


Unconventional Strategies for Aphid Management in Sorghum


Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Site Location
2.2. Insecticide Applications
2.3. Field Design and Treatments
2.4. Non-Natural Infestations of Sorghum Aphid Populations
2.5. Insect Sampling
2.6. Cumulative Aphid Days and Sprayed Plants
2.7. Yield Sampling
2.8. Data Processing
2.9. Statistical Analysis
3. Results
3.1. Aphid Abundance over Time in the 2021 Season
3.2. Aphid Abundance over Time in the 2022 Season
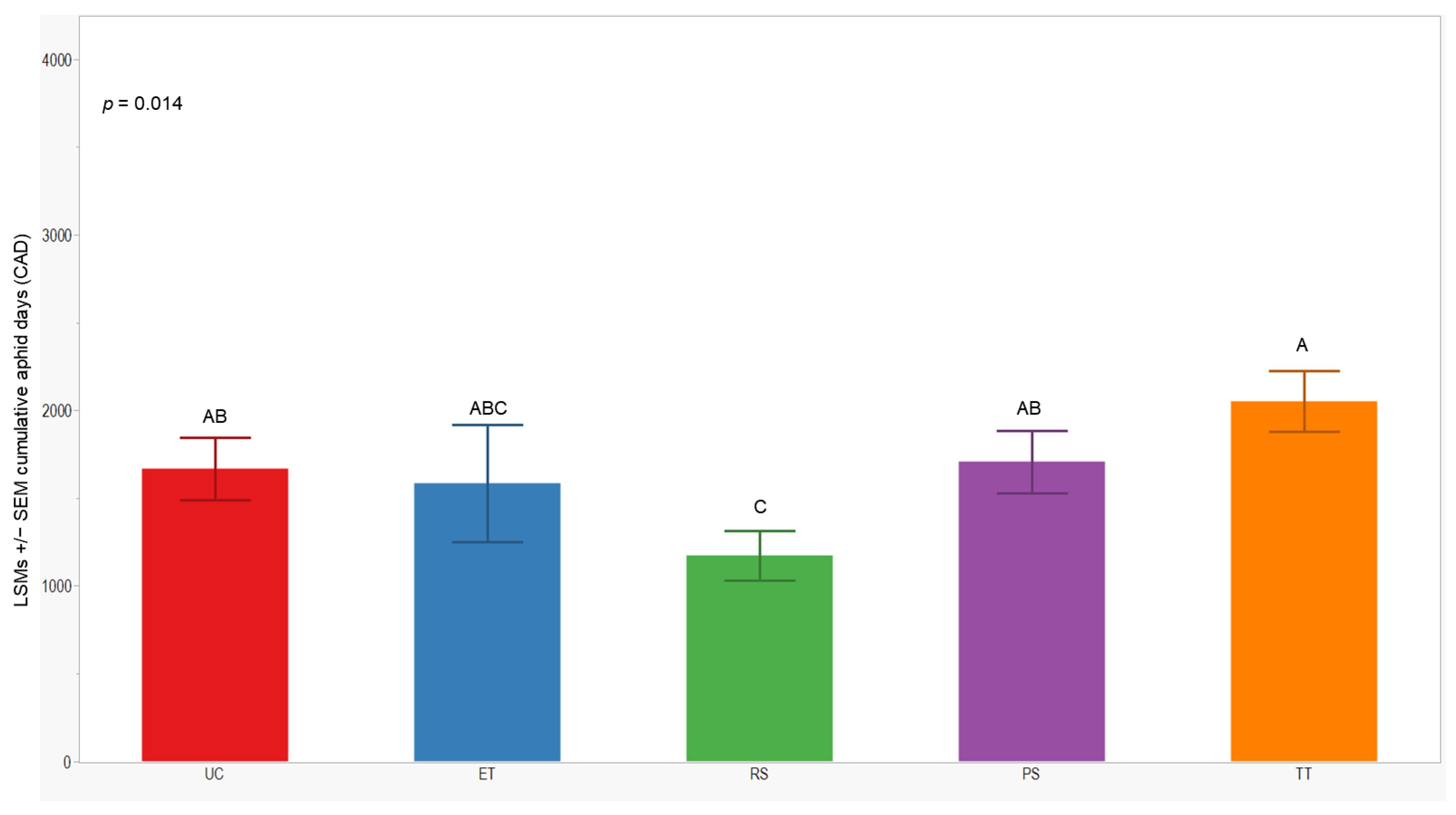
3.3. Cumulative Aphid Days
3.4. Cumulative Sprayed Plants and Grain Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mandava, N.B. Handbook of Natural Pesticides: Methods: Volume I: Theory, Practice, and Detection; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-1-351-08961-6. [Google Scholar]
- Kogan, M. Integrated Pest Management: Historical Perspectives and Contemporary Developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Flint, M.L.; Van den Bosch, R. Introduction to Integrated Pest Management; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 1-4615-9212-7. [Google Scholar]
- Pimentel, D.; Levitan, L. Pesticides: Amounts Applied and Amounts Reaching Pests. BioScience 1986, 36, 86–91. [Google Scholar] [CrossRef]
- Krupke, C.; Long, E. Intersections between Neonicotinoid Seed Treatments and Honey Bees. Curr. Opin. Insect Sci. 2015, 10, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Krupke, C.; Holland, J.D.; Long, E.Y.; Eitzer, B.D. Planting of Neonicotinoid-Treated Maize Poses Risks for Honey Bees and Other Non-Target Organisms over a Wide Area without Consistent Crop Yield Benefit. J. Appl. Ecol. 2017, 54, 1449–1458. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental Fate and Exposure; Neonicotinoids and Fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.E. Insecticide Resistance in the Western Flower Thrips, Frankliniella occidentalis. Integr. Pest Manag. Rev. 2000, 5, 131–146. [Google Scholar] [CrossRef]
- Kranthi, K.R.; Jadhav, D.R.; Kranthi, S.; Wanjari, R.R.; Ali, S.S.; Russell, D.A. Insecticide Resistance in Five Major Insect Pests of Cotton in India. Crop Prot. 2002, 21, 449–460. [Google Scholar] [CrossRef]
- Margni, M.; Rossier, D.; Crettaz, P.; Jolliet, O. Life Cycle Impact Assessment of Pesticides on Human Health and Ecosystems. Agric. Ecosyst. Environ. 2002, 93, 379–392. [Google Scholar] [CrossRef]
