Simple Summary
This study discusses the effects of cyantraniliprole, a new insecticide, on two lepidopteran pests and a biological control agent. Cyantraniliprole showed higher toxicity to one pest species compared to the other, prolonged developmental stages, and increased detoxification activity in both pest species. It also had a negative but slight impact on the biological control agent at the recommended concentrations. The study suggests that cyantraniliprole can effectively control the pests while being compatible with the control agent, but further field studies are needed for validation.
Abstract
Cyantraniliprole is a novel anthranilic diamide insecticide registered for controlling chewing and sucking insect pests. Here, the lethal and sublethal effects of this insecticide on two destructive lepidopteran pests, Spodoptera littoralis Boisduval and Agrotis ipsilon Hufnagel, were evaluated. Because the effects of novel insecticides on beneficial and non-target arthropods must be considered, the impact of cyantraniliprole on a generalist biological control agent, Chrysoperla carnea [Stephens 1836], were also examined. Overall, our study revealed that cyantraniliprole was more toxic to A. ipsilon than to S. littoralis. Moreover, the LC15 and LC50 of the insecticide significantly prolonged the duration of the larval and pupal stages and induced enzymatic detoxification activity in both species. Treatment of the second-instar larvae of C. carnea with the recommended concentration of cyantraniliprole (0.75 mg/L) doubled the mortality rates and resulted in a slight negative effect on the biology and detoxification enzymes of C. carnea. Our results indicate that both sublethal and lethal concentrations of cyantraniliprole can successfully suppress S. littoralis and A. ipsilon populations. They also suggest that C. carnea, as a generalist predator, is compatible with cyantraniliprole under the modelled realistic field conditions. In future investigations, insights into the effects of cyantraniliprole on S. littoralis, A. ipsilon, and C. carnea under field conditions will be required to appropriately validate our results.
1. Introduction
Economic losses in agriculture due to insect pests often necessitate the use of chemical pesticides [1], which have historically been the primary method for controlling pest populations. In recent decades, alternative control methods, such as biological control, have become more appealing [2]. Given that one primary focus of integrated pest management (IPM) is the use of non-chemical control measures, the applicability of biological control methods measures is under constant consideration [3]. However, natural enemies are not always successful in pest management, and more immediate corrective interventions to suppress the growth of pest populations are often needed [3,4]. One limitation of relying on chemical control methods is that broad-spectrum insecticides can interfere with natural biological control [5]. Therefore, it is crucial to assess the impact of insecticides on biological control agents when developing effective pest control strategies [6,7].
Noctuidae is the most challenging family in the Noctuoidea superfamily [8]. It consists of about 1150 genera comprised of more than 12,000 diverse and ecologically successful species [9,10]. Noctuid moths are the most important agricultural pests globally and include species capable of adapting to new climatic or ecological situations, such as cotton leafworm, Spodoptera littoralis [Boisduval, 1833], and black cutworm, Agrotis ipsilon [Hufnagel, 1766] [11]. Unfortunately, both insects have developed resistance to conventional insecticides, including organophosphates, carbamates, and pyrethroids [12,13,14], as well as to several newer insecticides [15,16,17]. S. littoralis is a highly polyphagous and destructive pest that attacks a wide range of economically important crops throughout the world and causes yield losses of up to 50%, typically due to larval foraging activity [18,19]. A. ipsilon is also a major polyphagous pest that damages more than 30 important crops worldwide [20]. The larvae of A. ipsilon can consume more than 400 cm2 of foliage during their development [21,22].
As a potential predator, the green lacewing, Chrysoperla carnea [Stephens, 1836] (Neuroptera: Chrysopidae), is a polyphagous foliage-dwelling predator that has been promoted as a biological agent for a wide range of pest species, including aphids, lepidopteran eggs and neonates, whiteflies, leafhoppers, scales, mites, and other soft-bodied insects [23,24,25]. Several studies have been performed on the predation capacity of C. carnea on lepidopteran prey [26,27,28], including eggs and neonate larvae of S. littoralis [29] and cotton bollworm, Helicoverpa armigera [Hübner, 1808] [30]. Although green lacewings are found in a wide range of agroecosystems [25], they are mainly utilized to control greenhouse and outdoor crop pests through augmentative releases [31,32]. The excessive and indiscriminate use of pesticides [33] has negatively impacted field populations of C. carnea [34,35] and necessitated the use of more selective insecticides.
Diamide insecticides have emerged as the most recent and successful chemistries for controlling both sap-feeding insect pests and chewing pests [36]. Several studies have shown that this group has a high insecticidal activity on different lepidopteran species such as tobacco cutworm, Spodoptera litura [Fabricius, 1775] [37], Spodoptera exigua (Hübner, 1808) [38], S. littoralis [39], A. ipsilon [13,22], and Mamestra brassicae [Linnaeus, 1758] [40]. Unfortunately, diamide insecticides can also harm natural arthropod enemies in agricultural systems, thus triggering secondary pest outbreaks [41].
Cyantraniliprole is a second-generation diamide insecticide that is widely used to control various lepidopteran, coleopteran, dipteran, and hemipteran pests [42,43]. As it has systemic and translaminar behavior, it can be employed in soil or as a foliar application for pest management [44]. As a highly potent agonist of insect ryanodine receptors, cyantraniliprole activates the ryanodine receptor channels [45] that disrupt calcium balance.
Generally, insecticides have varying effects on insect defense enzyme systems, which are usually associated with insect death or the development of resistance. Several studies have shown that insects protect themselves against insecticides by increasing the activity of detoxifying enzymes such as cytochrome P450 monooxygenases (P-450s), esterases, and glutathione S-transferases (GSTs) [39]. Additionally, the insecticide breakdown often exposes insects to sublethal concentrations following application in crops, which impacts various biological and biochemical parameters [46]. So, determining the sublethal effects of insecticides on insects is crucial for understanding their impact on agroecosystems and prolonging the long-term efficiency of these insecticides in insect management. Therefore, this study aims to assess the effect of cyantraniliprole on two lepidopteran pests, S. littoralis and A. ipsilon, and on C. carnea as a potential predator of the two species. This study will hopefully allow for a better understanding of pest and predator interactions and help develop a more effective IPM program.
2. Materials and Methods
2.1. Insect Cultures
2.1.1. Target Pests
Cultures of S. littoralis and A. ipsilon were obtained from the Department of Entomology at the Faculty of Agriculture, Cairo University, Egypt. S. littoralis was maintained in the laboratory at 25 ± 2 °C and 55 ± 5% RH [47,48,49], while A. ipsilon was maintained at 27 ± 2 °C and 55 ± 5% RH [48,49]. Sexed pupae of both insects were kept covered in glass jars lined with paper towels until adult emergence. The incipient adult moths (seven males and five females) were transferred to a larger jar draped with cotton wool soaked in a 10% sugar solution as dietary supplement [46]. The deposited eggs were collected daily and transferred to new jars for hatching [14]. Neonates were fed castor oil leaves, and a colony of mass-reared larvae was maintained as described above. Experimental treatments were conducted using the second-instar larvae of S. littoralis and A. ipsilon.
2.1.2. Non-Target Pest
C. carnea larvae were obtained from the Biological Control Laboratory in Aswan Governorate, Egypt. Larval cultures were reared on UV-sterilized eggs of Ephestia kuehniella [Zeller, 1879] that were stored at 2–4 °C. Adults were maintained on an artificial diet consisting of honey, yeast, and water (7:4:4, respectively) in sterile plastic containers (17 × 25 cm) covered with muslin cloth in insect rearing room at 25 °C and 60% RH and 14:10 h- dark–light intervals. The colony was maintained for two generations without an infusion of wild stock before experiments on the second-instar larvae began.
2.2. Insecticide
The commercial formulation of cyantraniliprole Benevia 100 OD (FMC Company, Philadelphia, PA, USA) was used in this study with a field rate of application of 75 mL/100 L water.
2.3. Toxicity of Cyantraniliprole to Target Species
Cyantraniliprole toxicity bioassays on newly ecdysed second-instar larvae of S. littoralis and A. ipsilon were performed as described by [39] and [22]. Insecticide dosages were optimized in preliminary studies. Five concentrations (5, 2.5, 1.25, 0.125, and 0.0125 mg/L) of cyantraniliprole were used to determine LC15 and LC50, and experiments were repeated twice to confirm the results. The leaf-dipping technique was used according to [50] as follows: for experiments, castor leaves were dipped in each concentration for 20 s and then allowed to dry in the air for 30 min. Five replicates with ten larvae each were used for each concentration. Leaves dipped in water were used in the control group. The larvae were allowed to feed on treated leaves for 24 h, and the surviving larvae were then transferred to a clean dry container containing fresh untreated leaves [39,50]. Larval mortality was recorded daily for four consecutive days post-treatment, and the lethal and sublethal concentrations were calculated [14] and corrected using Abbott’s formula [51].
