
Effect of High Temperature on Abamectin and Thiamethoxam Tolerance in Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae)

 ,
, 
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Isolation and Cloning Experiments
2.3. Insecticide Bioassays
2.4. Temperature Treatments
2.5. Combined Temperature and Insecticide Treatment
2.6. RNA Interference
2.7. Quantitative Real-Time PCR
2.8. Data Analysis
3. Results
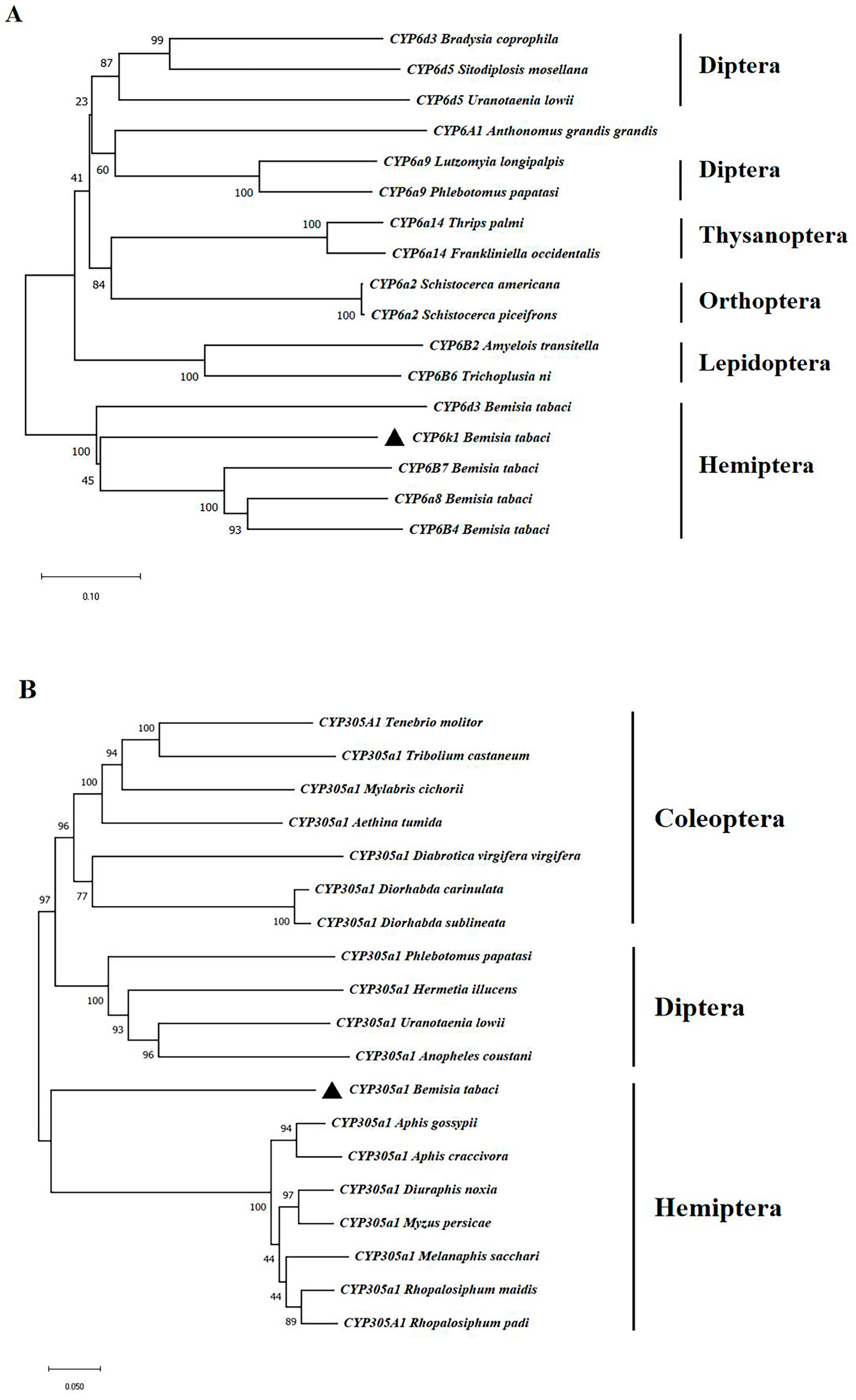
3.1. Sequence and Phylogenetic Analysis of P450s in B. tabaci MEAM1
3.2. Toxicity Assessments of Two Insecticides
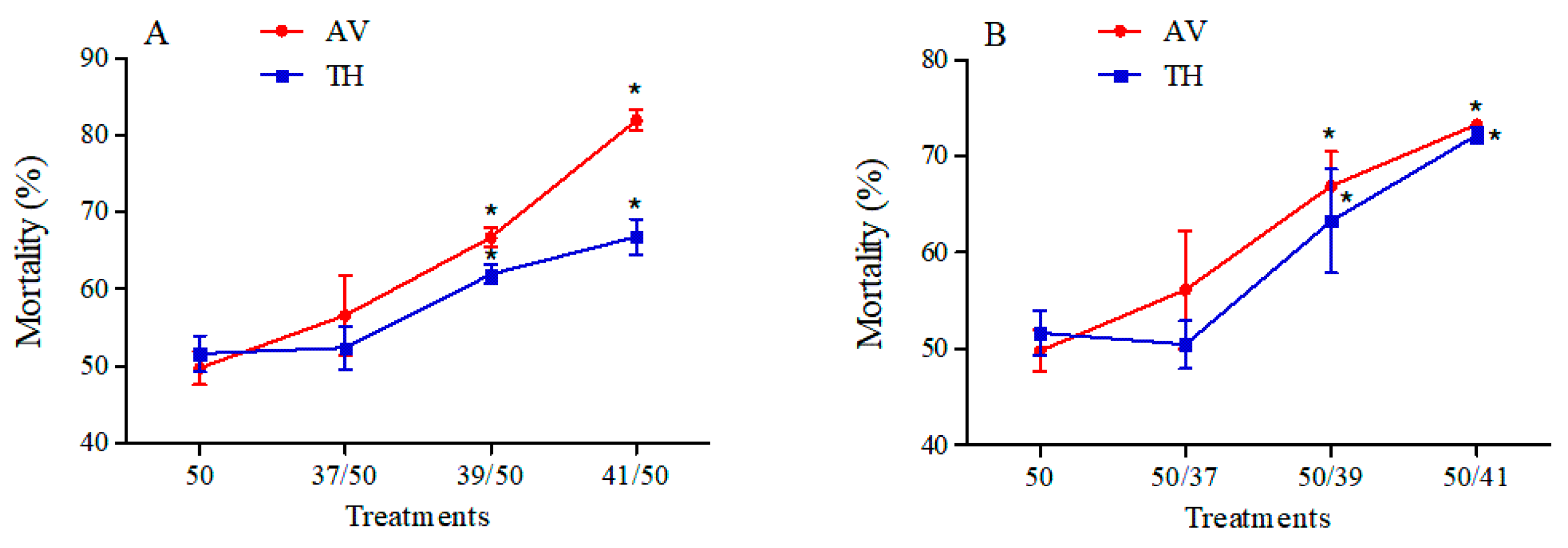
3.3. Mortality of B. tabaci MEAM1 in Response to High Temperature and Insecticide Exposure
3.4. Expression of BtCYP450s in Response to Thermal Stress
3.5. Expression of BtCYP450s in Response to Insecticide Stress
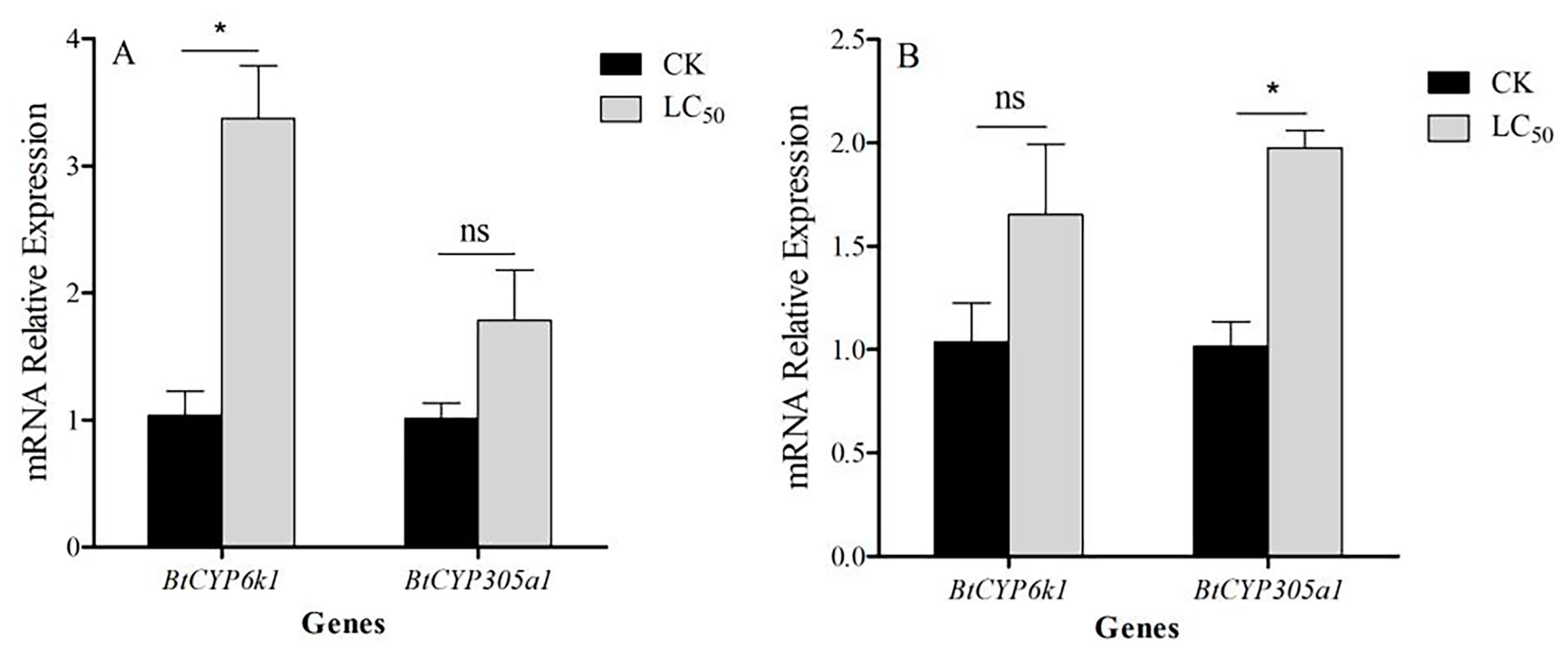
3.6. Expression of BtCYP450s in Response to Combined Stress of High Temperature and Insecticide
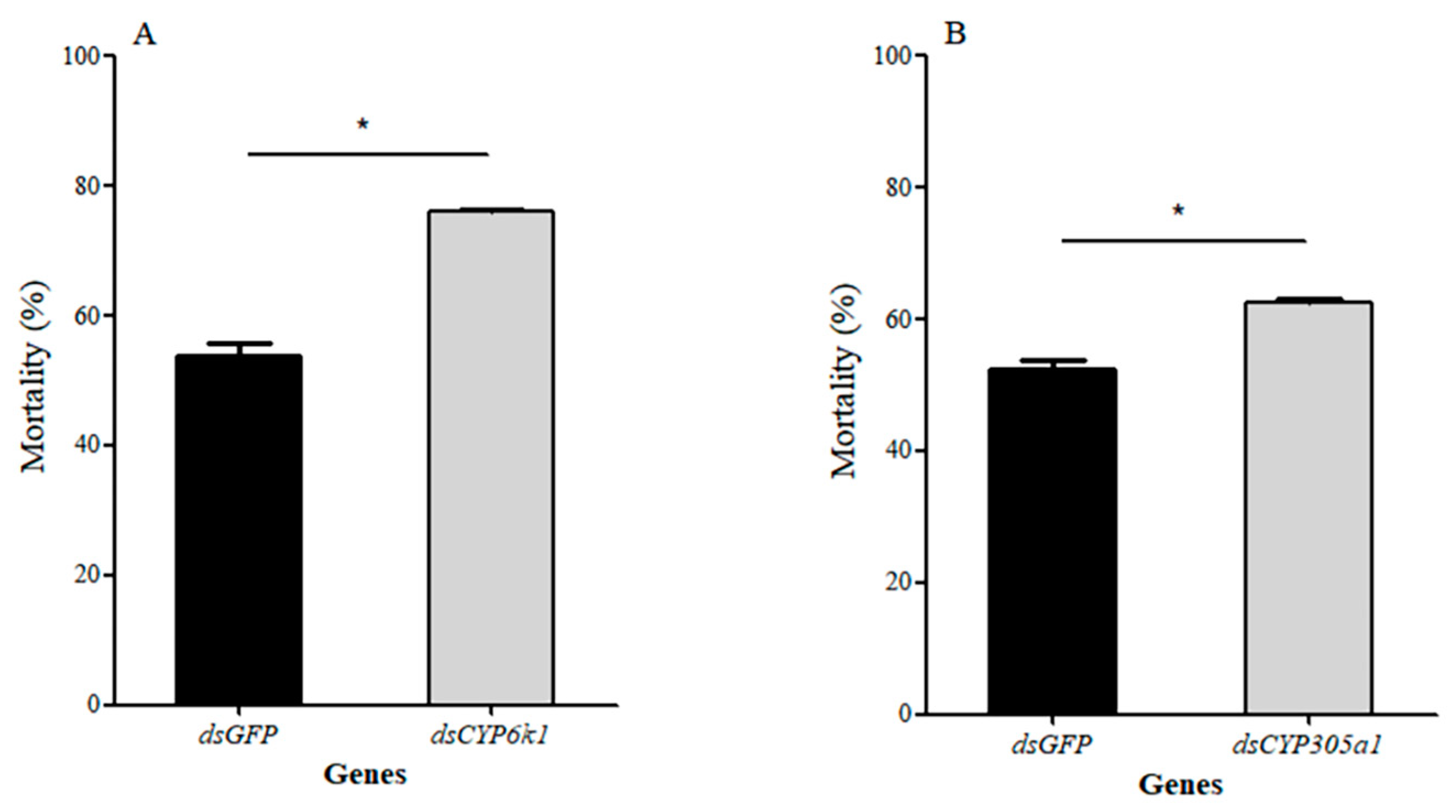
3.7. Effect of RNAi on Tolerance to Insecticides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, W.L.; Yin, F.; Chen, H.Y.; Li, Z.Y.; Feng, X.; Lin, Q.S. Comparison of resistance of Bemisia tabaci adults to six insecticides under different tillage regimes. J. Anhui Agric. Sci. 2019, 47, 158–160. [Google Scholar]
- Zhao, Q.N.; Huang, M.J.; Wei, X.G.; Yang, J.; Du, T.H.; Yin, C.; Xiang, W.S.; Yang, X.; Zhang, Y.J. Cloning of cytochrome CYP6EM1 gene in B. tabaci and its effect on imidacloprid resistance. Plant Prot. 2021, 47, 171–177. [Google Scholar]