- Van Lenteren, J.C. The State of Commercial Augmentative Biological Control: Plenty of Natural Enemies, but a Frustrating Lack of Uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Brewer, M.J.; Peairs, F.B.; Elliott, N.C. Invasive Cereal Aphids of North America: Ecology and Pest Management. Annu. Rev. Entomol. 2019, 64, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Haar, P.J.; Buntin, G.D.; Jacobson, A.; Pekarcik, A.; Way, M.O.; Zarrabi, A. Evaluation of Tactics for Management of Sugarcane Aphid (Hemiptera: Aphididae) in Grain Sorghum. J. Econ. Entomol. 2019, 112, 2719–2730. [Google Scholar] [CrossRef] [PubMed]
- Ciampitti, I.; Ruiz, D.; Onofre, R.; Jardine, R.; Lancaster, S.; Whitworth, J.; Aguilar, J. Kansas Sorghum Management 2021. 2021. Available online: https://bookstore.ksre.ksu.edu/pubs/MF3046.pdf (accessed on 4 March 2024).
- Singh, B.U.; Padmaja, P.G.; Seetharama, N. Biology and Management of the Sugarcane Aphid, Melanaphis sacchari (Zehntner) (Homoptera: Aphididae), in Sorghum: A Review. Crop Prot. 2004, 23, 739–755. [Google Scholar] [CrossRef]
- Bowling, R.D.; Brewer, M.J.; Kerns, D.L.; Gordy, J.; Seiter, N.; Elliott, N.E.; Buntin, G.D.; Way, M.O.; Royer, T.A.; Biles, S.; et al. Sugarcane Aphid (Hemiptera: Aphididae): A New Pest on Sorghum in North America. J. Integr. Pest Manag. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- McCornack, B.; Zukoff, S.; Michaud, J.; Whitworth, J. Scouting Sugarcane Aphids. Available online: https://www.myfields.info/sites/default/files/page/ScoutCard%20KSU%20v05312017.pdf (accessed on 10 November 2023).
- Gordy, J.W.; Brewer, M.J.; Bowling, R.D.; Buntin, G.D.; Seiter, N.J.; Kerns, D.L.; Reay-Jones, F.P.F.; Way, M.O. Development of Economic Thresholds for Sugarcane Aphid (Hemiptera: Aphididae) in Susceptible Grain Sorghum Hybrids. J. Econ. Entomol. 2019, 112, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.W.; Radcliffe, E.B.; MacRae, I.V.; Ragsdale, D.W.; Olson, K.D.; Badibanga, T. Border Treatment to Reduce Insecticide Use in Seed Potato Production: Biological, Economic, and Managerial Analysis. Am. J. Potato Res. 2008, 86, 31. [Google Scholar] [CrossRef]
- Lindenmayer, J.C.; Giles, K.L.; Elliott, N.C.; Knutson, A.E.; Bowling, R.; Brewer, M.J.; Seiter, N.J.; McCornack, B.; Brown, S.A.; Catchot, A.L.; et al. Development of Binomial Sequential Sampling Plans for Sugarcane Aphid (Hemiptera: Aphididae) in Commercial Grain Sorghum. J. Econ. Entomol. 2020, 113, 1990–1998. [Google Scholar] [CrossRef]
- Bowling, R.; Brewer, M.; Knutson, A.; Biles, S.; Way, M.; Sekula-Ortiz, D. Scouting Sugarcane Aphids in South, Central, and West Texas; Texas A&M AgriLife Research: Weslaco, TX, USA, 2021; p. 2. Available online: http://agrilife.org/sca/files/2016/06/Sugarcane-Aphid-Quick-Checker.pdf (accessed on 10 March 2024).
