

Point Protection with Transfluthrin against Musca domestica L. in a Semi-Field Enclosure
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing
2.2. Permethrin Susceptibility Bioassay
2.3. Semi-Field Experimental Design
Transfluthrin and Control Traps
2.4. Statistical Analysis and Data Visualization
3. Results
3.1. Permethrin Susceptibility Bioassay
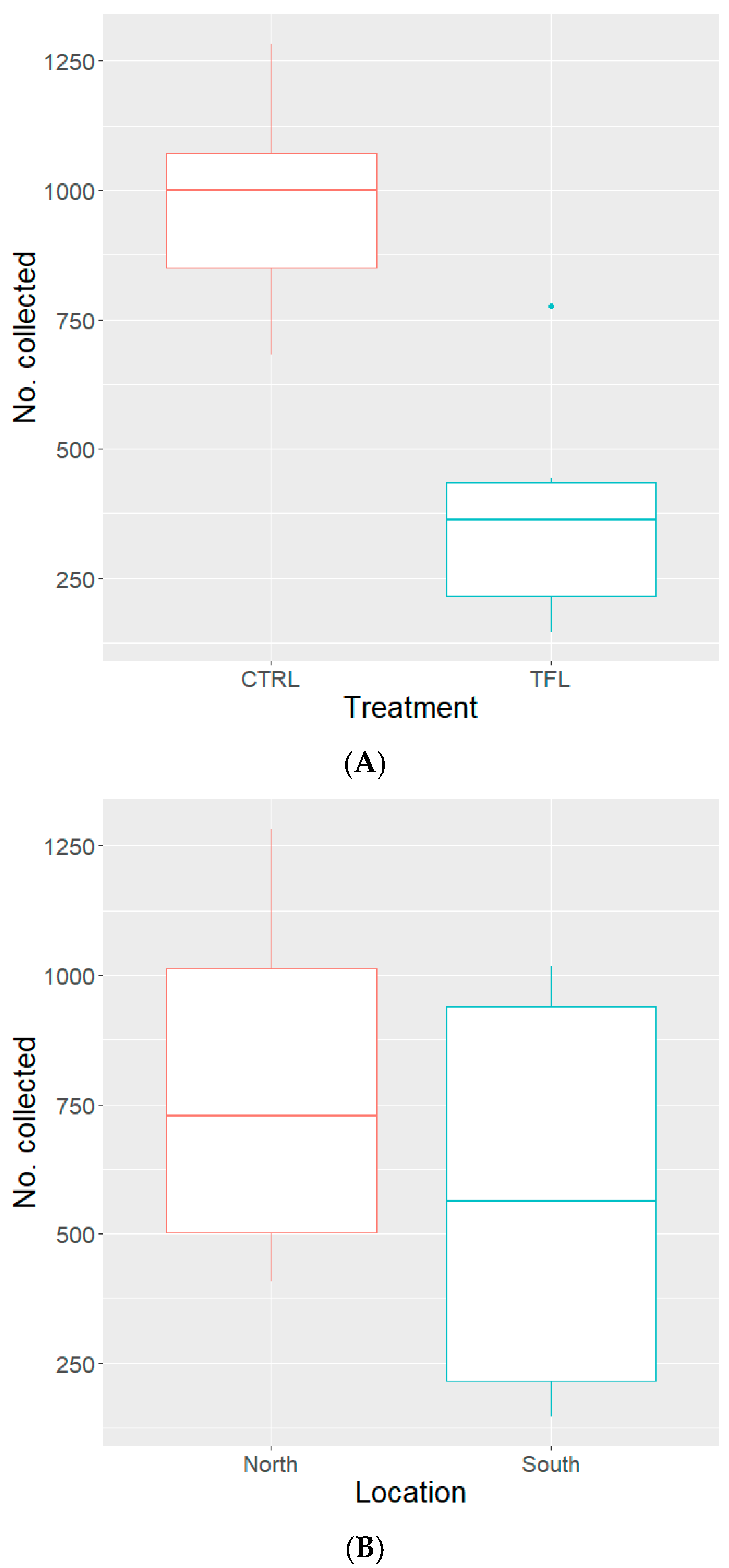
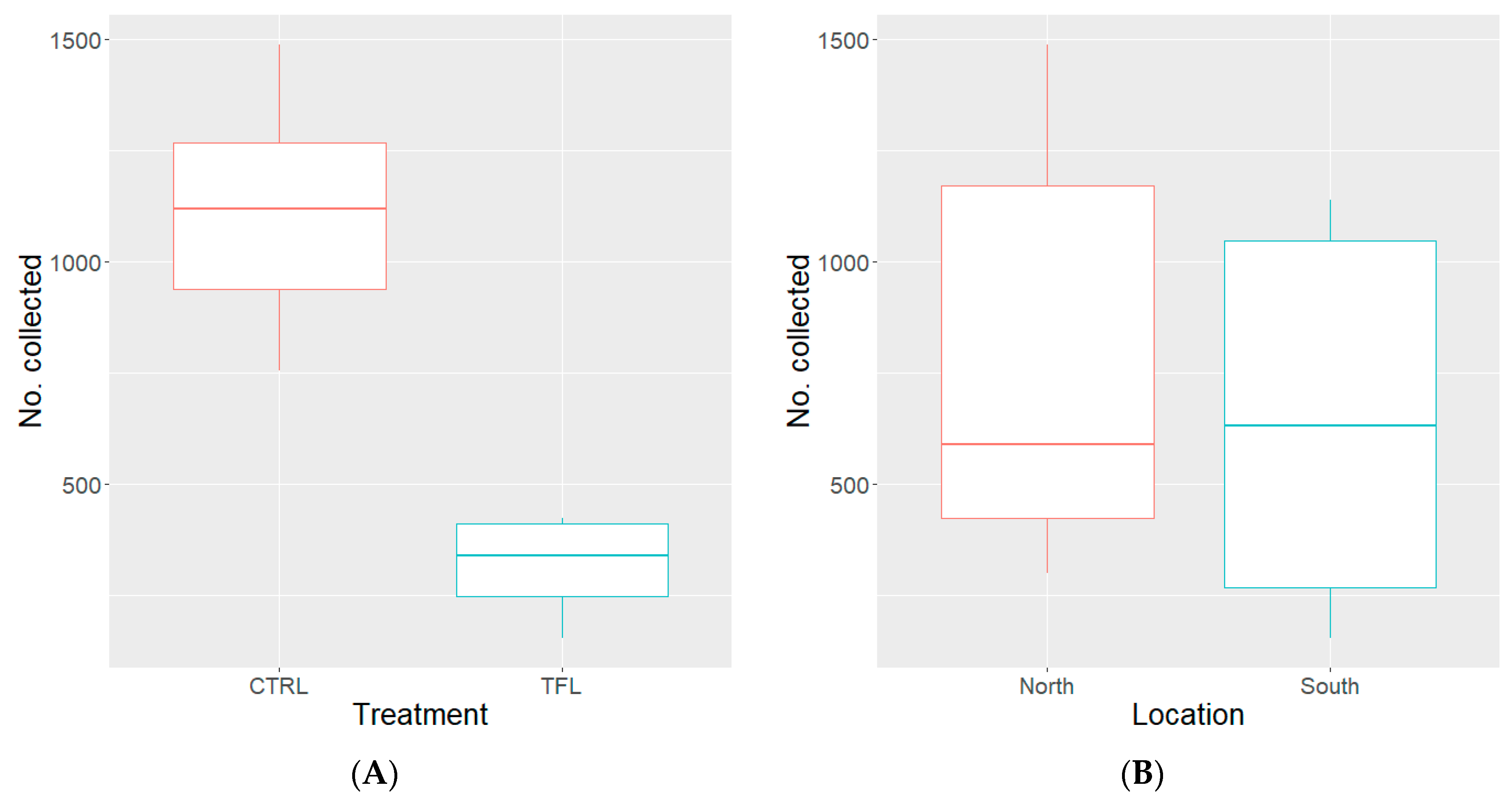
3.2. Semi-Field with “WHF” Permethrin Resistant Strain
3.3. Semi-Field with “CAR21” Permethrin Susceptible Strain
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khamesipour, F.; Lankarani, K.B.; Honarvar, B.; Kwenti, T.E. A Systematic Review of Human Pathogens Carried by the Housefly (Musca domestica L.). BMC Public Health 2018, 18, 1049. [Google Scholar] [CrossRef] [PubMed]
- Tsagaan, A.; Kanuka, I.; Okado, K. Study of Pathogenic Bacteria Detected in Fly Samples Using Universal Primer-Multiplex PCR. Mong. J. Agric. Sci. 2015, 15, 27–32. [Google Scholar] [CrossRef]
- Szalanski, A.L.; Owens, C.B.; McKay, T.; Steelman, C.D. Detection of Campylobacter and Escherichia coli O157:H7 from Filth Flies by Polymerase Chain Reaction. Med. Vet. Entomol. 2004, 18, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Smallegange, R.C.; den Otter, C.J. Houseflies, Annoying and Dangerous. In Emerging Pests and Vector-Borne Diseases in Europe; Bart, G.J.K., Willem, T., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; Volume 1, p. 500. ISBN 9789086860531. [Google Scholar]
- Malik, A.; Singh, N.; Satya, S. House Fly (Musca domestica): A Review of Control Strategies for a Challenging Pest. J. Environ. Sci. Health B 2007, 42, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Ware, G.W.; Whitacre, D. The Pesticide Book, 5th ed.; Thomson Purblications: Fresno, CA, USA, 2000. [Google Scholar]