- Kanakala, S.; Ghanim, M. Global genetic diversity and geographical distribution of Bemisia tabaci and its bacterial endosymbionts. PLoS ONE 2019, 14, e0213946. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.P.; Chu, D.; Ge, D.Q.; Wang, S.L.; Wu, Q.J.; Xie, W.; Jiao, X.G.; Liu, B.M.; Yang, X.; Yang, N.N.; et al. Further spread of and domination by Bemisia tabaci (Hemiptera: Aleyrodidae) biotype Q on field crops in China. J. Econ. Entomol. 2011, 104, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, W.; Wang, S.L.; Wu, Q.J.; Pan, H.P.; Li, R.M.; Yang, N.N.; Liu, B.M.; Xu, B.Y.; Zhou, X.M.; et al. Two cytochrome P450 genes are involved in imidacloprid resistance in field poputations of the whitefly, Bemisia tabaci, in China. Pestic. Biochem. Physiol. 2013, 107, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Berge, J.; Feyereisen, R.; Amichot, M. Cytochrome P450 monooxygenases and insecticide resistance in insects. Philos. T. R. Soc. B 1998, 353, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G. Cytochrome P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Argentine, J.A.; Claric, J.M.; Lin, H. Genetics and biochemical mechanisms of abamectin resistance in two isogenic strains of Colorado potato beetle. Pestic. Biochem. Physiol. 1992, 44, 191–207. [Google Scholar] [CrossRef]
- Nauen, R.; Stumpf, N.; Elbert, A. Toxocohigical and mechanistic studies on neonicotinoid cross resistance in Q-type Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2005, 8, 868–875. [Google Scholar]
