
Direct Observation of Feeding Behavior of Adult Tabanidae (Diptera) on Beef Cattle from Khon Kaen Province in Thailand

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Cattle
2.2. Tabanid Landings
2.3. Statistical Analysis
3. Results
3.1. Landing Sites of Tabanids on the Cattle
3.2. Time Period and Meteorological Factors of Tabanids Landing on Host
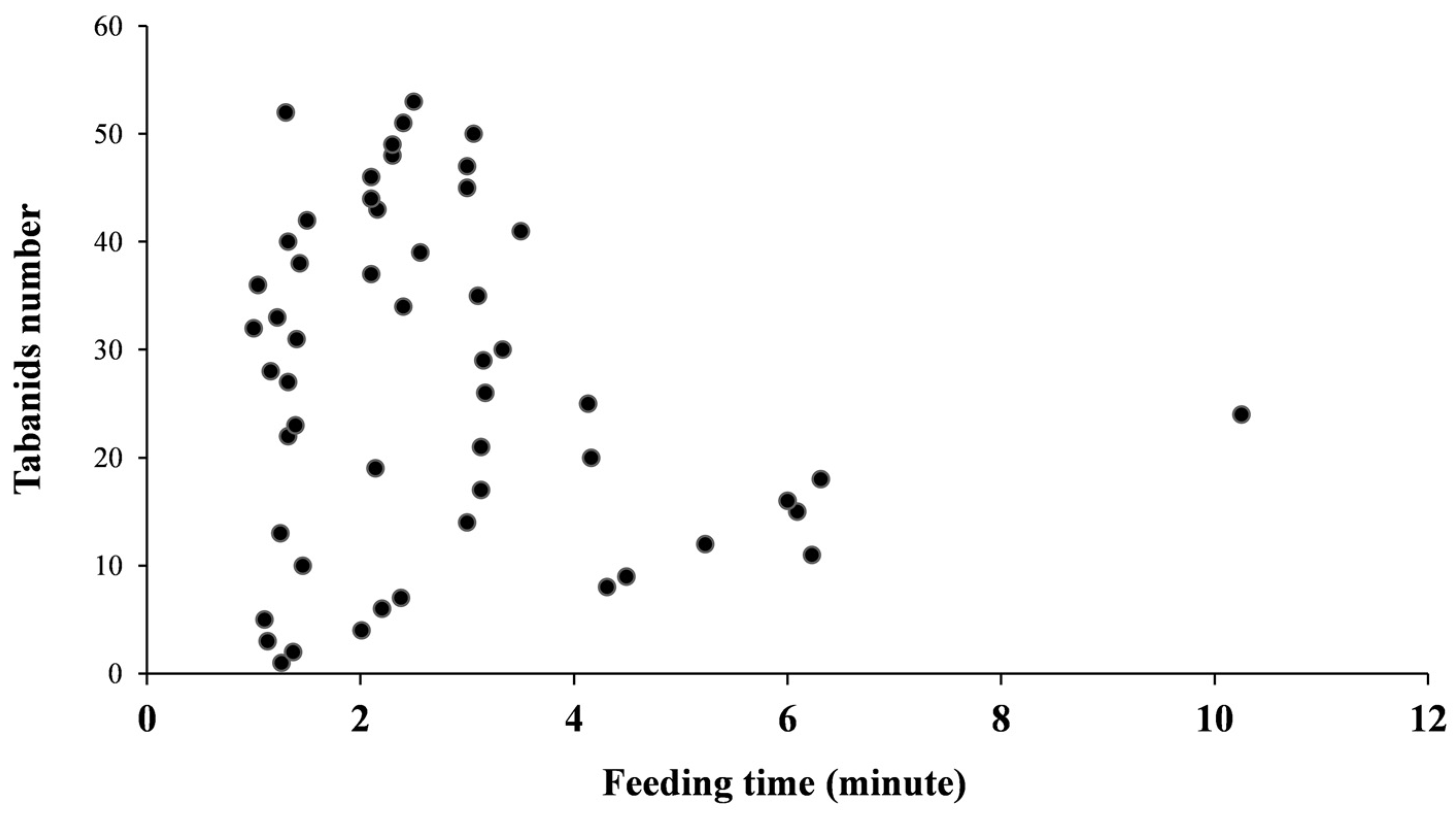
3.3. Feeding Duration
4. Discussion
4.1. Tabanid Landing Sites on the Cattle
4.2. Time of Day and Meteorological Factors of Tabanids Landing on Host
4.3. Feeding Duration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldacchino, F.; Desquesnes, M.; Mihok, S.; Foil, L.D.; Duvallet, G.; Jittapalapong, S. Tabanids: Neglected subjects of research, but important vectors of disease agents! Infect. Genet. Evol. 2014, 28, 596–615. [Google Scholar] [CrossRef]
- Morita, S.I.; Bayless, K.M.; Yeates, D.K.; Wiegmann, B.M. Molecular phylogeny of the horse flies: A framework for renewing tabanid taxonomy. Syst. Entomol. 2016, 41, 56–72. [Google Scholar] [CrossRef]
- Coscarón, S.; Philip, C.B. A revision of Mycteromyiini (‘genus Mycteomyia’of authors), a new tribe of Neotropical horse flies (Diptera, Tabanidae). Proc. Natl. Acad. Sci. USA 1979, 41, 427–452. [Google Scholar]
- Lessard, B.D.; Cameron, S.L.; Bayless, K.M.; Wiegmann, B.M.; Yeates, D.K. The evolution and biogeography of the austral horse fly tribe Scionini (Diptera: Tabanidae: Pangoniinae) inferred from multiple mitochondrial and nuclear genes. Mol. Phylogenetics Evol. 2013, 68, 516–540. [Google Scholar] [CrossRef] [PubMed]
- Mackerras, I.M. Classification and distribution of Tabanidae (Diptera). Aust. J. Zool. 1954, 2, 431–454. [Google Scholar] [CrossRef]
- Mackerras, I.M. The classification and distribution of tananidae (Diptera) 2. History: Morphology: Classification: Subfamily Pangoniinae. Aust. J. Zool. 1955, 3, 439–511. [Google Scholar] [CrossRef]
- Stone, A. Family Tabanidae. A Catalog of the Diptera of the Oriental Region. Suborder Brachycera Vol. 2, 2nd ed.; Delfinado, M.D., Hardy, D.E., Eds.; University Press of Hawaii: Honolulu, HI, USA, 1975; pp. 43–81. [Google Scholar]
