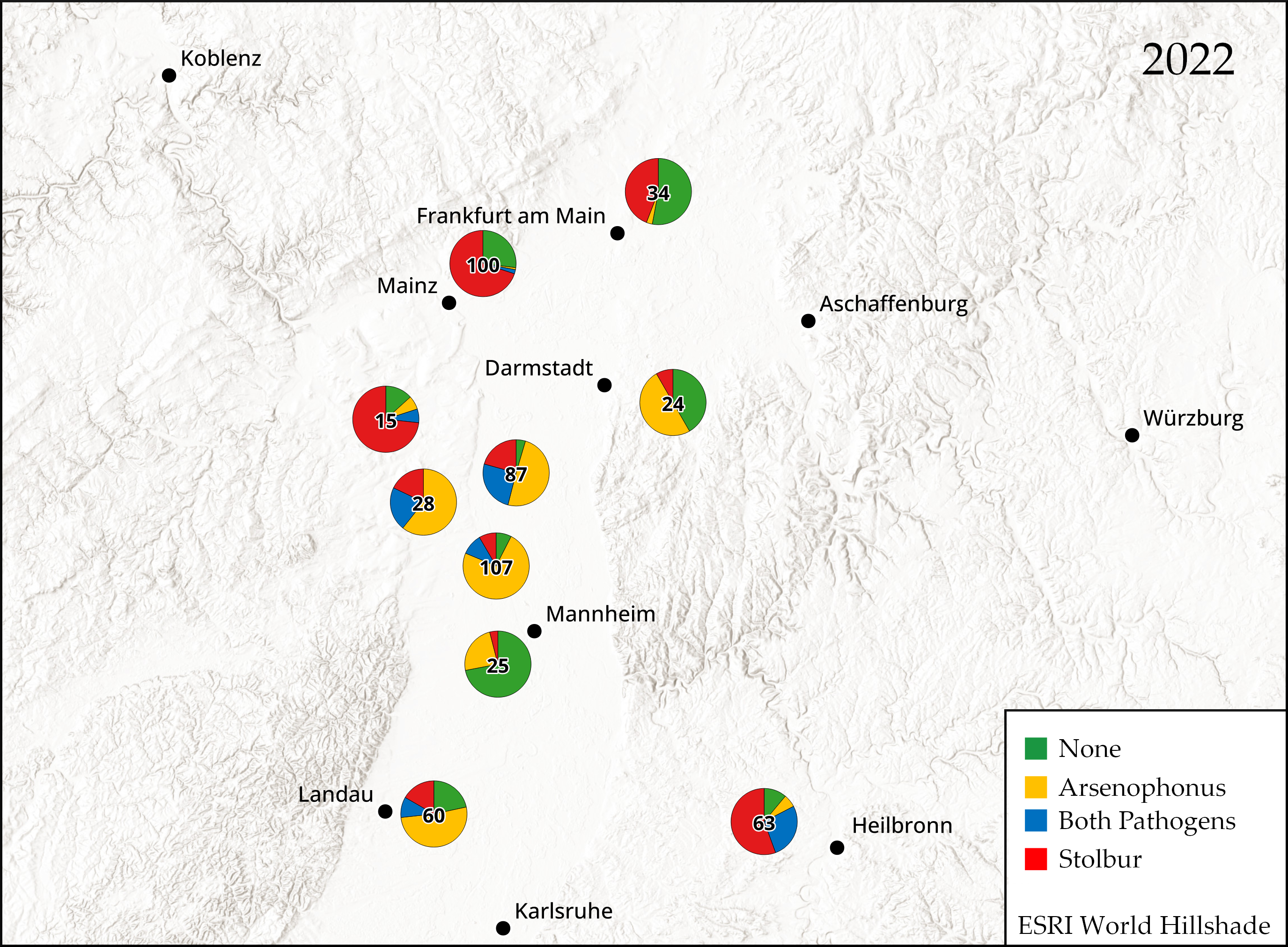
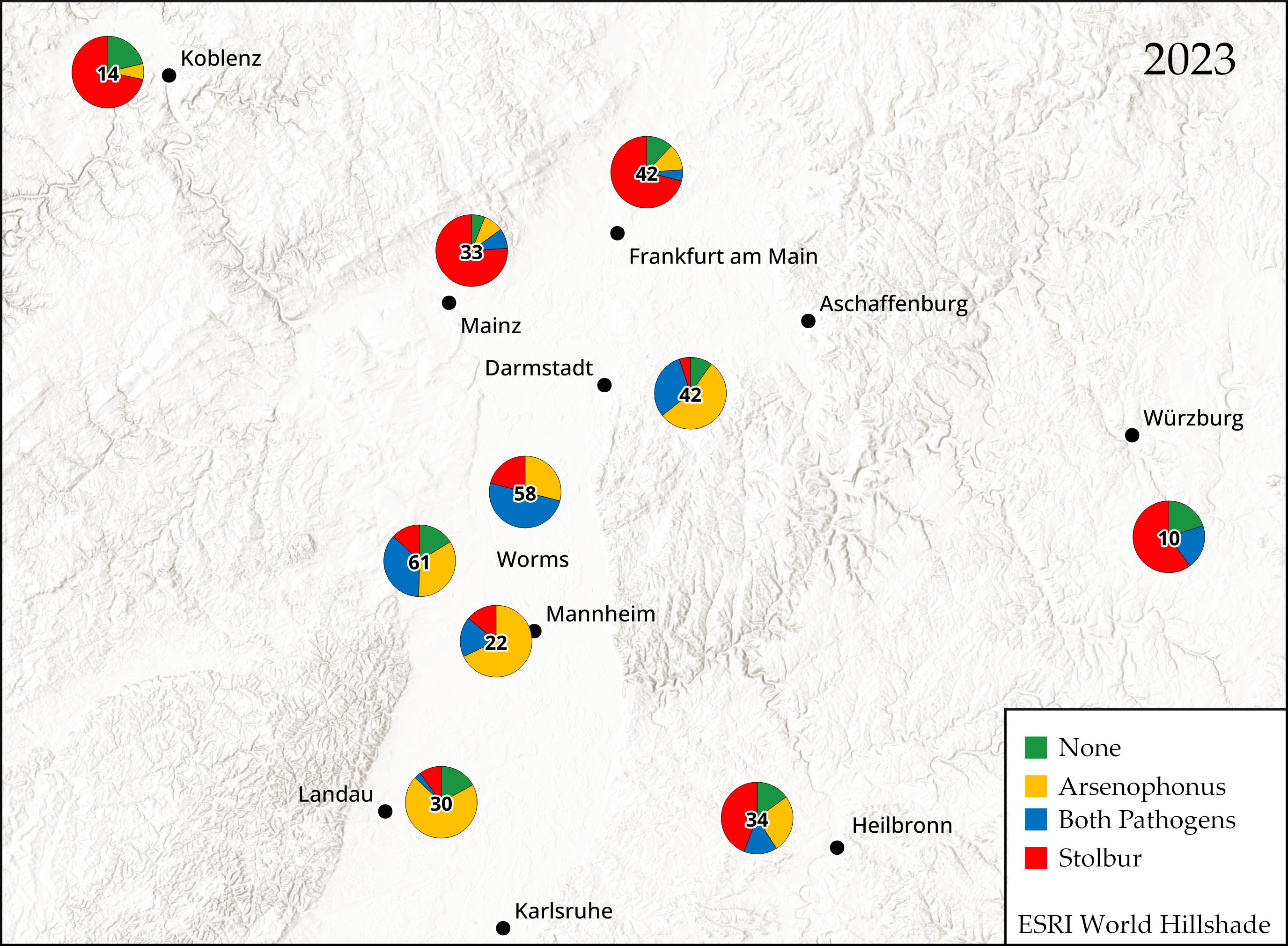
2.1. Detection of Arsenophonus and Stolbur in Potato Fields
2.2. Germination Trials
2.3. Transmission Assays
3. Results
3.1. Prevalence of Arsenophonus and Stolbur in Symptomatic Potato Tubers
3.2. Germination Assay
3.3. Transmission Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatineau, F.; Larrue, J.; Clair, D.; Lorton, F.; Richard-Molard, M.; Boudon-Padieu, E. A New Natural Planthopper Vector of Stolbur Phytoplasma in the Genus Pentastiridius (Hemiptera: Cixiidae). Eur. J. Plant Pathol. 2001, 107, 263–271. [Google Scholar] [CrossRef]
- Gatineau, F.; Jacob, N.; Vautrin, S.; Larrue, J.; Lherminier, J.; Richard-Molard, M.; Boudon-Padieu, E. Association with the Syndrome “Basses Richesses” of Sugar Beet of a Phytoplasma and a Bacterium-Like Organism Transmitted by a Pentastiridius sp. Phytopathology 2002, 92, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Sémétey, O.; Bressan, A.; Gatineau, F.; Boudon-Padieu, E. Development of a specific assay using RISA for detection of the bacterial agent of ‘basses richesses’ syndrome of sugar beet and confirmation of a Pentastiridius sp. (Fulgoromopha, Cixiidae) as the economic vector. Plant Pathol. 2007, 56, 797–804. [Google Scholar] [CrossRef]
- Bressan, A. Emergence and evolution of Arsenophonus bacteria as insect-vectored plant pathogens. Infect. Genet. Evol. 2014, 22, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Bressan, A.; Sémétey, O.; Arneodo, J.; Lherminier, J.; Boudon-Padieu, E. Vector transmission of a plant-pathogenic bacterium in the Arsenophonus clade sharing ecological traits with facultative insect endosymbionts. Phytopathology 2009, 99, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Bressan, A.; Sémétey, O.; Nusillard, B.; Clair, D.; Boudon-Padieu, E. Insect Vectors (Hemiptera: Cixiidae) and Pathogens Associated with the Disease Syndrome “Basses Richesses” of Sugar Beet in France. Plant Dis. 2008, 92, 113–119. [Google Scholar] [CrossRef]