2.4. Sublethal and Lethal Effects of Cyantraniliprole on Target Species
The impact of the cyantraniliprole LC15 and LC50 on S. littoralis and A. ipsilon development was assessed. Three replicates with 50 larvae each were used for each concentration/control (n = 150). Ninety-six hours after treatment, the surviving larvae were kept individually in a small, dry, clean cup with fresh, untreated castor leaves [14]. Developmental changes, i.e., larval and pupal durations (days), pupation percentage, pupal weight [g], adult emergence percentage, and sex ratio of S. littoralis and A. ipsilon, were recorded daily [50]. Additionally, fecundity and hatchability percentages were assessed using three replicates, each of five females and seven males [52].
2.5. Sublethal Effects of Cyantraniliprole on C. carnea
Toxicity bioassays were conducted on the second-instar larvae of C. carnea in a controlled chamber using two methods under standardized environmental conditions. Three concentrations of cyantraniliprole were used: the recommended concentration (0.75 mg/L), half the recommended concentration (0.37 mg/L), and a quarter of the recommended concentration (0.19 mg/L). To assess the acute toxicity of cyantraniliprole to C. carnea larvae, the larval mortality was recorded 24 h post-treatment and then for 120 h. Two insecticide contamination methods were used:
- The direct contact method: IOBC standard guidelines for C. carnea [53] were followed with some modifications. To estimate the residual effects of cyantraniliprole on C. carnea larvae, filter paper was dipped for 5 s in each concentration. For the control group, the filter paper was dipped in water. The treated and untreated paper was then allowed to dry in the air. For each treatment, five replicates with 10 larvae each were used. The larvae were placed on the contaminated filter paper for 24 h, after which they were transferred to individual Petri dishes (5 cm) to avoid cannibalism and provided with E. kuehniella eggs every two days.
- The insecticide-treated host method (feeding method): E. kuehniella egg cards [1 × 1 cm] were dipped for 5 s in cyantraniliprole concentrations, and each card was then placed in a 5 cm Petri dish with a single second-instar larva of C. carnea for 24 h before being replaced by an untreated one.
2.6. Sublethal Effects on C. carnea
The effect of field exposure to cyantraniliprole on the larval and pupal duration (days), pupation percentage, and emergence percentage was assessed. For each treatment, five replicates of second-instar larvae, each comprising 10 individuals, were utilized. The following formulas were used to calculate pupation and emergence percentages: pupation (%) = 100 × (total number of pupae/total number of larvae), adult emergence (%) = 100 × (total number of adults/total number of pupae).
2.7. Biochemical Assays
2.7.1. Sample Preparation
Second-instar larvae of the three insects were exposed to LC15 and LC50 of cyantraniliprole, as described above. A total of 50 mg fresh body weight of the surviving larvae were then used to measure the activities of detoxifying enzyme at 24, 48, 72, and 96 h post-treatment in S. littoralis and A. ipsilon and 120 h post-treatment in C. carnea. Five separate replicates were used for each analysis. The larvae were homogenized in 0.1 M phosphate buffer, pH 7.0 for carboxylesterase (CarE), or pH 6.5 for glutathione S-transferase (GST). The homogenates were centrifuged at 7000 rpm for 15 min, and the supernatants were used to determine enzyme activity and protein content, as described below.
2.7.2. CarE Assay
Alpha (α)- and beta (β)-esterase activities were measured as described by [54] and [40]. The homogenate was incubated for 15 min at 25 °C with alpha or beta-naphthyl acetate as the substrate. A mixture of Fast Blue B and sodium dodecyl sulfate was added to stop the reaction. The optical density was measured at 550 nm for α- esterase and 600 nm for β- esterase using a Jenway Spectrophotometer-7205UV/Vis, Dunmow, Essex, UK.
2.7.3. GST Assay
GST activity was determined as described by [55] and [40]. The assay mixture contained the sample enzyme solution with CDNB (1-chloro-2, 4-dinitrobenzene) as the substrate and GSH as a reagent. The optical density was recorded at 340 nm for 3 min with readings taken at 1-min intervals, using a Jenway Spectrophotometer-7205 UV/Vis., UK.
2.7.4. Protein Content
Coomassie brilliant blue assay [56] was used to determine the protein concentration.
2.8. Data Analysis
Four days post-exposure to cyantraniliprole, LC15 and LC50 to the second-instar larvae of S. littoralis and A. ipsilon were estimated using Probit analysis (EPA Probit analysis program, V. 1.5) [57]. All the biological and biochemical data were coded and entered using the statistical package SPSS V.22. Data were tested for satisfying assumptions of parametric tests, and continuous variables were subjected to the Shapiro–Wilk and Kolmogorov–Smirnov test for normality. Probability and percentile data were standardized for normality using Arcsine Square Root. Data were presented as mean and standard deviation. ANOVA was done for experimental groups using three replicates at least for each group, while post-hoc analysis was done using Tukey’s pairwise comparison. p-value was considered significant at <0.05. Chi (ꭓ2) was used for comparing the observed and expected frequencies of sex ratio using MiniTab (V. 14). Finally, data were visualized using R studio (V. 2022.02.4).
3. Results
3.1. Toxicity of Cyantraniliprole to S. littoralis and A. ipsilon
As shown in Table 1, the results of cyantraniliprole toxicity to the second-instar larvae of both insects revealed that S. littoralis larvae were more tolerant than A. ipsilon. The cyantraniliprole LC15 values were 1.95 and 0.019 mg/L, whereas the LC50 values were 8.17 and 0.33 mg/L to S. littoralis and A. ipsilon, respectively.

Table 1.
Toxicity of cyantraniliprole to the second-instar larvae of S. littoralis and A. ipsilon.
3.2. Toxicity of Cyantraniliprole to C. carnea
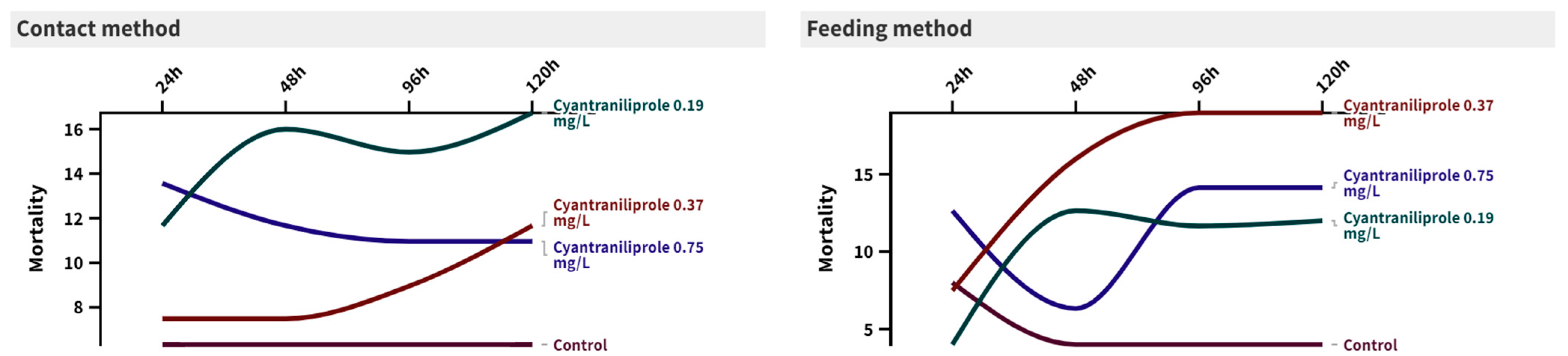
The toxicity of three concentrations of cyantraniliprole to the second-instar larvae of C. carnea is presented in Figure 1. It can be observed that the interactions between time and treatment were not significant for either the direct contact method or the feeding method. The recommended concentration (0.75 mg/L) of cyantraniliprole showed the highest mortality % after 24 h in both contact and feeding methods (24 ± 13.56 (F = 2.29, p = 0.117) and 10 ± 12.64 (F = 0.63, p = 0.606), respectively), compared to the control (10 ± 6.32 and 6 ± 8, respectively). After 120 h, the mortality % insignificantly increased, compared to the control, for both methods (30 ± 10.95 (F = 1.88, p = 0.174) and 30 ± 14.14 (F = 1.45, p = 0.265), respectively).