- Burton, J.J. Tabanini of Thailand above the Isthmus of Kra (Diptera: Tabanidae); Entomological Reprint Specialists; Smithsonian Libraries and Archives: Washington, DC, USA, 1978; p. 165. [Google Scholar]
- Phasuk, J.; Tharawoot, T.; Beaver, R.A.; Jittapalapong, S. Seasonal abundance of Tabanidae (Diptera) on dairy farms in Saraburi Province, Thailand. Thai J. Agric. Sci. 2011, 44, 175–181. [Google Scholar]
- Sontigun, N.; Boonhoh, W.; Phetcharat, Y.; Wongtawan, T. First study on molecular detection of hemopathogens in tabanid flies (Diptera: Tabanidae) and cattle in Southern Thailand. Vet. World 2022, 15, 2089. [Google Scholar] [CrossRef]
- Ramos, C.J.R.; de Souza Franco, C.; da Luz, S.P.; Marques, J.; de Souza, K.M.; do Nascimento, L.F.N.; das Neves, G.B.; Moreira, R.S.; Miletti, L.C. First record of Trypanosoma evansi DNA in Dichelacera alcicornis and Dichelacera januarii (Diptera: Tabanidae) flies in South America. Parasit. Vectors 2023, 16, 4. [Google Scholar] [CrossRef]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical lumpy skin disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Ndzeshang, L.B.; Fombad, F.F.; Njouendou, A.J.; Chunda, V.C.; Gandjui, N.V.T.; Akumtoh, D.N.; Chounna, P.W.N.; Steven, A.; Pionnier, N.P.; Layland, L.E.; et al. Generation of Loa loa infective larvae by experimental infection of the vector, Chrysops silacea. PLoS Negl. Trop. Dis. 2020, 14, e0008415. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.P.; Mohd Rajdi, N.Z.I.; Mohamad, M.A.; Mohamed, M.; Hamdan, R.H.; Goriman Khan, M.A.K.; Ahmad Syazwan, S.; Seng Hua, L. First Report of Trypanosoma theileri in Equine Host and Tabanus sp. in Malaysia. J. Equine Vet. Sci. 2022, 108, 103807. [Google Scholar] [CrossRef] [PubMed]
- Office of Agricultural Economics; Export Statistics of Male and Female Cattle Another Sentence. Available online: http://impexp.oae.go.th/service/export.php (accessed on 20 May 2024).