- Karunker, I.; Benting, J.; Lueke, B.; Ponge, T.; Nauen, R.; Roditakis, E.; Vontas, J.; Gorman, K.; Denholm, I.; Morin, S. Over-expression of cytochrome P450 CYP6CM1 is associated with high resistance to imidacloprid in the B and Q biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Mol. Biol. 2008, 38, 634–644. [Google Scholar] [CrossRef]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Bautista, M.A.M.; Miyata, T.; Miura, K.; Tanakaet, T. RNA interference-mediated knockdown of a cytochrome P450, CYP6BG1, from the diamondback moth, Plutella xylostella, reduces larval resistance to permethrin. Insect Biochem. Mol. Biol. 2009, 39, 38–46. [Google Scholar] [CrossRef]
- Luo, D.; Xu, C.F.; Yin, L.X.; Zhang, S.Y.; Liu, X.P. Effects of ecological factors on mating and reproduction of moth insects. J. Environ. Entomol. 2017, 39, 963–973. [Google Scholar]
- Gullan, P.J.; Cranston, P.S.; Archer, M. The Insects: An Outline of Entomology, 3rd ed.; Blackwell Publishing Publisher: Oxford, UK, 2005; Volume 91, pp. 245–254. [Google Scholar]
- Wu, G.; Jiang, S.; Miyata, T. Seasonal Changes of Methamidophos Susceptibility and Biochemical Properties in Plutella xylostella (Lepidoptera: Yponomeutidae) and Its Parasitoid Cotesia plutellae (Hymenoptera: Braconidae). J. Econ. Entomol. 2004, 97, 1689–1698. [Google Scholar] [CrossRef]
- Feng, H.Z.; Liu, Y.H.; He, L.; Li, M.; Lu, R.E. Cross tolerance of carmine spider mite, Tetranychus cinnabarinus, to abamectin and high temperature. J. ZheJiang Univ. (Agric. Life Sci.) 2010, 36, 159–167. [Google Scholar]
- Mansour, S.I. Influence of temperature on the toxicity and stability of insecticide resistance against Spodoptera littoralis (Lepidoptera: Noctuidae). Bull. Natl. Res. Cent. 2023, 47, 85. [Google Scholar]
