
Special Survival Strategy of First-Instar Scorpions Revealed by Synchronous Molting Behavior from Social Facilitation of Maternal Care and Reciprocal Aggregation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Maintenance
2.2. The Behavior of Newborn Scorpions Climbing onto the Backs of Female Scorpions
2.3. The Molting Behavior of First-Instar Scorpions
2.4. The Aggregating Behavior of First-Instar Scorpions during the Molting Process
2.5. The Interplay between Newborn and Adult Male Scorpions
2.6. Calculation and Statistical Analysis
3. Results
3.1. Unique Feature of Aggregating Molting Behavior in First-Instar Scorpions
3.2. Special Maternal Care Reduced the Time for First-Instar Scorpions to Climb onto Their Mother’s Back before Molting
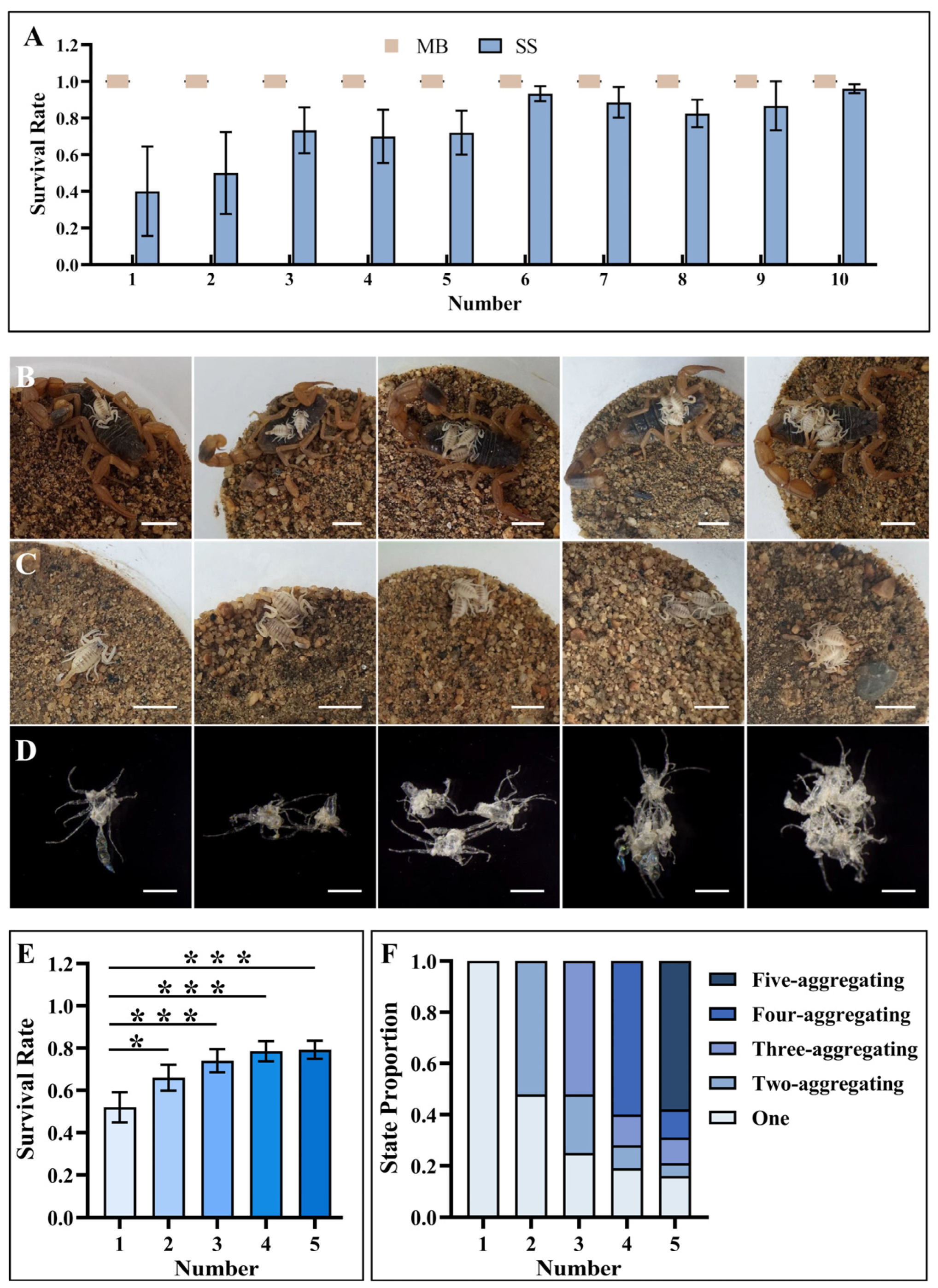
3.3. Maternal Care Increased the Survival Rate of First-Instar Scorpions during the Aggregate Molting Process
3.4. The First-Instar Number Differentially Affected Aggregate Molting Behavior in the Presence/Absence of Maternal Care
3.5. Predatory Behavior of Adult Male Scorpions toward Newborn Scorpions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Minelli, A.; Boxshall, G.; Fusco, G. An Introduction to the Biology and Evolution of Arthropods. In Arthropod Biology and Evolution; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–15. [Google Scholar] [CrossRef]
- Wong, M.K.L.; Guenard, B.; Lewis, O.T. Trait-based ecology of terrestrial arthropods. Biol. Rev. Camb. Philos. Soc. 2019, 94, 999–1022. [Google Scholar] [CrossRef] [PubMed]
- Moussian, B. The Arthropod Cuticle. In Arthropod Biology and Evolution; Springer: Berlin/Heidelberg, Germany, 2013; pp. 171–196. [Google Scholar] [CrossRef]
- Kim, K.W. Social facilitation of synchronized molting behavior in the spider Amaurobius ferox (Araneae, Amaurobiidae). J. Insect Behav. 2001, 14, 401–409. [Google Scholar] [CrossRef]
- Webster, S.G. Seasonal anecdysis and moulting synchrony in field populations of Palaemon elegans (Rathke). Estuar. Coast. Shelf Sci. 1982, 15, 85–94. [Google Scholar] [CrossRef]
- Howe, N.R. Partial molting synchrony in the giant Malaysian prawn, Macrobrachium rosenbergii: A chemical communication hypothesis. J. Chem. Ecol. 1981, 7, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Leinaas, H.P. Synchronized moulting controlled by communication in group-Living Collembola. Science 1983, 219, 193–195. [Google Scholar] [CrossRef]
- Stone, R.P. Mass molting of tanner crabs Chionoecetes bairdi in a Southeast Alaska-Estuary. Alsk. Fish. Res. Bull 1999, 6, 19–28. [Google Scholar] [CrossRef]
- Corrales-García, A.; Esteve, J.; Zhao, Y.; Yang, X. Synchronized moulting behaviour in trilobites from the Cambrian Series 2 of South China. Sci. Rep. 2020, 10, 14099. [Google Scholar] [CrossRef]