- Department of Research and Strategy, National Food Institute (Thailand). Available online: https://www.foodfocusthailand.com/whats-in/current-situation-and-direction-of-the-thai-beef-cattle-industry (accessed on 20 May 2024).
- Duvallet, G.; Fontenille, D.; Robert, V. (Eds.) Entomologie Médicale et Vétérinaire, 1st ed.; IRD Éditions, Éditions Quæ: Marseille, France, 2017; pp. 165–179. [Google Scholar]
- Mulandane, F.C.; Snyman, L.P.; Brito, D.R.A.; Bouyer, J.; Fafetine, J.; Van Den Abbeele, J.; Oosthuizen, M.; Delespaux, V.; Neves, L. Evaluation of the relative roles of the Tabanidae and Glossinidae in the transmission of trypanosomosis in drug resistance hotspots in Mozambique. Parasites Vectors 2020, 13, 219. [Google Scholar] [CrossRef]
- Desquesnes, M.; Thaisungnoen, K.; Chalermwong, P.; Nevot, A.; Fossaert, C.; Lucas, A.; Onju, S.; Boonsaen, P.; Jittapalapong, S. The Use of “Tail-Pedometers” to Evaluate the Impact of Dipterans in Feeder Cattle. Insects 2022, 13, 616. [Google Scholar] [CrossRef] [PubMed]
- Muzari, M.O.; Skerratt, L.F.; Jones, R.E.; Duran, T.L. Alighting and feeding behaviour of tabanid flies on horses, kangaroos and pigs. Vet. Parasitol. 2010, 170, 104–111. [Google Scholar] [CrossRef]
- Brady, J.; Shereni, W. Landing responses of tsetse fly Glossina mor- sitans morsitans Westwood and the stable fly Stomoxys calcitrans (L.) (Diptera: Glossinidae & Muscidae) to black-and-white patterns: A laboratory study. Bull. Entomol. Res. 1988, 78, 301–311. [Google Scholar]
- Baldacchino, F.; Puech, L.; Manon, S.; Hertzog, L.R.; Jay-Robert, P. Biting behaviour of Tabanidae on cattle in mountainous summer pastures, Pyrenees, France, and effects of weather variables. Bull. Entomol. Res. 2014, 104, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Herczeg, T.; Száz, D.; Blahó, M.; Barta, A.; Gyurkovszky, M.; Farkas, R.; Horváth, G. The effect of weather variables on the flight activity of horseflies (Diptera: Tabanidae) in the continental climate of Hungary. Parasitol. Res. 2015, 114, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Horváth, G.; Pereszlényi, Á.; Tóth, T.; Polgár, S.; Jánosi, I.M. Attractiveness of thermally different, uniformly black targets to horseflies: Tabanus tergestinus prefers sunlit warm shiny dark targets. R. Soc. Open Sci. 2019, 6, 191119. [Google Scholar] [CrossRef]
- Bunmee, T.; Chaiwang, N.; Kaewkot, C.; Jaturasitha, S. Current situation and future prospects for beef production in Thailand—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 968–975. [Google Scholar] [CrossRef]
- Department of Livestock Development; Information on the Number of Livestock in Thailand 2023. Available online: https://region3.dld.go.th (accessed on 12 July 2024).
