
Searching for More Effective Food Baits for Tephritid Fruit Flies (Diptera: Tephritidae): Performance of Newly Developed Vial-Lures Relative to Torula Yeast Borax


 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. General Methods
2.1.1. Traps
2.1.2. Lures
2.1.3. Trapping Protocol
2.2. Location-Specific Methods: Study Sites, Baits Tested, Target Species, and Experimental Design
2.3. Statistical Analysis
3. Results
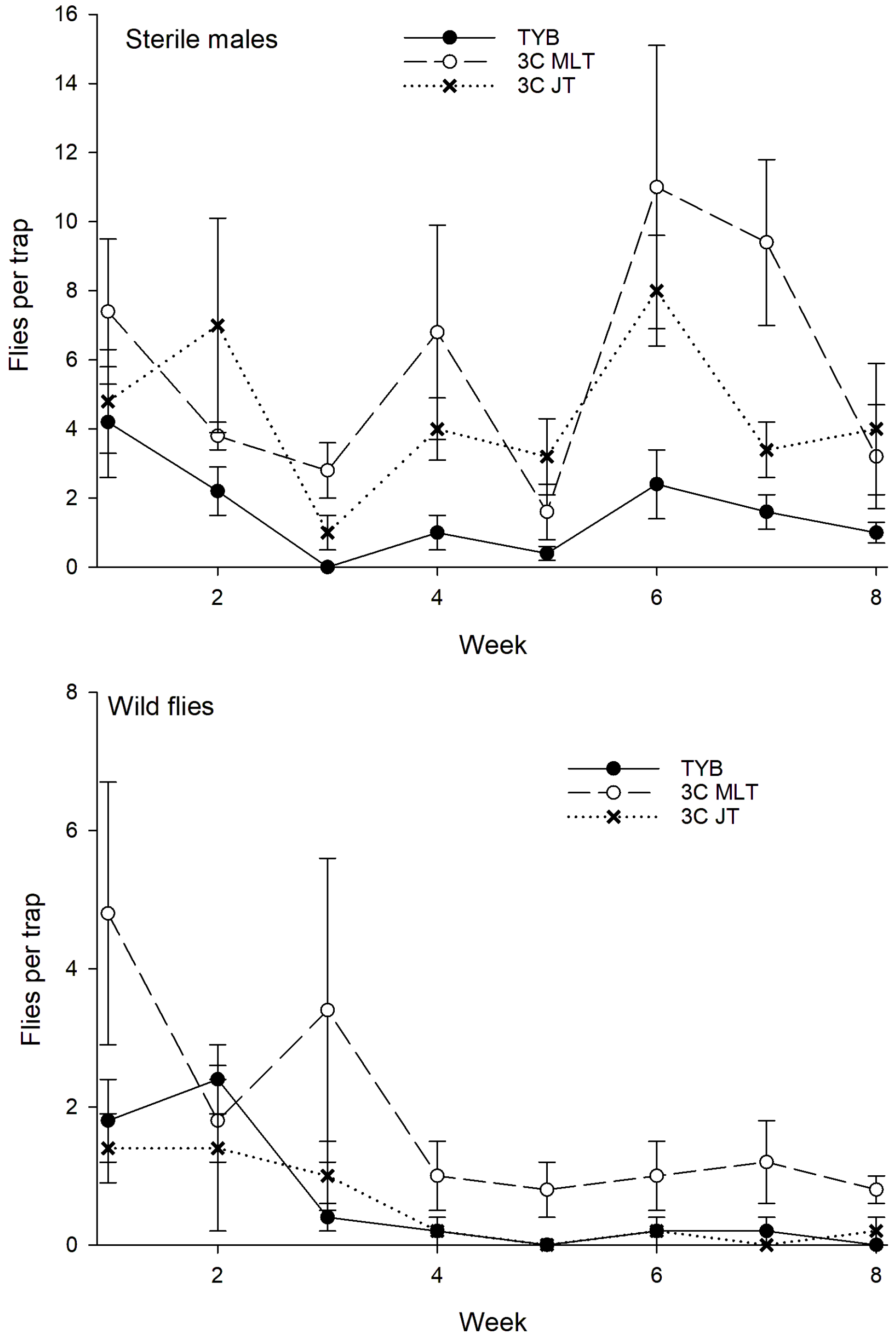
3.1. Ceratitis capitata
3.1.1. Argentina
3.1.2. Mexico
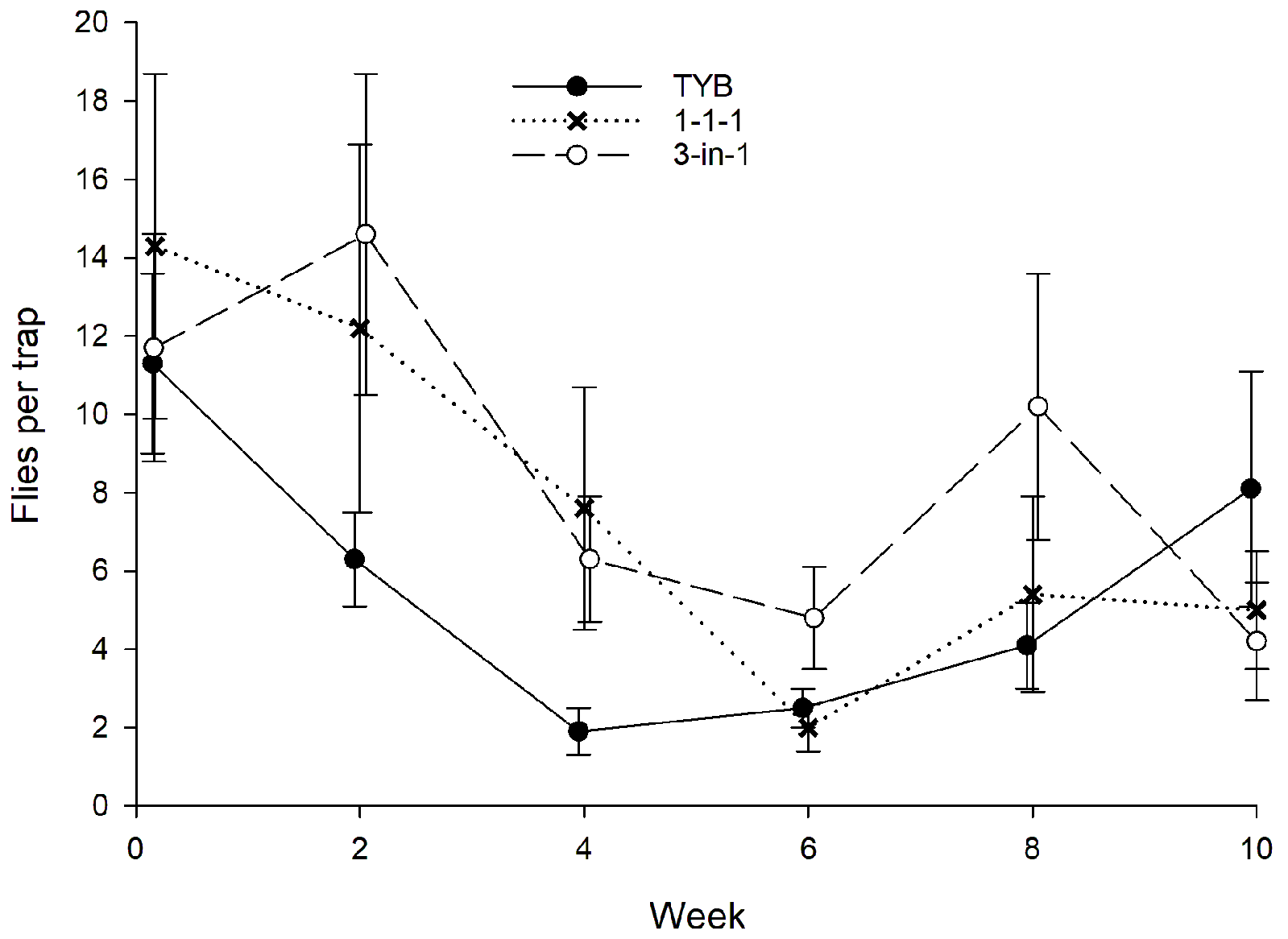
3.1.3. Hawaii
3.1.4. Colombia
3.2. Anastrepha obliqua
3.2.1. Mexico
3.2.2. Honduras
3.3. Anastrepha ludens
3.4. Anastrepha spp.
3.5. Bactrocera dorsalis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO/IAEA (Food and Agriculture Organization/International Atomic Energy Agency). Trapping Guidelines for Area-Wide Fruit Fly Programs, 2nd ed.; Enkerlin, W.R., Reyes-Flores, J., Eds.; FAO: Rome, Italy, 2018. [Google Scholar]
- Tan, K.H.; Nishida, R.; Jang, E.B.; Shelly, T.E. Pheromones, Male Lures, and Trapping of Tephritid Fruit Flies. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 15–74. [Google Scholar]
- Manrakhan, A.; Daneel, J.-H.; Beck, R.; Virgilio, M.; Meganck, K.; De Meyer, M. Efficacy of trapping systems for monitoring of Afrotropical fruit flies. J. Appl. Entomol. 2017, 141, 825–840. [Google Scholar] [CrossRef]
- Martinez, A.J.; Salinas, E.J.; Rendon, P. Capture of Anastrepha species (Diptera: Tephritidae) with Multilure traps and Biolure attractants in Guatemala. Fla. Entomol. 2007, 90, 258–263. [Google Scholar] [CrossRef]
- Shelly, T.E.; Kurashima, R.; Martin, R.M.; Fezza, T.; Bazelet, C. Weathering of 3-component synthetic food cones: Effects on residual amount, release rate, and field capture of 3 pest species of fruit flies (Diptera: Tephritidae). Environ. Entomol. 2023, 52, 408–415. [Google Scholar] [CrossRef]
- Royer, J.E.; Agovaua, S.; Bokosou, J.; Kurika, K.; Mararuai, A.; Mayer, D.G.; Niangu, B. Responses of fruit flies (Diptera: Tephritidae) to new attractants in Papua New Guinea. Austral Entomol. 2017, 57, 40–49. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Kouloussis, N.A.; Hendrichs, J.; Carey, J.R.; Heath, R.R. Early detection and population monitoring of Ceratitis capitata (Diptera: Tephritidae) in a mixed-fruit orchard in northern Greece. J. Econ. Entomol. 2001, 94, 971–978. [Google Scholar] [CrossRef]