- Guo, L.; Su, M.M.; Liang, P.; Li, S.; Chu, D. Effects of high temperature on insecticide tolerance in whitefly Bemisia tabaci (Gennadius) Q biotype. Pestic. Biochem. Physiol. 2018, 150, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Hong, I.J.; Kim, A.Y.; Phuong, N.; Ho, K.D.; Ho, K.Y. Temperature-independent increase in the detoxifying enzyme activity of insecticide-resistant small brown planthoppers and Drosophila. J. Asia-Pac. Entomol. 2021, 24, 70–76. [Google Scholar]
- Xiao, N.; Pan, L.L.; Zhang, C.R.; Shan, H.W.; Liu, S.S. Differential tolerance capacity to unfavourable low and high temperatures between two invasive whiteflies. Sci. Rep. 2016, 6, 24306. [Google Scholar] [CrossRef]
- Liu, X.N. Hidden Species Monitoring of Tobacco Whiteflies and Transcriptome Analysis of MED Hidden Species under High and Low Temperature Stress. Master’s Thesis, Yangzhou University, Yangzhou, China, 2017. [Google Scholar]
- Bai, J. Comparative transcriptome analysis of two Bemisia tabaci cryptospecies under temperature and insecticide stress. 2024, Unpublished manuscript. Last modified 1 May 2024. Microsoft Word file.
- Livak, K.J.; Schmitgen, T.D. Analysis of relative gene expression data using realtime quantification PCR and the 2(-Delta Delta CT) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, R.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Yang, N.N.; Yang, X.; Pan, H.P.; Zhou, X.M.; Bai, L.Y.; Xu, B.Y.; et al. Reference gene selection for qRT-PCR analysis in the sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 2013, 8, e53006. [Google Scholar] [CrossRef] [PubMed]