- Lytle, A.J.; Huseth, A.S. Impact of Foliar Insecticide Sprays on Melanaphis sacchari (Hemiptera: Aphididae) and Natural Enemy Populations in Grain Sorghum. Crop Prot. 2021, 149, 105764. [Google Scholar] [CrossRef]
- Zukoff, A.; McCornack, B.; Whitworth, R. Sorghum Insect Pest Management. Available online: https://bookstore.ksre.ksu.edu/pubs/MF742.pdf (accessed on 15 January 2024).
- Brewer, M.J.; Gordy, J.W.; Kerns, D.L.; Woolley, J.B.; Rooney, W.L.; Bowling, R.D. Sugarcane Aphid Population Growth, Plant Injury, and Natural Enemies on Selected Grain Sorghum Hybrids in Texas and Louisiana. J. Econ. Entomol. 2017, 110, 2109–2118. [Google Scholar] [CrossRef]
- Pekarcik, A.J.; Jacobson, A.L. Evaluating Sugarcane Aphid, Melanaphis sacchari (Hemiptera: Aphididae), Population Dynamics, Feeding Injury, and Grain Yield among Commercial Sorghum Varieties in Alabama. J. Econ. Entomol. 2021, 114, 757–768. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Abbreviation | Treatment Name | Treatment Description and CSP |
|---|---|---|
| UC | Untreated control | No insecticide applications were made. No plants were sprayed. |
| ET | Economic threshold | All plants in the two middle rows of the plot were sprayed based on an economic threshold of ≥40 mean sorghum aphid wingless/leaf on 12 randomly evaluated plants. A CSP of 2588 was sprayed. |
| RS | Randomly sprayed | Consisted of the same number of plants treated as in PS but randomly sprayed plants regardless of sorghum aphid wingless presence. A CSP of 612 was sprayed. |
| PS | Plant-specific | Consisted of specific plants sprayed upon visually confirming the presence of wingless (≥1 sorghum aphid wingless/leaf) on 12 randomly evaluated plants. A CSP of 612 was sprayed. |
| TT | Tally threshold | Consisted of individual plants sprayed based on an economic threshold per plant basis of ≥40 sorghum aphid wingless/plant on 12 randomly evaluated plants. A CSP of 214 was sprayed. |
| p-Value for Type III Test of Fixed Effect | 2021 Season Wingless Aphids | 2022 Season Wingless Aphids |
|---|---|---|
| Evaluation baseline | 0.492 | 0.285 |
| Block | 0.018 | 0.452 |
| Treatment (Trt) | 0.068 | <0.001 |
| Evaluation phase (EP) | <0.001 | <0.001 |
| Sample day (D) | <0.001 | <0.001 |
| Trt × EP | <0.001 | <0.001 |
| Trt × D | 0.098 | 0.113 |
| EP × D | <0.001 | <0.001 |
| Trt × EP × D | <0.001 | 0.288 |
| Treatment Abbreviation | Treatment Name | 2021 Season CSP | 2022 Season CSP |
|---|---|---|---|
| UC | Untreated control | - | - |
| ET | Economic threshold | 1388 | 1200 |
| RS | Randomly sprayed | 280 | 332 |
| PS | Plant-specific | 280 | 332 |
| TT | Tally threshold | 92 | 122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grijalva, I.; Kang, Q.; Flippo, D.; Sharda, A.; McCornack, B. Unconventional Strategies for Aphid Management in Sorghum. Insects 2024, 15, 475. https://doi.org/10.3390/insects15070475
Grijalva I, Kang Q, Flippo D, Sharda A, McCornack B. Unconventional Strategies for Aphid Management in Sorghum. Insects. 2024; 15(7):475. https://doi.org/10.3390/insects15070475
Chicago/Turabian StyleGrijalva, Ivan, Qing Kang, Daniel Flippo, Ajay Sharda, and Brian McCornack. 2024. "Unconventional Strategies for Aphid Management in Sorghum" Insects 15, no. 7: 475. https://doi.org/10.3390/insects15070475
APA StyleGrijalva, I., Kang, Q., Flippo, D., Sharda, A., & McCornack, B. (2024). Unconventional Strategies for Aphid Management in Sorghum. Insects, 15(7), 475. https://doi.org/10.3390/insects15070475