- Ćurčić, Ž.; Stepanović, J.; Zübert, C.; Taški-Ajduković, K.; Kosovac, A.; Rekanović, E.; Kube, M.; Duduk, B. Rubbery Taproot Disease of Sugar Beet in Serbia Associated with ‘Candidatus Phytoplasma solani’. Plant Dis. 2021, 105, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Duduk, B.; Ćurčić, Ž.; Stepanović, J.; Böhm, J.W.; Kosovac, A.; Rekanović, E.; Kube, M. Prevalence of a ‘Candidatus Phytoplasma solani’-Related Strain Designated as New 16SrXII-P Subgroup over ‘Candidatus Arsenophonus phytopathogenicus’ in Sugar Beet in Eastern Germany. Plant Dis. 2023, 107, 3792–3800. [Google Scholar] [CrossRef] [PubMed]
- Bressan, A.; Holzinger, W.E.; Nusillard, B.; Sémétey, O.; Gatineau, F.; Simonato, M.; Boudon-Padieu, E. Identification and biological traits of a planthopper from the genus Pentastiridius (Hemiptera: Cixiidae) adapted to an annual cropping rotation. Eur. J. Entomol. 2009, 106, 405–413. [Google Scholar] [CrossRef]
- Kosovac, A.; Ćurčić, Ž.; Stepanović, J.; Rekanović, E.; Duduk, B. Epidemiological role of novel and already known ‘Ca. P. solani’ cixiid vectors in rubbery taproot disease of sugar beet in Serbia. Sci. Rep. 2023, 13, 1433. [Google Scholar] [CrossRef]
- Emeljanov, A.F. Nomenclatorial changes in the family Cixiidae (Homoptera, Auchenorrhyncha, Fulgoroidea), with fixation of type species of the genus Reptalus Emeljanov, 1971 and description of a new subgenus. Zootaxa 2020, 4780, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Behrmann, S.; Schwind, M.; Schieler, M.; Vilcinskas, A.; Martinez, O.; Lee, K.-Z.; Lang, C. Spread of bacterial and virus yellowing diseases of sugar beet in South and Central Germany from 2017–2020. Sugar Ind. 2021, 146, 476–485. [Google Scholar] [CrossRef]
- Mahillon, M.; Groux, R.; Bussereau, F.; Brodard, J.; Debonneville, C.; Demal, S.; Kellenberger, I.; Peter, M.; Steinger, T.; Schumpp, O. Virus Yellows and Syndrome “Basses Richesses” in Western Switzerland: A Dramatic 2020 Season Calls for Urgent Control Measures. Pathogens 2022, 11, 885. [Google Scholar] [CrossRef] [PubMed]