- Teng, X.; Wan, F.; Chu, D. Bemisia tabaci Biotype Q Dominates Other Biotypes Across China. Fla. Entomol. 2010, 93, 363–368. [Google Scholar] [CrossRef]
- Li, X.C.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.W.; Wang, G.H.; Li, Y.H.; Yu, Q.Y.; Zhang, X.S.; Zhu, Y.; Xiang, Z.H. Molecular cloning, sequence analysis and transcriptional activity determination of cytochrome P450 gene CYP18A1 in the silkworm, Bombyx mori. Acta Entomol. Sin. 2008, 51, 237–245. [Google Scholar]
- Lu, K.; Song, Y.Y.; Zeng, R.S. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef]
- Yang, X.; Deng, S.; Wei, X.G.; Yang, J.; Zhao, Q.N.; Yin, C.; Du, T.H.; Guo, Z.J.; Xia, J.X.; Yang, Z.Z.; et al. MAPK-directed activation of the whitefly transcription factor CREB leads to P450-mediated imidacloprid resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 10246–10253. [Google Scholar] [CrossRef]
- Manikandan, P.; Nagini, S. Cytochrome P450 structure, function and clinical significance: A review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef]
- Song, Z.; Gao, X.; Wu, M.; Lu, F.P.; Qin, H.M. Structure, function and application of cytochrome P450 enzymes. Microbiol. China 2020, 47, 2245–2254. [Google Scholar]
- Rauch, N.; Nauen, R. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci (Hemiptera: Aleyrodidae). Arch. Insect Biochem. Physiol. 2003, 54, 165–176. [Google Scholar] [CrossRef]
- Qin, P.H.; Zheng, H.Y.; Tao, Y.L.; Zhang, Y.J.; Chu, D. Genome-Wide Identification and Expression Analysis of the Cytochrome P450 Gene Family in Bemisia tabaci MED and Their Roles in the Insecticide Resistance. Int. J. Mol. Sci. 2023, 24, 5899. [Google Scholar] [CrossRef]