- Britch, S.C.; Linthicum, K.J.; Kline, D.L.; Aldridge, R.L.; Golden, F.V.; Wittie, J.; Henke, J.; Hung, K.; Gutierrez, A.; Snelling, M.; et al. Transfluthrin Spatial Repellent on US Military Materials Reduces Culex tarsalis Incursion in a Desert Environment. J. Am. Mosq. Control Assoc. 2020, 36, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Scrivener, A.M.; Peters, B.A.; Miller, P.F. Efficacy of a Novel Area-Repellent Aerosol Formulation against Mosquitoes and Flies (Diptera: Culicidae, Muscidae). In Proceedings of the 8th International Conference on Urban Pests, Zurich, Switzerland, 20–23 July 2014; pp. 153–161. [Google Scholar]
- Morrison, A.C.; Reiner, R.C., Jr.; Elson, W.H.; Astete, H.; Guevara, C.; Del Aguila, C.; Bazan, I.; Siles, C.; Barrera, P.; Kawiecki, A.B.; et al. Efficacy of a Spatial Repellent for Control of Aedes-Borne Virus Transmission: A Cluster-Randomized Trial in Iquitos, Peru. Proc. Natl. Acad. Sci. USA 2022, 119, e2118283119. [Google Scholar] [CrossRef] [PubMed]
- Field, L.M.; Emyr Davies, T.G.; O’Reilly, A.O.; Williamson, M.S.; Wallace, B.A. Voltage-Gated Sodium Channels as Targets for Pyrethroid Insecticides. Eur. Biophys. J. 2017, 46, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Baker, O.S.; Norris, E.J.; Burgess, E.R., IV. Insecticidal and Synergistic Potential of Three Monoterpenoids against the Yellow Fever Mosquito, Aedes aegypti (Diptera: Culicidae), and the House Fly, Musca domestica (Diptera: Muscidae). Molecules 2023, 28, 3250. [Google Scholar] [CrossRef] [PubMed]
- Geden, C.J.; Johnson, D.M.; Taylor, D.B. Improved Sentinel Method for Surveillance and Collection of Filth Fly Parasitoids. J. Insect Sci. 2020, 20, 3. [Google Scholar] [CrossRef]
- Taylor, E.E.; Barguez-Arias, N.N.; Herrera, R.; King, B.H.; Burgess, E.R., IV. Sweeteners Allulose and Neotame for Potential Use in House Fly Baits. J. Appl. Entomol. 2023, 147, 790–797. [Google Scholar] [CrossRef]
- Warner, W.B. Attractant Composition for Synanthropic Flies. US Patent 5008107, 16 April 1991. [Google Scholar]
- Britch, S.C.; Kline, D.L.; Linthicum, K.J.; Urban, J.; Dickstein, E.; Aldridge, R.L.; Golden, F.V. Transfluthrin Spatial Repellent on US Military Camouflage Netting Reduces Tabanids in a Warm-Temperate Environment. J. Am. Mosq. Control Assoc. 2020, 36, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Estep, A.S.; Sanscrainte, N.D.; Pagac, A.A.; Geden, C.J.; Burgess, E.R., IV. USDA-ARS-CMAVE; Gainesville, FL, USA, 2024; (manuscript in preparation). [Google Scholar]