- Jović, J.; Ember, I.; Mitrović, M.; Cvrković, T.; Krstić, O.; Krnjajić, S.; Acs, Z.; Kölber, M.; Toševski, I. Molecular detection of potato stolbur phytoplasma in Serbia. Bull. Insectol. 2011, 64, 83–84. [Google Scholar]
- Mitrović, M.; Trivellone, V.; Jović, J.; Cvrković, T.; Jakovljevic, M.; Kosovac, A.; Krstic, O.; Toševski, I. Potential Hemipteran vectors of “stolbur” phytoplasma in potato fields in Serbia. Phyt. Moll. 2015, 5, S49. [Google Scholar] [CrossRef]
- Mitrović, M.; Jakovljević, M.; Jović, J.; Krstić, O.; Kosovac, A.; Trivellone, V.; Jermini, M.; Toševski, I.; Cvrković, T. ‘Candidatus phytoplasma solani’ genotypes associated with potato stolbur in Serbia and the role of Hyalesthes obsoletus and Reptalus panzeri (hemiptera, cixiidae) as natural vectors. Eur. J. Plant Pathol. 2016, 144, 619–630. [Google Scholar] [CrossRef]
- Behrmann, S.C.; Rinklef, A.; Lang, C.; Vilcinskas, A.; Lee, K.-Z. Potato (Solanum tuberosum) as a New Host for Pentastiridius leporinus (Hemiptera: Cixiidae) and Candidatus Arsenophonus Phytopathogenicus. Insects 2023, 14, 281. [Google Scholar] [CrossRef] [PubMed]
- Therhaag, E.; Schneider, B.; Zikeli, K.; Maixner, M.; Gross, J. Pentastiridius leporinus (Linnaeus, 1761) as a Vector of Phloem-Restricted Pathogens on Potatoes: ‘Candidatus Arsenophonus Phytopathogenicus’ and ‘Candidatus Phytoplasma Solani’. Insects 2024, 15, 189. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Behrmann, S.C.; Witczak, N.; Lang, C.; Schieler, M.; Dettweiler, A.; Kleinhenz, B.; Schwind, M.; Vilcinskas, A.; Lee, K.-Z. Biology and Rearing of an Emerging Sugar Beet Pest: The Planthopper Pentastiridius leporinus. Insects 2022, 13, 656. [Google Scholar] [CrossRef]
- Biedermann, R.; Niedringhaus, R. Die Zikaden Deutschlands: Bestimmungstafeln für alle Arten; Wissenschaftlich-Akademischer-Buchvertrieb: Osnarbrueck, Germany, 2004; ISBN 978-3-939202-00-4. [Google Scholar]