- Epsky, N.D.; Kendra, P.E.; Schnell, E.Q. History and Development of Food-Based Attractants. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 75–118. [Google Scholar]
- Bateman, M.A.; Morton, T.C. The importance of ammonia in proteinaceous attractants for fruit flies (Family: Tephritidae). Aust. J. Agric. Res. 1981, 32, 883–903. [Google Scholar] [CrossRef]
- Mazor, M.; Peysakhis, A.; Galun, R. The role of ammonia in the attraction of females of the Mediterranean fruit fly to protein hydrolysate baits. Entomol. Exp. Appl. 1987, 43, 25–29. [Google Scholar] [CrossRef]
- Heath, R.R.; Epsky, N.D.; Guzman, A.; Dueben, B.D.; Manukian, A.; Meyer, W.L. Development of a dry plastic insect trap with food-based synthetic attractant for the Mediterranean and Mexican fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1995, 88, 1307–1315. [Google Scholar] [CrossRef]
- Calkins, C.O.; Schroeder, W.J.; Chambers, D.L. Probability of detecting Caribbean fruit fly, Anastrepha suspensa (Loew) (Diptera: Tephritidae), populations with McPhail traps. J. Econ. Entomol. 1984, 77, 198–201. [Google Scholar] [CrossRef]
- Torres-Quezada, E.A.; Mancebo, M.J.; Robles, L.A.; Maurer, W.; Cuesta, J.G.; Carreño, S.M.M.; Lopez, L. Optimizing torula bait for Anastrepha suspensa (Diptera: Tephritidae) trapping in the Dominican Republic. Fla. Entomol. 2021, 104, 18–26. [Google Scholar] [CrossRef]
- Miranda, M.A.; Alonso, R.; Alemany, A. Field evaluation of medfly (Dipt., Tephritidae) female attractants in a Mediterranean agrosystem (Balearic Islands, Spain). J. Appl. Entomol. 2001, 125, 333–339. [Google Scholar] [CrossRef]
- Broughton, S.; de Lima, C.P.F. Field evaluation of female attractants for monitoring Ceratitis capitata (Diptera: Tephritidae) under a range of climatic conditions and population levels in Western Australia. J. Econ. Entomol. 2002, 95, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Mwatawala, M.W.; de Meyer, M.; Makundi, R.H.; Maerere, A.P. Biodiversity of fruit flies (Diptera, Tephritidae) in orchards in different agro-ecological zones of the Morogoro region, Tanzania. Fruits 2006, 61, 321–332. [Google Scholar] [CrossRef]