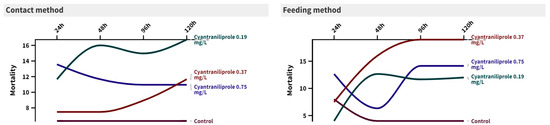
Figure 1.
Mortality percentages of C. carnea at 24, 48, 96, and 120 h post-treatment with three concentrations (0.19, 0.37, and 0.75 mg/L) of cyantraniliprole using two different methods of exposure.
3.3. Lethal and Sublethal Effects of Cyantraniliprole
3.3.1. On S. littoralis and A. ipsilon
The effects of cyantraniliprole on the development of S. littoralis and A. ipsilon are presented in Table 2 and Table 3. Treating the freshly molted second-instar larvae of both insects with LC15 and LC50 of cyantraniliprole resulted in the significant prolongation of larval (F = 106.98, p = 0.000 and F = 11.15, p = 0.000, respectively) and pupal (F = 48.40, p = 0.000 and F = 91.91, p = 0.000, respectively) durations (Table 2). Nevertheless, no significant differences were observed (Table 2) in the pupation rate (F = 4.45, p = 0.065 and F = 0.65, p = 0.556, respectively), emergence percentages (F = 3.0, p = 0.125 and F = 0.25, p = 0.788, respectively), or sex ratio (Table 3) of the emerged adults for both S. littoralis and A. ipsilon, respectively. Similarly, no significant differences (F = 1.26, p = 0.348 and F = 0.68, p = 0.543, respectively) were observed in fecundity (eggs laid per female) for both S. littoralis and A. ipsilon, respectively (Table 2).
Table 2.
Effects of cyantraniliprole on the development, fecundity, and fertility of S. littoralis and A. ipsilon.
Table 3.
Effect of exposure to LC15 and LC50 of cyantraniliprole via the leaf dipping method on sex ratio of S. littoralis and A. ipsilon.
As for the hatchability rate, it significantly decreased in both species after treatment with the LC50 of cyantraniliprole. For S. littoralis, cyantraniliprole treatments also caused a significant reduction in pupal weights in both males (F = 19.59, p = 0.000) and females (F = 29.45, p = 0.000) compared to the control. For A. ipsilon, a significant difference in pupal weight was only observed in males (F = 3.58, p = 0.032), while in females (F = 1.33, p = 0.270), there was no significant difference compared to the control treatment (Table 2).
3.3.2. On C. carnea
The effects of cyantraniliprole on C. carnea development are presented in Table 4. Regarding the effect on larval duration, pupation rate, and pupal duration, no two-way interaction was detected between the tested concentrations and the contamination methods (two-way ANOVA, p > 0.05). Additionally, cyantraniliprole did not affect larval duration, pupation rate, or pupal duration at any concentration. Although cyantraniliprole decreased the emergence ratio at all concentrations, compared to the control, this reduction was statistically significant only at 0.37 and 0.75 mg/L (F = 25.12, p = 0.011) in the contact method. However, at a concentration of 0.75 mg/L cyantraniliprole significantly reduced the emergence ratio by 1.6-fold (F = 4.26, p = 0.022) in the feeding method, compared to the control.
Table 4.
Effect of cyantraniliprole on C. carnea development.
3.4. Effect of Cyantraniliprole on Detoxifying Enzymes
3.4.1. S. littoralis and A. ipsilon
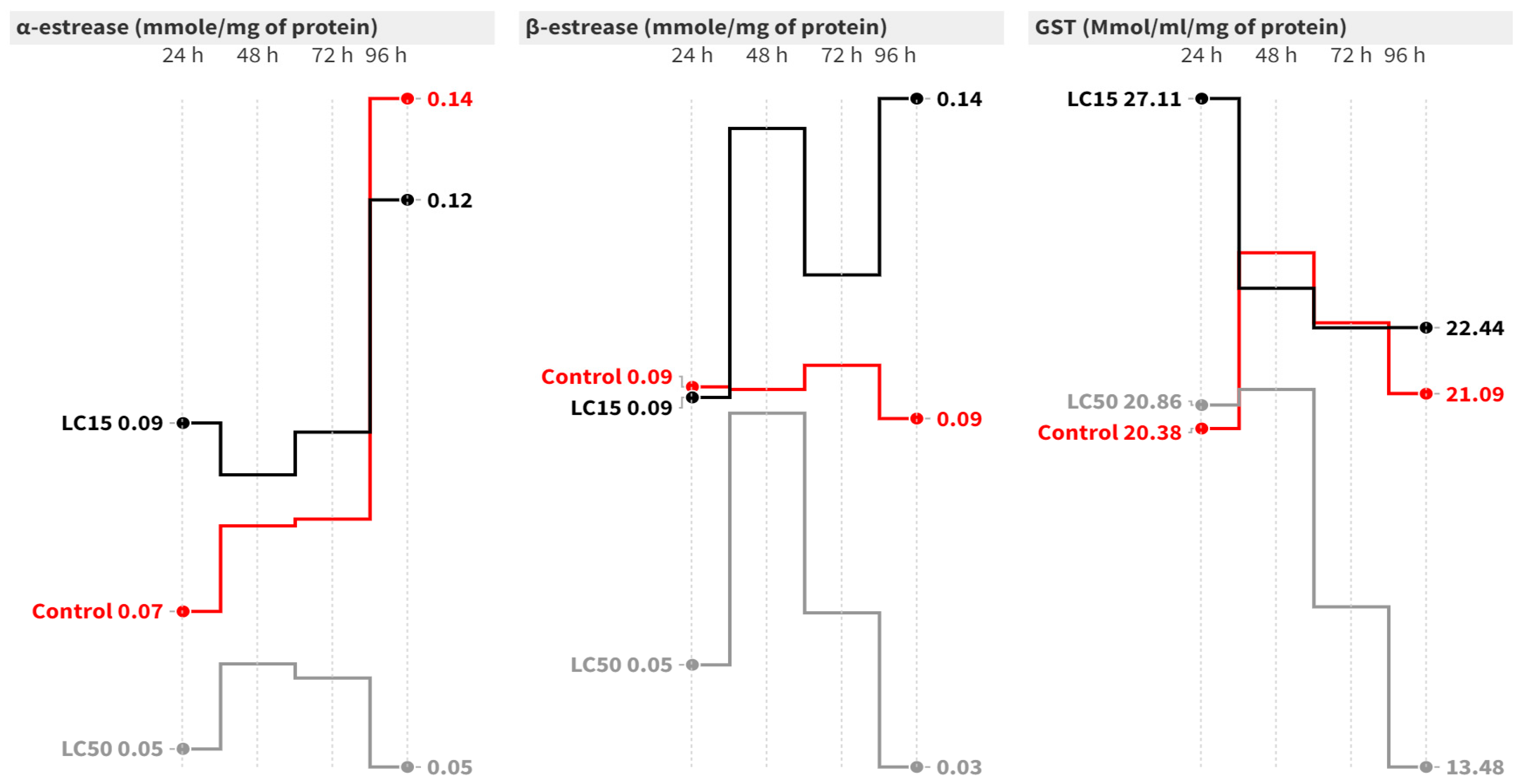
In the case of S. littoralis, the elapsed time had no significant effect on the α-esterase activity at the LC50 level (Figure 2). At 24 and 96 h post-contamination with LC50, α-esterase activity significantly decreased by 2.25- [F = 11.32, p = 0.009] and 3.07-fold [F = 8.35, p = 0.018], respectively, compared to the control. Concerning the β-esterase assay, both the elapsed time and treatment had significant effects. The LC15 caused a significant increase in β-esterase activity at 48 and 96 h post-contamination compared to the control group [Figure 2]. On the other hand, the LC50 significantly increased β-esterase activity at all intervals except after 96 h, where the activity dropped to 0.03 mmol/mg of protein [F = 33.76, p = 0.001] compared to the control [Figure 2].
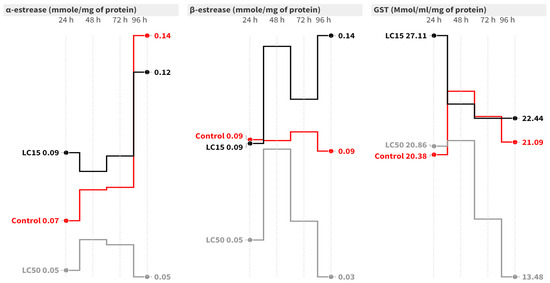
Figure 2.