- Sun, H.; Tong, K.P.; Kasai, S.; Scott, J.G. Overcoming Super-Knock down Resistance (Super-Kdr) Mediated Resistance: Multi-Halogenated Benzyl Pyrethroids Are More Toxic to Super-Kdr than Kdr House Flies. Insect Mol. Biol. 2016, 25, 126–137. [Google Scholar] [CrossRef]
- Scott, J.G.; Wen, Z. Toxicity of Fipronil to Susceptible and Resistant Strains of German Cockroaches (Dictyoptera: Blattellidae) and House Flies (Diptera: Muscidae). J. Econ. Entomol. 1997, 90, 1152–1156. [Google Scholar] [CrossRef]
- Busvine, J.R. Mechanism of Resistance to Insecticide in Houseflies. Nature 1951, 168, 193–195. [Google Scholar] [CrossRef]
- Liu, N.; Scott, J.G. Increased Transcription of CYP6D1 Causes Cytochrome P450-Mediated Insecticide Resistance in House Fly. Insect Biochem. Mol. Biol. 1998, 28, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yue, X. Insecticide Resistance and Cross-Resistance in the House Fly (Diptera: Muscidae). J. Econ. Entomol. 2000, 93, 1269–1275. [Google Scholar] [CrossRef]
- Goodenough, J.L.; Wilson, D.D.; Agee, H.R. Electroretinographic Measurements for Comparison of Visual Sensitivity of Wild and Mass-Reared Screwworm Flies, Cochliomyia hominivorax (Diptera: Calliphoridae)1. J. Med. Entomol. 1977, 14, 309–312. [Google Scholar] [CrossRef]
- Robinson, A.; de Oliveira Gomes, L.R.; Abdurahman, O.S.; Alemayehu, W.; Shuka, G.; Melese, E.; Guye, M.; Legesse, D.; Elias, E.; Temam, K.; et al. Evaluation of the Efficacy of Insecticide-Treated Scarves to Protect Children from the Trachoma Vector Musca sorbens (Diptera: Muscidae): A Phase II Randomised Controlled Trial in Oromia, Ethiopia. eClinicalMedicine 2022, 49, 101487. [Google Scholar] [CrossRef] [PubMed]
- Khater, H.F.; Geden, C.J. Efficacy and Repellency of Some Essential Oils and Their Blends against Larval and Adult House Flies, Musca domestica L. (Diptera: Muscidae). J. Vector Ecol. 2019, 44, 256–263. [Google Scholar] [CrossRef]