- Conway, H.E.; Forrester, T.O. Comparison of Mexican fruit fly (Diptera: Tephritidae) capture between McPhail traps with torula and Multilure traps with Biolures in South Texas. Fla. Entomol. 2007, 90, 579–580. [Google Scholar] [CrossRef]
- Epsky, N.D.; Hendrichs, J.; Katsoyannos, B.I.; Vásquez, L.A.; Ros, J.P.; Zümreoglu, A.; Pereira, R.; Bakri, A.; Seewooruthun, S.I.; Heath, R.R. Field evaluation of female-targeted trapping systems for Ceratitis capitata (Diptera: Tephritidae) in seven countries. J. Econ. Entomol. 1999, 92, 156–164. [Google Scholar] [CrossRef]
- Epsky, N.D.; Kendra, P.E.; Peña, J.E.; Heath, R.R. Comparison of synthetic food-based lures and liquid protein baits for capture of Anastrepha suspensa (Diptera: Tephritidae) adults. Fla. Entomol. 2011, 94, 180–185. [Google Scholar] [CrossRef]
- Katsoyannos, B.I.; Heath, R.R.; Papadopoulos, N.T.; Epsky, N.D.; Hendrichs, J. Field evaluation of Mediterranean fruit fly (Diptera: Tephritidae) female selective attractants for use in monitoring programs. J. Econ. Entomol. 1999, 92, 583–589. [Google Scholar] [CrossRef]
- Sobrinho, R.B.; Lindemberg, A.; Mesquita, M.; Enkerlin, W.; Guimaraes, J.A.; Bandeira, C.T.; Peixoto, M.J.A. Evaluation of fruit fly attractants in the state of Ceará—Brazil. Rev. Cien. Agro. 2004, 35, 253–258. [Google Scholar]
- Thomas, D.B.; Epsky, N.D.; Serra, C.A.; Hall, D.G.; Kendra, P.E.; Heath, R.R. Ammonia formulations and capture of Anastrepha fruit flies (Diptera: Tephritidae). J. Entomol. Sci. 2008, 43, 76–85. [Google Scholar] [CrossRef]
- Ekesi, S.; Mohamed, S.; Tanga, C.M. Comparison of food-based attractants for Bactrocera invadens (Diptera: Tephritidae) and evaluation of mazoferm-spinosad bait spray for field suppression in mango. J. Econ. Entomol. 2014, 107, 299–309. [Google Scholar] [CrossRef]
- Thomas, D.B.; Holler, T.C.; Heath, R.R.; Salinas, E.J.; Moses, A.L. Trap-lure combinations for surveillance of Anastrepha fruit flies (Diptera: Tephritidae) in an agricultural area. Fla. Entomol. 2001, 84, 344–351. [Google Scholar] [CrossRef]