Mean of the detoxification enzyme activity of carboxylesterase (α- and β-esterase) and GST at 24, 48, 72, and 96 h post-treatment of S. littoralis second-instar larvae with LC15 and LC50 of cyantraniliprole.
Concerning GST activity in S. littoralis larvae, the elapsed time did not significantly change the GST activity in the control or the LC15 groups. However, treatment with LC50 caused a significant reduction in GST activity at 96 h post-contamination (F = 23.64, p = 0.001) compared to the control (Figure 2).
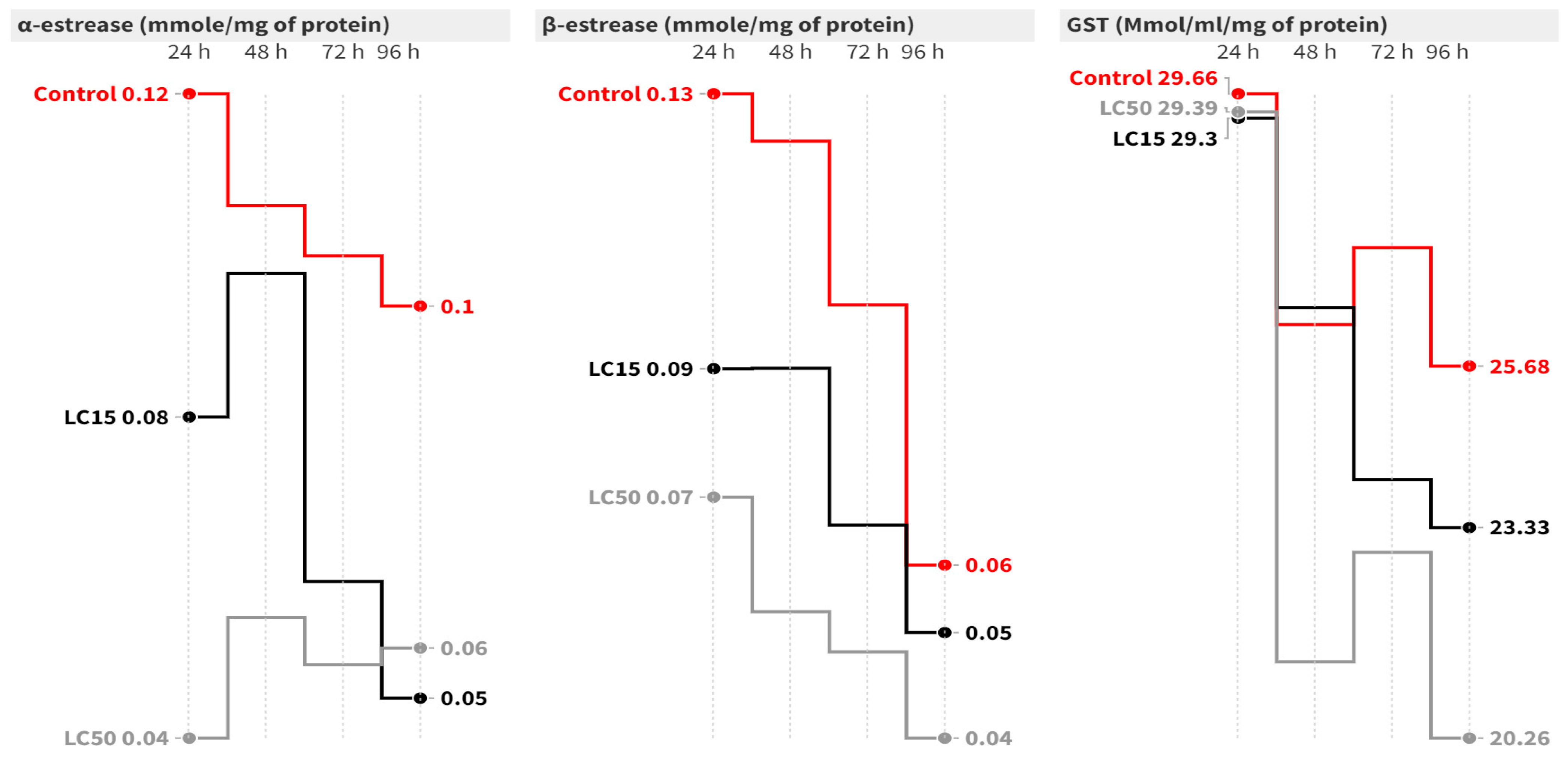
In the case of A. ipsilon, at 96 h post-contamination, the LC15 of cyantraniliprole caused a significant inhibition of 2.25-fold [F = 9.20, p = 0.015] in the α-esterase activity compared to the control group [Figure 3]. On the other hand, the LC50 of cyantraniliprole significantly decreased α-esterase activity at 24 [F = 7.80, p = 0.021], 48 [F = 17.49, p = 0.003], 72 (F = 6.44, p = 0.032) and 96 h (F = 9.20, p = 0.015) post-contamination. Concerning β-esterase, the highest activity was recorded in the control group, while significantly lower activity was observed in the LC15 and LC50 groups. The LC15 caused a significant inhibition in β-esterase activity at 48 h (F = 25.34, p = 0.001) post-contamination, while the LC50 significantly decreased the activity at 24 (F = 7.63, p = 0.022), 48 (F = 25.34, p = 0.001), 72 (F = 11.40, p = 0.009), and 96 h (F = 1.73, p = 0.254) post-contamination = compared to the untreated group (Figure 3). Concerning GST activity, it insignificantly declined at all intervals in all groups compared to the control group (Figure 3).
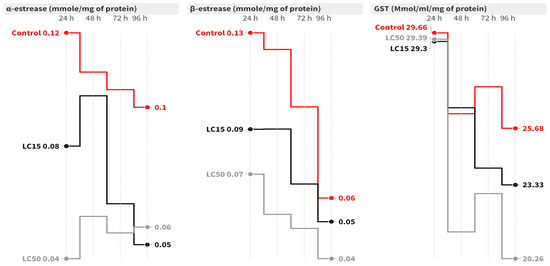
Figure 3.
Mean of the detoxification enzyme activity of carboxylesterase (α- and β-esterase) and GST at 24, 48, 72, and 96 h post-treatment of A. ipsilon second-instar larvae with LC15 and LC50 of cyantraniliprole.