- Statistix 10; Statistix: Tallahassee, FL, USA, 2015.
- Changbunjong, T.; Sedwisi, P.; Weluwanarak, T.; Nitiyamatawat, E.; Sariwongchan, R.; Chareonviriyaphap, T. Species diversity and abundance of Tabanus spp. (Diptera: Tabanidae) in different habitats of Thailand. J. Asia. Pac. Entomol. 2018, 21, 134–139. [Google Scholar] [CrossRef]
- Raymond, H.L.; Rousseau, F. Abundance of horse flies (Diptera, Tabanidae) and reactions of cattle in a traditional farm of French-Guiana. Acta Oecologica 1987, 8, 125–134. [Google Scholar]
- Mohamed-Ahmed, M.M.; Mihok, S. Alighting of Tabanidae and muscids on natural and simulated hosts in the Sudan. Bull. Entom. Res. 2009, 99, 561–571. [Google Scholar] [CrossRef]
- Lendzele, S.S.; Eisenbarth, A.; Christophe, Z.K.R.; Mavoungou, J.F.; Renz, A. Aspects of the bionomics of hematophagous symbovine dipterans in a hyper-infested rangeland of Ngaoundere (Adamawa-Cameroon). J. Asia. Pac. Entomol. 2019, 22, 1019–1030. [Google Scholar] [CrossRef]
- Krcmar, S.; Marić, S. Analysis of the feeding sites for some horse flies (Diptera, Tabanidae) on a human in Croatia. Coll. Antropol. 2006, 30, 901–904. [Google Scholar]
- Altunsoy, F. Host and Feeding Side Preferences of the Horse Fly (Diptera: Tabanidae) Species. J. Entomol. Res. Soc. 2015, 17, 107–115. [Google Scholar]
- Todd, D.H. The biting fly Stomoxys calcitrans (L.) in dairy herds in New Zealand. N. Zeal. J. Agr. Res. 1964, 7, 60–79. [Google Scholar] [CrossRef]
- Mullens, B.A.; Gerhardt, R.R. Feeding-behavior of some Tennessee Tabanidae. Environ. Entomol. 1979, 8, 1047–1051. [Google Scholar] [CrossRef]
- Waage, J.K.; Davies, C.R. Host-mediated competition in a bloodsucking insect community. J. Anim. Ecol. 1986, 55, 171–180. [Google Scholar] [CrossRef]
- Olalekan Odeniran, P.; Oluwafemi Ademola, I. Alighting and feeding behavior of trypanosome-transmitting vectors on cattle in Nigeria. J. Med. Entomol. 2018, 55, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Miletti, L.C.; Colombo, B.B.; Cardoso, C.P.; Stalliviere, F.M.; Tavares, K.C.S.; Komati, L.K.O.; Vieira, L.L.; Christen, S.E.; Ramos, C.J.R. Prevalence, seasonality and behaviour of Tabanidae (Diptera) captured on a horse in the Planalto Serrano of Santa Catarina State, Brazil. Int. J. Trop. Insect Sci. 2011, 31, 122–126. [Google Scholar] [CrossRef]
- Amano, K. Statistical analyses of the influence of meteorological factors on flight activity of female tabanids. Kontyû 1985, 53, 161–172. [Google Scholar]
- McElligott, P.E.; Galloway, T.D. Daily activity patterns of horse flies (Diptera: Tabanidae: Hybomitra spp.) in northern and southern Manitoba. Can. Entomol. 1991, 123, 371–378. [Google Scholar] [CrossRef]
- Gibson, G.; Torr, S.J. Visual and olfactory responses of haematophagous Diptera to host stimuli. Med. Vet. Entomol. 1999, 13, 2–23. [Google Scholar] [CrossRef]