- Wei, X.G.; Hu, J.Y.; Yang, J.; Yin, C.; Du, T.H.; Huang, M.J.; Fu, B.L.; Gong, P.P.; Liang, J.J.; Liu, S.N.; et al. Cytochrome P450 CYP6DB3 was involved in thiamethoxam and imidacloprid resistance in Bemisia tabaci Q (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2023, 194, 105468. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Fang, Y.; Che, W.N.; Zhang, Q.H.; Wang, J.D.; Luo, C. Metabolic Resistance in Abamectin-Resistant Bemisia tabaci Mediterranean from Northern China. Toxins 2022, 14, 424. [Google Scholar] [CrossRef]
- Bonato, O.; Lurette, A.; Vidal, C.; Fargues, J. Modelling temperature-dependent bionomics of Bemisia tabaci (Q-biotype). Physiol. Entomol. 2007, 32, 50–55. [Google Scholar] [CrossRef]
- Zhang, C.; Yan, S.Q.; Shen, B.B.; Ali, S.; Wang, X.M.; Jin, F.L.; Cuthbertson, A.G.S.; Qiu, B.L. RNAi knock-down of the Bemisia tabaci Toll gene (BtToll) increases mortality after challenge with destruxin A. Mol. Immunology. 2017, 88, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Bai, J. High temperature enhance tolerance to abamectin and thiamethoxam in Bemisia tabaci (Hemiptera: Aleyrodidae). 2024, Unpublished manuscript. Last modified 1 May 2024. Microsoft Word file.
- Wang, Y.C.; Chang, Y.W.; Bai, J.; Zhang, X.X.; Iqbal, J.; Lu, M.X.; Du, Y.Z. Temperature affects the tolerance of Liriomyza trifolii to insecticide abamectin. Ecotoxicol. Environ. Saf. 2021, 218, 112307. [Google Scholar] [CrossRef]