- Bressan, A.; Moral García, F.J.; Boudon-Padieu, E. The prevalence of ‘Candidatus Arsenophonus phytopathogenicus’ infecting the planthopper Pentastiridius leporinus (Hemiptera: Cixiidae) increase nonlinearly with the population abundance in sugar beet fields. Environ. Entomol. 2011, 40, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Bundesanstalt für Landwirtschaft und Ernährung. Bericht zur Markt- und Versorgungslage Zucker; Federal Office for Agriculture and Food: Bonn, Germany, 2023. [Google Scholar]
- Ćurčić, Ž.; Kosovac, A.; Stepanović, J.; Rekanović, E.; Kube, M.; Duduk, B. Multilocus Genotyping of ‘Candidatus Phytoplasma solani’ Associated with Rubbery Taproot Disease of Sugar Beet in the Pannonian Plain. Microorganisms 2021, 9, 1950. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, H.; Wodicka, B. On the etiology of the occurrence of callose in the tubers of stolbur-diseased potato plants. J. Plant Dis. Prot. 1981, 88, 584–587. [Google Scholar]
- Santi, S.; de Marco, F.; Polizzotto, R.; Grisan, S.; Musetti, R. Recovery from stolbur disease in grapevine involves changes in sugar transport and metabolism. Front. Plant Sci. 2013, 4, 171. [Google Scholar] [CrossRef] [PubMed]
- Paltrinieri, S.; Bertaccini, A. Detection of phytoplasmas in plantlets grown from different batches of seed-potatoes. Bull. Insectol. 2007, 60, 379–380. [Google Scholar]
- Bertaccini, A.; Duduk, B. Phytoplasma and phytoplasma diseases: A review of recent research. Phytopathol. Mediterr. 2009, 48, 355–378. [Google Scholar]
- Christensen, N.M.; Nicolaisen, M.; Hansen, M.; Schulz, A. Distribution of phytoplasmas in infected plants as revealed by real-time PCR and bioimaging. Mol. Plant Microbe Interact. 2004, 17, 1175–1184. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| None | Arsenophonus | Stolbur | Both | |
|---|---|---|---|---|
| Germinated plants | 2/33 | 7/97 | 14/101 | 3/67 |
| Germination rate | 93.9% | 92.8% | 86.1% | 95.5% |