- Hazarika, H.; Tyagi, V.; Krishnatreyya, H.; Islam, J.; Boruah, D.; Kishor, S.; Chattopadhyay, P.; Zaman, K. Essential Oil Based Controlled-Release Non-Toxic Evaporating Tablet Provides Effective Repellency against Musca domestica. Acta Trop. 2020, 210, 105620. [Google Scholar] [CrossRef]
- Andreazza, F.; Valbon, W.; Dong, K. Transfluthrin Enhances Odorant Receptor-Mediated Spatial Repellency in Aedes aegypti. Pestic. Biochem. Physiol. 2023, 192, 105387. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Fly Strain | N | Slope (SE) | LD50 (95% CL) (ng/fly) | LD90 (95% CL) (ng/fly) | Resistance Ratio LD50 (RR) | Resistance Ratio LD90 (RR) |
|---|---|---|---|---|---|---|
| CAR21 | 400 | 1.56 (0.13) | 11 (10–13) | 26 (22–32) | -- | -- |
| WHF | 400 | 0.80 (0.08) | 1720 (1149–2073) | 8534 (6134–13,642) | 149.6 | 328.2 |
| Response Var. | Treatment | Param. Est. | S.E. | Z | p |
|---|---|---|---|---|---|
| Collected ‘WHF’ house flies | Intercept | 6.93 | 0.15 | 46.47 | <0.0001 |
| Treatment | −0.63 | 0.21 | −2.98 | 0.003 | |
| Location | −0.09 | 0.21 | −0.41 | 0.69 | |
| Treatment × Location | −0.84 | 0.30 | −2.78 | 0.005 | |
| Collected ‘CAR21’ house flies | Intercept | 7.08 | 0.14 | 49.50 | <0.0001 |
| Treatment | −1.13 | 0.20 | −5.56 | <0.0001 | |
| Location | −0.13 | 0.20 | −0.63 | 0.53 | |
| Treatment × Location | −0.28 | 0.29 | −0.96 | 0.337 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldridge, R.L.; Pagac, A.A.; Norris, E.J.; Kline, D.L.; Geden, C.J.; Linthicum, K.J. Point Protection with Transfluthrin against Musca domestica L. in a Semi-Field Enclosure. Insects 2024, 15, 277. https://doi.org/10.3390/insects15040277
Aldridge RL, Pagac AA, Norris EJ, Kline DL, Geden CJ, Linthicum KJ. Point Protection with Transfluthrin against Musca domestica L. in a Semi-Field Enclosure. Insects. 2024; 15(4):277. https://doi.org/10.3390/insects15040277
Chicago/Turabian StyleAldridge, Robert L., Alexandra A. Pagac, Edmund J. Norris, Daniel L. Kline, Christopher J. Geden, and Kenneth J. Linthicum. 2024. "Point Protection with Transfluthrin against Musca domestica L. in a Semi-Field Enclosure" Insects 15, no. 4: 277. https://doi.org/10.3390/insects15040277