3.4.2. C. carnea
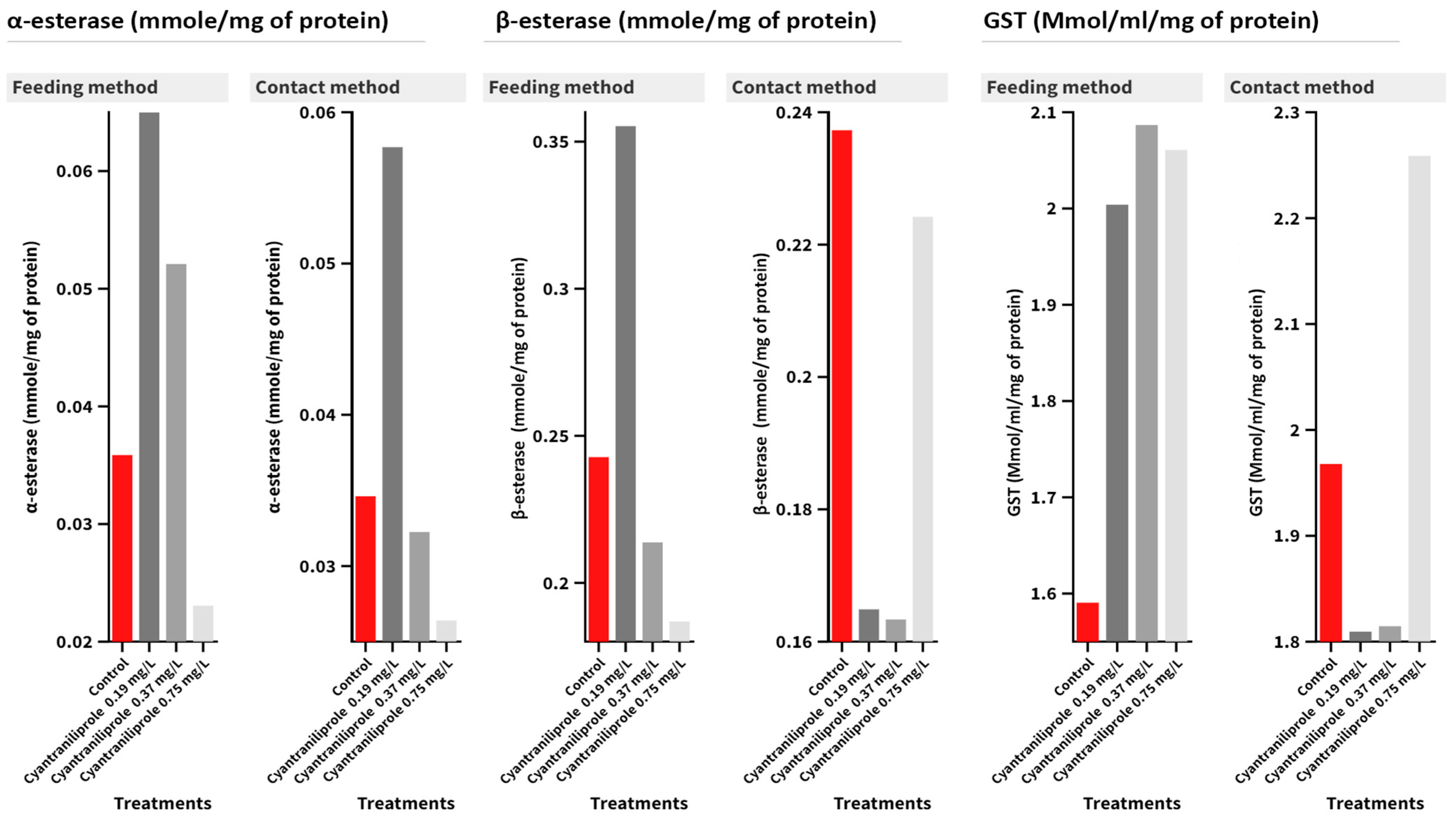
In the case of α-esterase activity, no significant interaction was detected between the treatment and administration method (Figure 4). Cyantraniliprole treatments insignificantly affected the activity, while no effect was recorded in the feeding method (F = 3.56, p = 0.067) or the contact one (F = 6.13, p = 0.018). As for β-esterase activity, the interaction between treatments and contamination methods was insignificant. Except in 0.19 mg/L-treated larvae in the feeding contamination method (F = 12.94, p = 0.002), there were significant differences when compared with the control. As for GST activity, in the feeding method, there was a significant difference (F = 3.92, p = 0.054), while in the contact method, there were no significant differences in GST activity (F = 0.89, p = 0.487) compared to the control treatment (Figure 4).
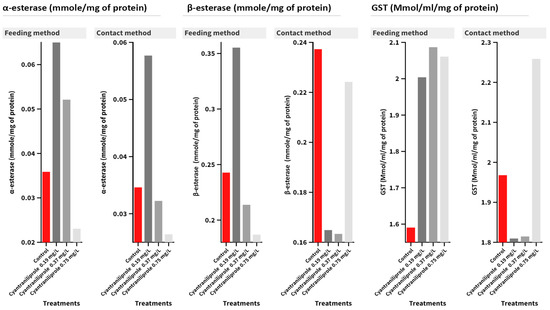
Figure 4.
Mean of the detoxification enzyme activity of carboxylesterase (α- and β-esterase) and GST at 96 h post-treatment of C. carnea second-instar larvae with the recommended (0.75 mg/L), half recommended (0.37 mg/L), and quarter recommended (0.19 mg/L) dosages of cyantraniliprole.
4. Discussion
The management of arthropod pests in vegetables and high-value crops relies heavily on chemical control. However, there are many insecticide application options to choose from, and each differs in its effectiveness, costs, and risks to the environment and human health [58,59,60,61]. Therefore, it is necessary to make well-founded decisions that consider how pesticides impact all aspects of pest management including natural enemies, crop production, economic outcomes, and resistance [16,62]. In this study, we assessed the effects of cyantraniliprole as a novel insecticide on the potential management of two destructive lepidopteran pests, S. littoralis and A. ipsilon. Additionally, we assessed the adverse effect of this insecticide on the biological control agent C. carnea.
Cyantraniliprole exhibited insecticidal activity against the second-instar larvae of both S. littoralis and A. ipsilon. The LC50 to S. littoralis was significantly higher (~24.7-fold) than that to A. ipsilon. This indicates that A. ipsilon is more susceptible to cyantraniliprole. This is consistent with [13], who found, using an artificial diet, that cyantraniliprole caused high mortality in the fourth-instar A. ipsilon larvae (LC50 of 0.354 μg g−1). In addition to mortality, sublethal effects on larval development due to perturbations in the development of neural tissues by neurotoxic substances [35] may play a key role in managing insect pests [63]. In the current study, the larval and pupal durations of S. littoralis and A. ipsilon were prolonged due to exposure to cyantraniliprole. This could indicate that cyantraniliprole-treated larvae invest more resources in detoxification rather than development, and as a result, development is slowed relative to the control group [64]. Accordingly, pupal weight significantly decreased in all treatments, compared to the control groups. Similarly, the hatching rate decreased in both species as the cyantraniliprole LC50 significantly reduced emergence ~1.2 and 1.17 times in S. littoralis and A. ipsilon, respectively. Additionally, LC50 of cyantraniliprole insignificantly reduced the number of eggs laid per family ~1.4 and 1.2 times in both species, respectively, as in several studies that reported that the average number of eggs laid by adult females decreased after treatment with anthranilic diamide [13,38,65,66].
Insects in the natural environment are exposed to lethal and sublethal concentrations of insecticides [67], which may affect diverse enzymatic activities [68]. Generally, detoxification enzyme activity, such as CarEs and GSTs, can be useful for monitoring insecticide resistance development [69,70]. In addition, these enzymes are one of the adaptation mechanisms that enhance the insect’s metabolic capacity to counteract pesticides [71]. Thus, the competence of insects to detoxify is reflected by the reaction of detoxification enzymes to insecticides [17]. The exposure to lethal and sublethal concentrations (LC50 and LC15) of cyantraniliprole induced the activity of detoxification enzymes in both S. littoralis and A. ipsilon compared to the control group. In addition, medium to high levels of diamide resistance could be mediated by target site mutations and enhanced detoxification triggered by up-regulating the expression of genes encoding detoxification enzymes [72]. As revealed by our results, α-esterase activity significantly increased 1.3-fold after the S. littoralis second-instar larvae were treated with the LC15. Overall, this suggests that α-esterase is a key enzyme in S. littoralis detoxification. Likewise, [68] showed that a sublethal concentration of cyantraniliprole significantly increased CarE activity in the small brown planthopper Laodelphax striatellus [Fallén, 1826]. The activities of S. littoralis β-esterase and GST, however, decreased after treatment with the cyantraniliprole LC50. This decrease in detoxification enzyme activity is typically considered a positive marker of delayed resistance to toxic compounds [73]. Comparable results were observed in the activities of A. ipsilon CarE and GST following exposure to sublethal dosages of cyantraniliprole.
As new insecticides are registered, their effects on the arthropod community must be evaluated to ensure the important ecosystem services provided by predators in the farming system. Thus, the use of selective insecticides is crucial for maintaining the integral role of biological control in the food web [74,75]. Indiscriminate pesticide usage in agricultural fields can disrupt beneficial insect activities by reducing their overall abundance and species composition [76]. For example, some insecticides can block physiological or biochemical processes that affect the survival, growth, development, reproduction, and/or behavior of some natural enemy species [76,77]. Our study evaluated the effects of cyantraniliprole on C. carnea. Although cyantraniliprole had no significant effects on C. carnea larval mortality (two-fold increase with 0.75 mg/L at 24 h), the recommended dose of the insecticide prolonged larval duration and significantly reduced C. carnea emergence (Table 4). However, a potential effect of several conventional chemical insecticides has been observed on non-target organisms including Chrysoperla zastrowi (Sillemi) [78], Chrysoperla nipponensis (Steinmann, 1964) [79], and C. carnea [80]. Evaluating lifetable parameters can also provide insights into the response of natural enemies to selective insecticides from a population viewpoint [81]. Although cyantraniliprole at concentrations of 0.37 and 0.72 mg/L decreased the C. carnea emergence ratio relative to the control group, no significant differences were observed in larval duration, pupal duration, or pupation rate compared to the control. In addition, there was no significant interaction between the treatment and contamination methods when detoxification enzyme activities were assayed. However, the 0.19 mg/L cyantraniliprole concentration significantly increased α-esterase activity compared to the control and the two other concentrations evaluated. On the other hand, significant effects on C. carnea GST activity were observed only after 96 h, regardless of treatment or contamination method at LC50 value. Generally, arthropods rely heavily on detoxification enzymes for endogenous insecticide production. These enzymes can remove toxins and detoxify insecticides [82]. Therefore, several studies elucidated the role of these enzymes on lepidopteran insect pests [17,22,39,62], which suggests the possibility of discovering new insecticidals that act by interfering with metabolic-mediated enzymes in insects [83].