- Jang, E.B.; Holler, T.C.; Moses, A.L.; Salvato, M.H.; Fraser, S. Evaluation of a single-matrix food attractant tephritid fruit fly bait dispenser for use in federal detection programs. Proc. Hawaii. Entomol. Soc. 2007, 39, 1–8. [Google Scholar]
- Holler, T.C.; Peebles, M.; Young, A.; Whiteman, L.; Olson, S.; Sivinski, J. Efficacy of the Suterra Biolure individual female fruit fly attractant packages vs. the Unipak version. Fla. Entomol. 2009, 92, 667–669. [Google Scholar] [CrossRef]
- Shelly, T.; Kurashima, R.; Fezza, T. Field evaluation of three-component solid food-based dispenser versus torula yeast for capturing Mediterranean and oriental fruit flies (Diptera: Tephritidae). J. Asia-Pac. Entomol. 2020, 23, 825–831. [Google Scholar] [CrossRef]
- Shelly, T.; Fezza, T.; Kurashima, R. Captures of oriental fruit flies and melon flies (Diptera: Tephritidae) in traps baited with torula yeast borax solution or 2- or 3-component synthetic food cones in Hawaii. Fla. Entomol. 2022, 105, 37–43. [Google Scholar] [CrossRef]
- Holler, T.C.; Sivinski, J.; Jenkins, C.; Fraser, S. A comparison of yeast hydrolysate and synthetic food attractants for capture of Anastrepha suspensa (Diptera: Tephritidae). Fla. Entomol. 2002, 89, 419–420. [Google Scholar] [CrossRef]
- IAEA [International Atomic Energy Agency]. Development of Female Medfly Attractant Systems for Trapping and Sterility Assessment. In Final Report of a Coordinated Research Programme, 1995–1998, IAEA-TECDOC-1099; International Atomic Energy Agency Vienna International Centre: Vienna, Austria, 1998. [Google Scholar]
- Vargas, R.I.; Stark, J.D.; Nishida, T. Population dynamics, habitat preference, and seasonal distribution patterns of oriental fruit fly and melon fly (Diptera: Tephritidae) in an agricultural area. Environ. Entomol. 1990, 19, 1820–1828. [Google Scholar] [CrossRef]
- Leblanc, L.; Vargas, R.I.; Rubinoff, D. Captures of pest fruit flies (Diptera: Tephritidae) and nontarget insects in BioLure and torula yeast traps in Hawaii. Environ. Entomol. 2010, 39, 1626–1630. [Google Scholar] [CrossRef]
- Shelly, T.; Fezza, T.; Kurashima, R. Weathering of torula yeast borax food bait and capture of oriental, Mediterranean, and melon fruit flies in Hawaii (Diptera: Tephritidae). Proc. Hawaii. Entomol. Soc. 2022, 54, 53–62. [Google Scholar]