- Liu, F.; Miyata, T.; Wu, J.Z.; Li, C.W.; Wu, G.; Zhao, S.X.; Xie, L.H. Effects of temperature on fitness costs, insecticide susceptibility and heat shock protein in insecticide-resistant and -susceptible Plutella xylostella. Pestic. Biochem. Physiol. 2008, 91, 45–52. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insecticides | LC50/mg·L−1 | 95% Confidence Interval | Linear Fitting Equation | R2 | Chi-Square | p Value |
|---|---|---|---|---|---|---|
| AB | 0.1507 | 0.0572−0.2443 | y = 2.4169x + 6.724 | 0.962 | 0.692 | 0.241 |
| TH | 2.1343 | 0.8256−3.4430 | y = 2.5283x + 4.479 | 0.9355 | 1.732 | 0.213 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Liu, Y.; Wang, Y.; Chang, Y.; Wu, Q.; Gong, W.; Du, Y. Effect of High Temperature on Abamectin and Thiamethoxam Tolerance in Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae). Insects 2024, 15, 399. https://doi.org/10.3390/insects15060399
Zhou M, Liu Y, Wang Y, Chang Y, Wu Q, Gong W, Du Y. Effect of High Temperature on Abamectin and Thiamethoxam Tolerance in Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae). Insects. 2024; 15(6):399. https://doi.org/10.3390/insects15060399
Chicago/Turabian StyleZhou, Mi, Yuncai Liu, Yucheng Wang, Yawen Chang, Qingjun Wu, Weirong Gong, and Yuzhou Du. 2024. "Effect of High Temperature on Abamectin and Thiamethoxam Tolerance in Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae)" Insects 15, no. 6: 399. https://doi.org/10.3390/insects15060399
APA StyleZhou, M., Liu, Y., Wang, Y., Chang, Y., Wu, Q., Gong, W., & Du, Y. (2024). Effect of High Temperature on Abamectin and Thiamethoxam Tolerance in Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae). Insects, 15(6), 399. https://doi.org/10.3390/insects15060399