5. Conclusions
Overall, our study shows that cyantraniliprole is more toxic to A. ipsilon than to S. littoralis. Moreover, the LC15 and LC50 can significantly prolong both larval and pupal duration, which could help in the management of both pest populations. On the other hand, the use of selective insecticides in conjunction with biocontrol agents will increase biological efficacy and reduce the environmental consequences of incorporating insecticides in IPMs. Our data suggest that the predator C. carnea is compatible with cyantraniliprole under the modeled realistic field conditions. In future investigations, insights into the effects of cyantraniliprole on A. ipsilon, S. littoralis, and C. carnea under field conditions will be required to appropriately validate our results.
Author Contributions
M.A. and M.A.M.M. were responsible for conceptualization, methodology, software, validation, formal analysis, visualization, supervision, project administration, writing—original draft preparation, writing—review, and editing. A.H.E.K. and E.-D.S.I. helped with methodology, writing—original draft preparation, and writing—review, visualization, and supervision. N.A.A. contributed to the formal analysis, supervision, and resources. Finally, J.K.J. and A.F. helped with software, validation, formal analysis, writing—original draft preparation, writing—review, and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The datasets generated during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank J. Joe Hull for English editing assistance. Additionally, we thank Hamed Khair-Allah for improving the manuscript considerably.
Conflicts of Interest
The authors declare no competing interests.
References
- Singh, B.; Kaur, A. Control of insect pests in crop plants and stored food grains using plant saponins: A review. LWT Food Sci. Technol. 2018, 87, 93–101. [Google Scholar] [CrossRef]
- Jayasekhar Babu, P.; Saranya, S.; Longchar, B.; Rajasekhar, A. Nanobiotechnology-mediated sustainable agriculture and post-harvest management. Curr. Res. Biotechnol. 2022, 4, 326–336. [Google Scholar] [CrossRef]
- Jalali, M.A.; Van Leeuwen, T.; Luc, T.; De Patrick, C. Toxicity of selected insecticides to the two-spot ladybird Adalia bipunctata. Phytoparasitica 2009, 37, 323–326. [Google Scholar] [CrossRef]
- Medina, P.; Budia, F.; del Estal, P.; Adán, A.; Viñuela, E. Toxicity of fipronil to the predatory lacewing Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Biocontrol Sci. Technol. 2004, 14, 261–268. [Google Scholar] [CrossRef]
- Moscardini, V.F.; da Costa Gontijo, P.; Carvalho, G.A.; Oliveira, R.L.; Maia, J.B.; Silva, F.F. Toxicity and sublethal effects of seven insecticides to eggs of the flower bug Orius insidiosus (Say) (Hemiptera: Anthocoridae). Chemosphere 2013, 92, 490–496. [Google Scholar] [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Eiden, A.; Cooper, R.; Zha, C.; Wang, D.; Reilly, E. Changes in indoor insecticide residue levels after adopting an integrated pest management program to control German Cockroach infestations in an apartment building. Insects 2019, 10, 406. [Google Scholar] [CrossRef] [PubMed]
- Regier, J.C.; Mitter, C.; Mitter, K.; Cummings, M.P.; Bazinet, A.L.; Hallwachs, W.; Janzen, D.H.; Zwick, A. Further progress on the phylogeny of Noctuoidea (Insecta: Lepidoptera) using an expanded gene sample. Syst. Entomol. 2017, 42, 82–93. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, H.; Helen, H.-S.; Wang, J.-J. Effect of host plants on development, fecundity and enzyme activity of Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). Agric. Sci. China 2011, 10, 1232–1240. [Google Scholar] [CrossRef]
- Keegan, K.L.; Rota, J.; Zahiri, R.; Zilli, A.; Wahlberg, N.; Schmidt, B.C.; Lafontaine, J.D.; Goldstein, P.Z.; Wagner, D.L. Toward a stable global Noctuidae (Lepidoptera) taxonomy. Insect Syst. Divers. 2021, 5, 1–24. [Google Scholar] [CrossRef]
- Henaish, M.; Elmetwaly, N. Identification and taxonomic notes of Spodoptera species (Lepidoptera: Noctuidae) known to occur in Egypt. Egypt. Acad. J. Biol. Sci. A Entomol. 2020, 13, 161–175. [Google Scholar] [CrossRef]
- Abo-Elghar, G.C.; Rashwan, M.H.; El-Bermawy, Z.A.; Radwan, H.S.; Hussien, A.H. Monitoring for resistance in the cotton leafworm, Spodoptera littoralis (Boisd.) against mixtures with benzoylphenyl ureas. Bull. Entomol. Soc. Egypt Econ. Ser. 1992, 19, 249–259. [Google Scholar]
- Xu, C.; Zhang, Z.; Cui, K.; Zhao, Y.; Han, J.; Liu, F.; Mu, W. Effects of sublethal concentrations of cyantraniliprole on the development, fecundity and nutritional physiology of the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae). PLoS ONE 2016, 11, e0156555. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.M.; Elmenofy, W.H.; Osman, E.A.; El-Said, N.A.; Awad, M. Biological impact, oxidative stress and adipokinetic hormone activities of Agrotis ipsilon in response to bioinsecticides. Plant Prot. Sci. 2022, 58, 326–337. [Google Scholar] [CrossRef]
- Hafez, S.S.M.; EL-Malla, A.; Ali, R.E.; El-Hadek, M.K. Resistance monitoring in cotton leafworm Spodoptera littoralis to certain bioinsecticides during ten cotton seasons in eight governorates on Egypt. J. Biol. Chem. 2018, 35, 590–594. [Google Scholar]
- Fouad, E.A.; Ahmed, F.S.; Moustafa, M.A.M. Monitoring and biochemical impact of insecticides resistance on field populations of Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae) in Egypt. Pol. J. Entomol. 2022, 91, 109–118. [Google Scholar] [CrossRef]
- Moustafa, M.A.M.; Osman, E.A.; Mokbel, E.M.S.; Fouad, E.A. Biochemical and molecular characterization of chlorantraniliprole resistance in Spodoptera littoralis (Lepidoptera: Noctuidae). Crop Prot. 2024, 177, 106533. [Google Scholar] [CrossRef]
- Garrido-Jurado, I.; Montes-Moreno, D.; Sanz-Barrionuevo, P.; Quesada-Moraga, E. Delving into the Causes and Effects of Entomopathogenic Endophytic Metarhizium brunneum Foliar Application-Related Mortality in Spodoptera littoralis Larvae. Insects 2020, 11, 429. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Life table parameters and digestive enzyme activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on selected legume cultivars. Insects 2022, 13, 661. [Google Scholar] [CrossRef]
- Sayed, R.M.; El Sayed, T.S.; Rizk, S.A. Potency of Bio Magic (Metarhizium anisopliae fungus) and gamma radiation in the black cut worm, Agrotis ipsilon (Hufnagel) larvae. Egypt. J. Biol. Pest Control 2023, 33, 1. [Google Scholar] [CrossRef]
- Amin, A.H.; Bayoumi, A.E.; Dimetry, A.Z.; Youssef, D.A. Efficiency of Nano-formulations of neem and peppermint oils on the bionomics and enzymatic activities of Agrotis ipsilon larvae (Lepidoptera: Noctuidae). Int. J. Nat. Resour. Ecol. Manag. 2019, 4, 102–111. [Google Scholar] [CrossRef]
- Awad, M.; Ibrahim, E.S.; Osman, E.I.; Elmenofy, W.H.; Mahmoud, A.W.M.; Atia, M.A.M.; Moustafa, M.A.M. Nano-insecticides against the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae): Toxicity, development, enzyme activity, and DNA mutagenicity. PLoS ONE 2022, 17, e0254285. [Google Scholar] [CrossRef]
- Tauber, M.J.; Tauber, C.A.; Daane, K.M.; Hagen, K.S. Commercialization of predators: Recent lessons from green lacewings (Neuroptera: Chrysopidae: Chrysoperla). Am. Entomol. 2000, 46, 26–38. [Google Scholar] [CrossRef]