| Arsenophonus | Stolbur | |||
|---|---|---|---|---|
| Negative | Positive | Negative | Positive | |
| Infected stems | 0/26 | 0/33 | 0/22 | 2/37 |
| Infection rate | 0% | 0% | 0% | 5.4% |
| Median copies | - | - | - | 45 |
| Plant Species | Arsenophonus | Stolbur | ||
|---|---|---|---|---|
| Detection Rate | Median Copy Number | Detection Rate | Median Copy Number | |
| Potato | 65% (13/20) | 25% (5/20) | ||
| - Leaf | 10% (2/20) | 10,133 | 0% (0/20) | - |
| - Upper shoot | 35% (7/20) | 170 | 5% (1/20) | 45 |
| - Lower shoot | 35% (7/20) | 958 | 10% (2/20) | 6210 |
| - Roots | 30% (6/20) | 86 | 20% (4/20) | 187 |
| Sugar beet | 83% (5/6) | 50% (3/6) | ||
| - Leaf | 33% (2/6) | 129 | 17% (1/6) | 5918 |
| - Lower stem | 83% (5/6) | 2684 | 50% (3/6) | 182,076 |
| - Beetroot tip | 83% (5/6) | 5742 | 50% (3/6) | 78,061 |
| Potato (control) | 0% (0/6) | 0% (0/6) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinklef, A.; Behrmann, S.C.; Löffler, D.; Erner, J.; Meyer, M.V.; Lang, C.; Vilcinskas, A.; Lee, K.-Z. Prevalence in Potato of ‘Candidatus Arsenophonus Phytopathogenicus’ and ‘Candidatus Phytoplasma Solani’ and Their Transmission via Adult Pentastiridius leporinus. Insects 2024, 15, 275. https://doi.org/10.3390/insects15040275
Rinklef A, Behrmann SC, Löffler D, Erner J, Meyer MV, Lang C, Vilcinskas A, Lee K-Z. Prevalence in Potato of ‘Candidatus Arsenophonus Phytopathogenicus’ and ‘Candidatus Phytoplasma Solani’ and Their Transmission via Adult Pentastiridius leporinus. Insects. 2024; 15(4):275. https://doi.org/10.3390/insects15040275
Chicago/Turabian StyleRinklef, André, Sarah Christin Behrmann, David Löffler, Jan Erner, Martin Vincent Meyer, Christian Lang, Andreas Vilcinskas, and Kwang-Zin Lee. 2024. "Prevalence in Potato of ‘Candidatus Arsenophonus Phytopathogenicus’ and ‘Candidatus Phytoplasma Solani’ and Their Transmission via Adult Pentastiridius leporinus" Insects 15, no. 4: 275. https://doi.org/10.3390/insects15040275