- Chvala, M.; Hurka, K.; Chalupsky, J.; Knoz, J.; Minar, J.; Orszagh, I. “Krevsajici mouchy a strecci—Diptera.” Fauna CSSR-Ceskoslovenska Akademie ved. Sekce Biologicka 22 Krevsající Mouchy a Strecci—Diptera: Celedi Ceratopogonidae, Simuliidae, Tabanidae, Hypodermatidae, Oestridae, Gasterophilidae, Hippoboscidae a Nycteribiidae; Praha, Academia, Nakl. Ceskoslovenské akademie ved.; Smithsonian Libraries and Archives: Washington, DC, USA, 1980. [Google Scholar]
- Caro, T. Zebra Stripes; University of Chicago Press: Chicago, IL, USA, 2016. [Google Scholar]
- Volfova, V.; Tothova, V.; Volf, P. Hyaluronidase activity in the salivary glands of tabanid flies. Insect Biochem. Mol. Biol. 2016, 73, 38–46. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Body Part | Number of Tabanid Landing (Mean ± SD) | % |
|---|---|---|
| Fore udder | 0.033 ± 0.11 c | 0.17 |
| Tail | 0.167 ± 0.24 bc | 0.84 |
| Navel | 0.133 ± 0.23 bc | 0.68 |
| Leg | 15.067 ± 7.54 a | 76.35 |
| Dewlap | 0.267 ± 0.21 bc | 1.35 |
| Body | 2.83 ± 2.86 b | 14.36 |
| Under | 1.23 ± 1.30 bc | 6.25 |
| Total | 19.727 ± 11.81 | 100 |
| p (0.05) | p < 0.001 | |
| CV (%) | 109.64 |
| Leg | Tabanid (Mean ± SD) | % |
|---|---|---|
| Upper | 2.37 ± 1.76 | 15.70 |
| Lower | 12.70 ± 6.22 | 84.29 |
| p = 0.0004 * | ||
| Fore | 19.7 ± 9.29 | 43.58 |
| Hind | 25.4 ± 13.49 | 56.41 |
| p = 0.1356 NS |
| Time Span | Number of Tabanids Landing on Cattle (Mean ± SD) |
|---|---|
| 9.00–10.30 | 5.03 ± 3.19 b |
| 12.00–13.30 | 11.93 ± 5.88 a |
| 15.30–17.00 | 2.77 ± 1.80 b |
| p value | 0.0001 * |
| CV (%) | 61.30 |
| Temperature (Mean ± SD) | Relative Humidity (Mean ± SD) | |
|---|---|---|
| Morning (09.00–10.30) | 29.22 ± 2.65 | 55.9 ± 12.75 |
| Midday (12.00–13.30) | 31.26 ± 1.92 | 46.2 ± 17.13 |
| Afternoon (16.30–17.00) | 29.88 ± 2.10 | 50.3 ± 15.96 |
| df | SS | MS | F | Significance F | |
|---|---|---|---|---|---|
| Regression | 2 | 710.08 | 355.04 | 11.26 | 0.01 * |
| Residual | 7 | 220.76 | 31.54 | ||
| Total | 9 | 930.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thinnabut, K.; Maleewong, W.; Tangkawanit, U. Direct Observation of Feeding Behavior of Adult Tabanidae (Diptera) on Beef Cattle from Khon Kaen Province in Thailand. Insects 2024, 15, 602. https://doi.org/10.3390/insects15080602
Thinnabut K, Maleewong W, Tangkawanit U. Direct Observation of Feeding Behavior of Adult Tabanidae (Diptera) on Beef Cattle from Khon Kaen Province in Thailand. Insects. 2024; 15(8):602. https://doi.org/10.3390/insects15080602
Chicago/Turabian StyleThinnabut, Kanchana, Wanchai Maleewong, and Ubon Tangkawanit. 2024. "Direct Observation of Feeding Behavior of Adult Tabanidae (Diptera) on Beef Cattle from Khon Kaen Province in Thailand" Insects 15, no. 8: 602. https://doi.org/10.3390/insects15080602