- New, T.R. Introduction to the systematics and distribution of Coniopterygidae, Hemerobiidae, and Chrysopidae used in pest management. In Lacewings in the Crop Environment; Cambridge University Press: Cambridge, UK, 2001; pp. 6–28. [Google Scholar]
- Defarge, N.; Otto, M.; Hilbeck, A.A. Roundup herbicide causes high mortality and impairs development of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Sci. Total Environ. 2023, 865, 161158. [Google Scholar] [CrossRef] [PubMed]
- Kabissa, J.C.B.; Yarro, J.G.; Kayumbo, H.Y.; Juliano, S.A. Functional responses of two chrysopid predators feeding on Helicoverpa armigera (lep.: Noctuidae) and Aphis Gossypii (hom.: Aphididae). Entomophaga 1996, 41, 141–151. [Google Scholar] [CrossRef]
- El-Wakeil, N.M.E. New Aspects of Biological Control of Helicoverpa armigera in Organic Cotton Production. Ph.D. Dissertation, Cairo University, Cairo, Egypt, 2003; p. 140. [Google Scholar] [CrossRef]
- Parajulee, M.N.; Shrestha, R.B.; Leser, J.F.; Wester, D.B.; Blanco, C.A. Evaluation of the functional response of selected arthropod predators on bollworm eggs in the laboratory and effect of temperature on their predation efficiency. Environ. Entomol. 2006, 35, 379–386. [Google Scholar] [CrossRef]
- El-Dessouki, W.A.E.W.; Korish, S.K. Assessment of two biological control agents and insecticide on Spodoptera littoralis (Boisd.) under laboratory conditions. Egypt. J. Crop Prot. 2023, 18, 14–23. [Google Scholar] [CrossRef]
- Hassanpour, M.; Mohaghegh, J.; Iranipour, S.; Nouri-Ganbalani, G.; Enkegaard, A. Functional response of Chrysoperla carnea (Neuroptera: Chrysopidae) to Helicoverpa armigera (Lepidoptera: Noctuidae): Effect of prey and predator stages. Insect Sci. 2011, 18, 217–224. [Google Scholar] [CrossRef]
- Medina, P.; Smagghe, G.; Budia, F.; Del Estal, P.; Tirry, L.; Viñuela, E. Significance of penetration, excretion, and transovarial uptake to toxicity of three insect growth regulators in predatory lacewing adults. Arch. Insect Biochem. Physiol. 2002, 51, 91–101. [Google Scholar] [CrossRef]
- Turquet, M.; Pommier, J.J.; Piron, M.; Lascaux, E.; Lorin, G. Biological control of aphids with Chrysoperla carnea on strawberry. Acta Hortic. 2009, 842, 641–644. [Google Scholar] [CrossRef]
- Devine, G.; Furlong, M. Insecticide use: Contexts and ecological consequences. Agric. Hum. Values 2007, 24, 281–306. [Google Scholar] [CrossRef]
- Wilson, C.; Tisdell, C. Why farmers continue to use pesticides despite environmental, health and sustainability Costs. Ecol. Econ. 2001, 39, 449–462. [Google Scholar] [CrossRef]
- Desneux, N.; Denoyelle, R.; Kaiser, L. A multi-step bioassay to assess the effect of the deltamethrin on the parasitic wasp Aphidius ervi. Chemosphere 2006, 65, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cui, B.; Zhao, X.; Wang, Y.; Zeng, Z.; Sun, C.; Yang, D.; Liu, G.; Cui, H. Optimization and characterization of lambda-cyhalothrin solid nanodispersion by self-dispersing method. Pest Manag. Sci. 2019, 75, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Shu, B.; Hu, M.; Wang, Z.; Zhong, G. Sublethal effects of cyantraniliprole on the development and reproduction of the Cabbage cutworm, Spodoptera litura. J. South China Agric. Univ. 2014, 35, 64–68. [Google Scholar]
- Yu, H.; Xiang, X.; Yuan, G.; Chen, Y.; Wang, X. Effects of sublethal doses of cyantraniliprole on the growth and development and the activities of detoxifying enzymes in Spodoptera exigua (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2015, 6, 634–641. [Google Scholar]
- Moustafa, M.A.M.; Fouad, E.A.; Yasmin, A.M.; Hamow, K.Á.A.; Mikó, Z.; Molnár, B.P.; Fónagy, A. Toxicity and sublethal effects of chlorantraniliprole and indoxacarb on Spodoptera littoralis (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2021, 56, 115–124. [Google Scholar] [CrossRef]
- Moustafa, M.A.M.; Fouad, E.A.; Ibrahim, E.; Erdei, A.L.; Kárpáti, Z.; Fónagy, A. The comparative toxicity, biochemical and physiological impacts of chlorantraniliprole and indoxacarb on Mamestra brassicae (Lepidoptera: Noctuidae). Toxics 2023, 11, 212. [Google Scholar] [CrossRef]
- Hill, M.P.; Macfadyen, S.; Nash, M.A. Broad spectrum pesticide application alters natural enemy communities and may facilitate secondary pest outbreaks. PeerJ 2017, 5, e4179. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Y.; Wangyang, W.; Liu, F.; Cui, Y.; Duan, Y. Design, synthesis, and insecticidal activities of phthalamides containing a hydrazone substructure. J. Agric. Food Chem. 2010, 58, 6858–6863. [Google Scholar] [CrossRef]
- Foster, S.P.; Denholm, I.; Rison, J.L.; Portillo, H.E.; Margaritopoulis, J.; Slater, R. Susceptibility of standard clones and European field populations of the green peach aphid, Myzus persicae, and the cotton aphid, Aphis gossypii (Hemiptera: Aphididae), to the novel anthranilic diamide insecticide cyantraniliprole. Pest Manag. Sci. 2012, 68, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Kandil, M.A.; Moustafa, M.A.M.; Saleh, M.A.; Ateya, I.R. Dissipation kinetics and degradation products of cyantraniliprole in tomato plants and soil in the open field. Egypt. J. Chem. 2023, 66, 483–493. [Google Scholar] [CrossRef]
- Lahm, G.P.; Cordova, D.; Barry, J.D. New and selective ryanodine receptor activators for insect control. Bioorg. Med. Chem. Lett. 2009, 17, 4127–4133. [Google Scholar] [CrossRef] [PubMed]
- Kandil, M.A.; Abdel-kerim, R.N.; Moustafa, M.A.M. Lethal and sublethal effects of bio-and chemical insecticides on the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Egypt. J. Biol. Pest Control 2020, 30, 76. [Google Scholar] [CrossRef]
- Awad, M.; Hassan, N.N.; Alfuhaid, N.A.; Amer, A.; Salem, M.Z.M.; Fónagy, A.; Moustafa, M.A.M. Insecticidal and biochemical impacts with molecular docking analysis of three Essential Oils against Spodoptera littoralis (Lepidoptera: Noctuidae). Crop Prot. 2024, 180, 106659. [Google Scholar] [CrossRef]
- He, F.; Sun, S.; Tan, H.; Sun, X.; Qin, C.; Ji, S.; Li, X.; Zhang, J.; Jiang, X. Chlorantraniliprole against the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae): From biochemical/physiological to demographic responses. Sci. Rep. 2019, 9, 10328. [Google Scholar] [CrossRef]
- Moustafa, M.A.M.; Awad, M.; Amer, A.; Hassan, N.N.; Ibrahim, E.S.; Ali, H.M.; Akrami, M.; Salem, M.Z.M. Insecticidal activity of lemongrass essential oil as an ecofriendly agent against the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae). Insects 2021, 12, 737. [Google Scholar] [CrossRef]
- Moustafa, M.A.M.; Hassan, N.N.; Alfuhaid, N.A.; Amer, A.; Awad, M. Insights into the toxicity, biochemical activity and molecular docking of Cymbopogon citratus essential oils and citral on the cotton leafworm Spodoptera littoralis. J. Econ. Entomol. 2023, 116, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Moustafa, M.M.A.; Kákai, A.; Awad, M.; Fónagy, A. Sublethal effects of spinosad and emamectin benzoate on larval development and reproductive activities of the cabbage moth, Mamestra brassicae L. (Lepidoptera: Noctuidae). Crop Prot. 2016, 90, 197–204. [Google Scholar] [CrossRef]