- Karim, T.; Westrop, S.R. Taphonomy and Paleoecology of Ordovician Trilobite Clusters, Bromide Formation, South-Central Oklahoma. PALAIOS 2002, 17, 394–402. [Google Scholar] [CrossRef]
- Błażejowski, B.; Brett, C.E.; Kin, A.; Radwański, A.; Gruszczyński, M.; Sevastopulo, G. Ancient animal migration: A case study of eyeless, dimorphic Devonian trilobites from Poland. Palaeontology 2016, 59, 743–751. [Google Scholar] [CrossRef]
- Vannier, J.; Vidal, M.; Marchant, R.; El Hariri, K.; Kouraiss, K.; Pittet, B.; El Albani, A.; Mazurier, A.; Martin, E. Collective behaviour in 480-million-year-old trilobite arthropods from Morocco. Sci. Rep. 2019, 9, 14941. [Google Scholar] [CrossRef]
- Paterson, J.R.; Jago, J.B.; Brock, G.A.; Gehling, J.G. Taphonomy and palaeoecology of the emuellid trilobite Balcoracania dailyi (early Cambrian, South Australia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 249, 302–321. [Google Scholar] [CrossRef]
- Braddy, S.J. Eurypterid palaeoecology: Palaeobiological, ichnological and comparative evidence for a ‘mass–moult–mate’ hypothesis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 172, 115–132. [Google Scholar] [CrossRef]
- Vrazo, M.B.; Braddy, S.J. Testing the ‘mass-moult-mate’ hypothesis of eurypterid palaeoecology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 311, 63–73. [Google Scholar] [CrossRef]
- Haug, J.T.; Caron, J.-B.; Haug, C. Demecology in the Cambrian: Synchronized molting in arthropods from the Burgess Shale. BMC Biol. 2013, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Tetlie, O.E.; Brandt, D.S.; Briggs, D.E.G. Ecdysis in sea scorpions (Chelicerata: Eurypterida). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 265, 182–194. [Google Scholar] [CrossRef]
- Dunlop, J.A.; Erik Tetlie, O.; Prendini, L. Reinterpretation of the Silurian scorpion Proscorpius osborni (Whitfield): Integrating data from Palaeozoic and recent scorpions. Palaeontology 2008, 51, 303–320. [Google Scholar] [CrossRef]
- Waddington, J.; Rudkin, D.M.; Dunlop, J.A. A new mid-Silurian aquatic scorpion-one step closer to land? Biol. Lett. 2015, 11, 20140815. [Google Scholar] [CrossRef]
- Polis, G.A. The Biology of Scorpions; Stanford University Press: Palo Alto, CA, USA, 1991. [Google Scholar]
- Brownell, P.; Polis, G.A. Scorpion Biology and Research; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Polis, G.; Farley, R. Characteristics and environmental determinants of natality, growth and maturity in a natural population of the desert scorpion, Paruroctonus mesaensis (Scorpionida: Vaejovidae). J. Zool. 1979, 187, 517–542. [Google Scholar] [CrossRef]