- Heath, R.R.; Epsky, N.D.; Dueben, B.D.; Rizzo, J.; Jeronimo, F. Adding methyl-substituted ammonia derivatives to a food-based synthetic attractant on capture of the Mediterranean and Mexican fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1997, 90, 1584–1589. [Google Scholar] [CrossRef]
- Hall, D.G.; Burns, R.E.; Jenkins, C.C.; Hibbard, K.L.; Harris, D.L.; Sivinski, J.M.; Nigg, H.N. Field comparison of chemical attractants and traps for Caribbean fruit fly (Diptera: Tephritidae) in Florida citrus. J. Econ. Entomol. 2005, 98, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, A.; Cloonan, K.R.; Rohde, B.B.; Gill, M.A.; Mosser, L.K.; Crane, J.H.; Carrillo, D.; Kendra, P.E. Attraction and longevity of 2- and 3-component food cone lures for the Caribbean fruit fly, Anastrepha suspensa (Diptera: Tephritidae). J. Econ. Entomol. 2022, 115, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Yuval, B. Perimeter trapping strategy to reduce Mediterranean fruit fly (Diptera: Tephritidae) damage on different host species in Israel. J. Econ. Entomol. 2000, 93, 721–725. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ceratitis capitata | |||
| Location | Treatment | Lure | Trap |
| Argentina | 1 | TYB | MLT |
| 2 | 3-in-1 vial-lure | MLT | |
| 3 | 3-in-1 vial-lure | JT | |
| Mexico | 1 | TYB | MLT |
| 2 | 3-in-1 vial-lure | MLT | |
| 3 | 3-in-1 vial-lure | JT | |
| Hawaii | 1 | TYB | MLT |
| 2 | 3-in-1 vial-lure | MLT | |
| 3 | 1-1-1 vial-lure | MLT | |
| Colombia | 1 | TYB | MLT |
| 2 | 3-in-1 vial-lure | MLT | |
| 3 | 3-in-1 vial-lure | JT | |
| Anastrepha spp. | |||
| Location | Treatment | Lure | Trap |
| Mexico | 1 | TYB | MLT |
| 2 | 2-in-1 vial-lure | MLT | |
| 3 | 2-in-1 vial-lure | JT | |
| Colombia | 1 | TYB | MLT |
| 2 | 2-in-1 vial-lure | MLT | |
| 3 | 2-in-1 vial-lure | JT | |
| Honduras | 1 | TYB | MLT |
| 2 | 2-in-1 vial-lure | MLT | |
| 3 | 2-in-1 vial-lure | JT | |
| Bactrocera dorsalis | |||
| Location | Treatment | Lure | Trap |
| Hawaii | 1 | TYB | MLT |
| 2 | 3-in-1 vial-lure | MLT | |
| 3 | 1-1-1 vial-lure | MLT | |
| Ceratitis capitata | |||
| Performance Relative | |||
| Location | Vial-Lure | Trap | to TYB/MLT |
| Argentina | 3-in-1 | MLT | + |
| 3-in-1 | JT | − | |
| Mexico | 3-in-1 | MLT | + |