- Vogt, H.; Brown, K.; Candolfi, M.; Kühner, C.; Moll, M.; Travis, A.; Ufer, A.; Waldburger, M.; Waltersdorfer, A.; Bigler, F. Laboratory method to test effects of plant protection products on larvae of Chrysoperla carnea (Neuroptera: Chrysopidae). In Guidelines to Evaluate Side-Effects of Plant Protection Products to Non-Target Arthropods; IOBC/WPRS: Gent, Belgium, 2000; pp. 27–44. [Google Scholar]
- van Asperen, K. A study of housefly esterases by means of a sensitive colorimetric method. J. Insect Physiol. 1962, 8, 401–416. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar] [CrossRef]
- Walgenbach, J.F.; Bilbo, T.R.; Tussey, D.A.; Ogburn, E.C. Comparison of chemigation versus foliar insecticide use: Management of lepidopteran larvae and stink bugs in North Carolina field tomatoes with environmental and farmworker benefits. Pest Manag. Sci. 2021, 77, 58–765. [Google Scholar] [CrossRef]
- Bilbo, T.R.; Owens, D.R.; Golec, J.R.; Walgenbach, J.F. Impact of insecticide programs on pests, the predatory mite Phytoseiulus persimilis, and staked tomato profitability. Pest Manag. Sci. 2022, 78, 2390–2397. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.H.A.; Mohammed, S.H.; Eltaly, R.I.; Moustafa, M.A.M.; Fónagy, A.; Farag, S.M. Co-application of entomopathogenic fungi with chemical insecticides against Culex pipiens. J. Invertebr. Pathol. 2023, 198, 107916. [Google Scholar] [CrossRef] [PubMed]
- Farag, S.M.; Moustafa, M.A.M.; Fónagy, A.; Kamel, O.M.H.M.; Abdel-Haleem, D.R. Chemical composition of four essential oils and their adulticidal, repellence, and field oviposition deterrence activities against Culex pipiens L. (Diptera: Culicidae). Parasitol. Res. 2024, 123, 110. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.M.; Moteleb, R.I.A.; Ghoneim, Y.F.; Hafez, S.S.; Ali, R.E.; Eweis, E.E.A.; Hassan, N.N. Monitoring resistance and biochemical studies of three Egyptian field strains of Spodoptera littoralis (Lepidoptera: Noctuidae) to six insecticides. Toxics 2023, 11, 211. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, A.M.N.; Faroni, L.R.D.; de Oliveira, J.V.; Navarro, D.M.D.F.; Barbosa, D.R.S.; Breda, M.O.; de Franca, S.M. Lethal and sublethal responses of Sitophilus zeamais populations to essential oils. J. Pest Sci. 2017, 90, 589–600. [Google Scholar] [CrossRef]
- Hannig, G.T.; Ziegler, M.; Marçon, P.G. Feeding cessation effects of chlorantraniliprole, a new anthranilic diamide insecticide, in comparison with several insecticides in distinct chemical classes and mode-of-action groups. Pest Manag. Sci. 2009, 65, 969–974. [Google Scholar] [CrossRef]
- Han, W.; Zhang, S.; Shen, F.; Liu, M.; Ren, C.; Gao, X. Residual toxicity and sublethal effects of chlorantraniliprole on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2012, 68, 1184–1190. [Google Scholar] [CrossRef]
- Song, Y.; Dong, J.; Sun, H. Chlorantraniliprole at sublethal concentrations may reduce the population growth of the Asian corn borer, Ostrinia furnacalis (Lepidoptera: Pyralidae). Acta Entomol. Sin. 2013, 56, 446–451. [Google Scholar] [CrossRef]
- Kinareikina, A.; Silivanova, E. Impact of insecticides at sublethal concentrations on the enzyme activities in Adult Musca domestica L. Toxics 2023, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, X.; Liu, C.; Wen, S.; Xue, Y.; Jin, Y.; Zhang, G.; Xia, X. Effects of sublethal concentrations of cyantraniliprole on the biology and metabolic enzyme activities of Laodelphax striatellus (Fallén). Crop Prot. 2022, 156, 105964. [Google Scholar] [CrossRef]
- Rumpf, S.; Hetzel, F.; Frampton, C. Lacewings (Neuroptera: Hemerobiidae and Chrysopidae) and integrated pest management: Enzyme activity as biomarker of sublethal insecticide exposure. J. Econ. Entomol. 1997, 90, 102–108. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Luo, Y.; Sheikh, U.A.A.; Bamisile, B.S.; Khan, M.M.; Imran, M.; Hafeez, M.; Ghani, M.I.; Lei, N.; Xu, Y. Transcriptome analysis reveals differential effects of beta-cypermethrin and fipronil insecticides on detoxification mechanisms in Solenopsis invicta. Front. Physiol. 2022, 13, 1018731. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Fan, Z.; Sun, Z.; Chen, Y.; Gui, F. Insecticide susceptibility and detoxification enzyme activity of Frankliniella occidentalis under three habitat conditions. Insects 2023, 14, 643. [Google Scholar] [CrossRef]
- Zeng, X.; Pan, Y.; Tian, F.; Li, J.; Xu, H.; Liu, X.; Chen, X.; Gao, X.; Peng, T.; Bi, R.; et al. Functional validation of key cytochrome P450 monooxygenase and UDP-glycosyltransferase genes conferring cyantraniliprole resistance in Aphis gossypii Glover. Pestici. Biochem. Physiol. 2021, 176, 104879. [Google Scholar] [CrossRef]
- Pengsook, A.; Tharamak, S.; Keosaeng, K.; Koul, O.; Bullangpoti, V.; Kumrungsee, N.; Pluempanupat, W. Insecticidal and growth inhibitory effects of some thymol derivatives on the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae) and their impact on detoxification enzymes. Pest Manag. Sci. 2022, 78, 684–691. [Google Scholar] [CrossRef]
- Vandervoet, T.F.; Ellsworth, P.C.; Carriere, Y.; Naranjo, S.E. Quantifying conservation biological control for management of Bemisia tabaci (Hemiptera: Aleyrodidae) in cotton. J. Econ. Entomol. 2018, 111, 1056–1068. [Google Scholar] [CrossRef]
- Bordini, I.; Ellsworth, P.C.; Naranjo, S.E.; Fournier, A. Novel insecticides and generalist predators support conservation biological control in cotton. Biol. Control 2021, 154, 104502. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.A.; Lacerda, M.C.; Zanuncio, T.V.; Ramalho, F.d.S.; Polanczyk, R.A.; Serrão, J.E.; Zanuncio, J.C. Effect of the insect growth regulator diflubenzuron on the predator Podisus nigrispinus (Heteroptera: Pentatomidae). Ecotoxicology 2012, 21, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Shankarganesh, K.; Naveen, N.C.; Bishwajeet, P. Effect of Insecticides on Different Stages of Predatory Green Lacewing, Chrysoperla zastrowi sillemi (Esben.-Petersen). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1415–1422. [Google Scholar] [CrossRef]
- Ham, E.; Lee, J.S.; Jang, M.Y.; Park, J.K. Toxic effects of 12 pesticides on green lacewing, Chrysoperla nipponensis (Okamoto) (Neuroptera: Chrysopidae). Entomol. Res. 2019, 49, 305–312. [Google Scholar] [CrossRef]
- Golmohammadi, G.; Torshizi, H.-R.R.; Vafaei-Shooshtari, R.; Faravardeh, L.; Rafaei-Karehroudi, Z. Lethal and sublethal effects of three insecticides on first instar larvae of green lacewing, Chrysoperla carnea, Stephens. Int. J. Trop. Insect Sci. 2021, 41, 2351–2359. [Google Scholar] [CrossRef]
- Hsin, C. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates among Individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Fergani, Y.A.; Elbanna, H.M.; Hamama, H.M. Genotoxicity of some plant essential oils in cotton leafworm Spodoptera littoralis (Lepidoptera: Noctuidae): The potential role of detoxification enzymes. Egypt. J. Zool. 2020, 73, 53–66. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Z.; Abou-Zaid, M.M.; Arnason, J.T.; Liu, R.; Walshe-Roussel, B.; Waye, A.; Liu, S.; Saleem, A.; Caceres, L.A.; et al. Inhibition of insect glutathione S-transferase (GST) by conifer extracts. Arch. Insect Biochem. Physiol. 2014, 87, 234–249. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).