- Hjelle, J.T. Observations on the birth and post-birth behavior of Syntropis macrura Kraepelin (Scorpionida: Vaejovidae). J. Arachnol. 1973, 1, 221–227. [Google Scholar]
- Ugolini, A.; Carmignani, I.; Vannini, M. Mother-young relationship in Euscorpius: Adaptive value of the larval permanence on the mother’s Back (Scorpiones, Chactidae). J. Arachnol. 1986, 14, 43–46. [Google Scholar]
- Warburg, M.R. Pre- and post-parturial aspects of scorpion reproduction: A review. Eur. J. Entomol. 2012, 109, 139–146. [Google Scholar] [CrossRef]
- Meßlinger, K. Fine structure of scorpion trichobothria (Arachnida, Scorpiones). Zoomorphology 1987, 107, 49–57. [Google Scholar] [CrossRef]
- Lang, W.H.; Marcy, M. Some effects of early starvation on the survival and development of barnacle nauplii, Balanus improvisus (Darwin). J. Exp. Mar. Biol. Ecol. 1982, 60, 63–70. [Google Scholar] [CrossRef]
- Dautel, H.; Knülle, W. The influence of physiological age of Argas reflexus larvae (Acari: Argasidae) and of temperature and photoperiod on induction and duration of diapause. Oecologia 1998, 113, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. Sociobiology; Harvard University Press: Cambridge, MA, USA, 1975. [Google Scholar]
- Lourenço, W.R. Scorpions and life-history strategies: From evolutionary dynamics toward the scorpionism problem. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 19. [Google Scholar] [CrossRef]
- Farley, R.D. Developmental changes in the embryo, pronymph, and first molt of the scorpion Centruroides vittatus (scorpiones: Buthidae). J. Morphol. 2005, 265, 1–27. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Li, S.; Lu, S.; Wang, X.; Cao, Z.; Wu, Y. Special Survival Strategy of First-Instar Scorpions Revealed by Synchronous Molting Behavior from Social Facilitation of Maternal Care and Reciprocal Aggregation. Insects 2024, 15, 726. https://doi.org/10.3390/insects15090726
Guo Y, Li S, Lu S, Wang X, Cao Z, Wu Y. Special Survival Strategy of First-Instar Scorpions Revealed by Synchronous Molting Behavior from Social Facilitation of Maternal Care and Reciprocal Aggregation. Insects. 2024; 15(9):726. https://doi.org/10.3390/insects15090726
Chicago/Turabian StyleGuo, Yiyuan, Songryong Li, Sijia Lu, Xinrong Wang, Zhijian Cao, and Yingliang Wu. 2024. "Special Survival Strategy of First-Instar Scorpions Revealed by Synchronous Molting Behavior from Social Facilitation of Maternal Care and Reciprocal Aggregation" Insects 15, no. 9: 726. https://doi.org/10.3390/insects15090726
APA StyleGuo, Y., Li, S., Lu, S., Wang, X., Cao, Z., & Wu, Y. (2024). Special Survival Strategy of First-Instar Scorpions Revealed by Synchronous Molting Behavior from Social Facilitation of Maternal Care and Reciprocal Aggregation. Insects, 15(9), 726. https://doi.org/10.3390/insects15090726