| 3-in-1 | JT | ≈ | |
| Hawaii | 3-in-1 | MLT | ≈ |
| 1-1-1 | MLT | ≈ | |
| Colombia | 3-in-1 | MLT | ≈ |
| 3-in-1 | JT | − | |
| Anastrepha | |||
| Performance relative | |||
| Location | Vial-lure | Trap | to TYB/MLT |
| Colombia | 2-in-1 | MLT | − |
| Anastrepha spp. | 2-in-1 | JT | − |
| Honduras | |||
| A. obliqua | |||
| females | 2-in-1 | MLT | − |
| 2-in-1 | JT | − | |
| males | 2-in-1 | MLT | ≈ |
| 2-in-1 | JT | − | |
| Bactrocera dorsalis | |||
| Performance relative | |||
| Location | Vial-lure | Trap | to TYB/MLT |
| Hawaii | 3-in-1 | MLT | − |
| 1-1-1 | MLT | − | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enkerlin, W.; Arevalo, E.; Caballero, J.E.; Fezza, T.; Garavelli, E.; Martinez, D.B.; Rodriguez, P.A.; Shelly, T.; Thomas, M.E.; Villaseñor, A.; et al. Searching for More Effective Food Baits for Tephritid Fruit Flies (Diptera: Tephritidae): Performance of Newly Developed Vial-Lures Relative to Torula Yeast Borax. Insects 2025, 16, 53. https://doi.org/10.3390/insects16010053
Enkerlin W, Arevalo E, Caballero JE, Fezza T, Garavelli E, Martinez DB, Rodriguez PA, Shelly T, Thomas ME, Villaseñor A, et al. Searching for More Effective Food Baits for Tephritid Fruit Flies (Diptera: Tephritidae): Performance of Newly Developed Vial-Lures Relative to Torula Yeast Borax. Insects. 2025; 16(1):53. https://doi.org/10.3390/insects16010053
Chicago/Turabian StyleEnkerlin, Walther, Emilio Arevalo, Jose Eduardo Caballero, Thomas Fezza, Esteban Garavelli, Diana Beatriz Martinez, Pedro Alexander Rodriguez, Todd Shelly, Milthon Edgardo Thomas, Antonio Villaseñor, and et al. 2025. "Searching for More Effective Food Baits for Tephritid Fruit Flies (Diptera: Tephritidae): Performance of Newly Developed Vial-Lures Relative to Torula Yeast Borax" Insects 16, no. 1: 53. https://doi.org/10.3390/insects16010053
APA StyleEnkerlin, W., Arevalo, E., Caballero, J. E., Fezza, T., Garavelli, E., Martinez, D. B., Rodriguez, P. A., Shelly, T., Thomas, M. E., Villaseñor, A., & Flores, S. (2025). Searching for More Effective Food Baits for Tephritid Fruit Flies (Diptera: Tephritidae): Performance of Newly Developed Vial-Lures Relative to Torula Yeast Borax. Insects, 16(1), 53. https://doi.org/10.3390/insects16010053