1. Introduction
White grubs are the larvae of those scarabaeid beetles which feed below ground on plants roots. Although white grubs belong to the family Scarabaeidae, not all scarabaeids are white grubs—other groups such as dung beetles are also included in the family. Wilson [
1] provided a global review of what is known about white grub pests of sugar cane and their natural enemies, which is still valid. Wilson [
1] points out that Scarabaeidae is divided into several subfamilies, and white grubs occur in at least three of these: Melolonthinae, Rutelinae and Dynastinae. Dynastinae white grubs differ from the others in that the adults also feed on the cane and can cause very significant damage to young growth. The life cycle of a typical melolonthine white grub takes one year, although some species are known to take longer [
1].
The literature on sugar cane white grubs in Africa is limited, but it shows that the species of white grubs which cause damage to sugar cane vary from country to country. Box [
2] lists
Schizonycha vastatrix Chiaramonte (Melolonthinae) and
Heteronychus paolii Arrow (Dynastinae) as sugar cane pests in Somalia.
Heteronychus paolii is a replacement name for
H. sacchari Paoli which was described from Somalia, the adults attacking sugar cane below the surface of the ground [
3,
4]; it may well be a synonym of
H. abyssinica [
5]. There seems to be no other information available on the white grub pests of sugar cane in Somalia.
Le Pelley [
6] lists five Dynastinae, four Melolonthinae and three Rutelinae from Kenya, Uganda and Tanzania, which feed upon sugar cane. In Tanzania,
Cochliotis melolonthoides (Gerstaecker) was studied in some detail by Jepson [
7]; more recent studies and reports include those of Bjorking and Spry [
8], Waiyaki [
9,
10], Luhanga [
11], Luhanga
et al. [
12], and Saidi
et al. [
13]. There is a detailed report by Sweeney [
14] on the sugar cane pest species in Swaziland, updated by Williams [
15]. The dynastine
Heteronychus licas Klug is the most important white grub pest of sugar cane in Swaziland [
14,
16], parts of South Africa [
17] and is a problem as far west as Nigeria [
18].
Jepson [
7] studied
C. melolonthoides for two years in Tanzania, at the Arusha Chini Estate. The adults swarm from the first showers of the main rains in early October to the end of November, although there may be a subsidiary swarm at the beginning of the minor rains in March. The eggs which take 15 days to hatch are common in December. First instar larvae feed on humus and organic matter in the soil often at depths of up to 91 cm (36 inches); the second and third instars move up to feed upon roots much nearer the surface. The larvae develop to the third instar by June and continue feeding until August when they descend to a depth of 46–91 cm (18–36 inches) to pupate. The pupa last about 14 weeks before the adults emerge en masse again in October. Flight was recorded as lasting only from 18:40 h to 19:05 h and the beetles sometimes swarm around nearby trees. The adults were not observed to feed, and Jepson suggests the flight is purely for mate location. Jepson used a sample unit of 91 × 61 × 30.5 cm deep (36 × 24 × 12 inches) incorporating a cane stool (a primary shoot and its surrounding tillers) and its root system; it is not clear whether the quadrat was aligned across or along the cane row. Based on his experience in Tanzania and Mauritius he suggests that at 1000–8000 larvae/acre a light infestation is evident, at 8000–20,000/acre a moderate infestation and at 20,000–100,000/acre a heavy infestation; he suggests an economic damage level of 2–2.5 larvae per stool (8000–10,000/acre).
Jepson [
7] also observed the natural enemies of
C. melolonthoides in Tanzania.
Campsomeris mansueta (Gerstaecker), a large scoliid wasp, parasitized third instar larvae and may have accounted for 20%–50% mortality of that stage. Smaller species of
Campsomeris,
C. felina (Saussure),
C. lachesis (Saussure) and
C. caelebs (Sichel), attack second instar larvae but account for only 10%–25% mortality of this stage between them. A nematode was found in 2 out of 500 larvae, and considered of no significance at this level. Predatory beetle larvae, and asilid fly larvae were also present and probably capable of killing first instar larvae, but ants were not discussed. The incidence of predation, particularly of a soil living insect, is difficult to assess, but based on his observations, Jepson suggests predation might account for about 50% total larvae mortality.
Subsequently, Hocking [
19] reported that a fungus,
Ophiocordyceps barnesii (=
Cordyceps barnesii), killed mature larvae of
Cochliotis melolonthoides at Arusha Chini. Sung
et al. [
20] examined the phylogeny of
Cordyceps and transferred this and many similar fungi to the genus
Ophiocordyceps. In studies subsequent to those of Hocking, nearby in north-east Tanzania (Moshi), the only natural enemy Waiyaki [
9,
10] found was a “
Cordyceps sp” which was widespread in the estate, especially along the moist irrigation canals. This would also be
O. barnesii, which is further documented by Evans
et al. [
21]. However,
O. barnesii was originally described from Sri Lanka and Evans [
22] suggests that the African population is unlikely to be conspecific. The fungus infects only third instar larvae and the mummified remains from which the stromata develop are typically found in the top 3 cm of soil [
21].
At the Juba Sugar Project (JSP), Somalia, a combination of factors contributed to crop loss in the 1980s including poor drainage, soil types, warthogs and wild pigs, goats, cows, weeds, termites, as well as white grubs, although when the JSP was established, stem borers were considered the main insect pests [
23,
24]. In 1984 small local pockets of white grub infestation started to be noticed. A collection of insects associated with white grub damage was sent to CABI for determination around 1984; it included adults of
Cochliotis melolonthoides,
Schizonycha sp. and
Triodontella sp. Larvae were also sent, but the association of larvae and adults in the case of
Triodontella sp. and
Schizonycha sp. was only tentative. Subsequently, surveys were made at JSP and the large numbers of white grub larvae obtained in some areas was cause for alarm. The highest incidence of damage by white grubs was found in localized patches around the edge and in centers of some fields (
Figure 1). These surveys were carried out using quadrat samples of 0.5 × 0.5 × 0.5 m placed in the cane row (see
Section 2.2).
Figure 1.
Damage to sugar cane associated with white grubs, Juba Sugar Project, Somalia, October 1986. Note rough grassland in the middle distance, and “bush” in the distance.
Figure 1.
Damage to sugar cane associated with white grubs, Juba Sugar Project, Somalia, October 1986. Note rough grassland in the middle distance, and “bush” in the distance.
In May 1986, E. Tremblay and A.N. Jama spent two days at JSP. They collected with lights in the sugar cane and found at least four species of adult Scarabaeidae of which
Schizonycha sp. was dominant. In their short report, Tremblay and Jama [
25] suggest that since irrigation of the cane had been reduced since 1984, this may have caused the white grub problem by negating natural mortality factors such as pathogens. They went on to suggest improved irrigation might resolve the problem. It is against this background that MJWC spent 10 days at JSP at the end of October 1986 (the beginning of the minor rains) looking at the entire white grub complex [
26], and GBA spent two weeks at JSP in late April and early May 1987 (the beginning of the main rains) focusing on the two species of greatest concern,
C. melolonthoides and
Brachylepis werneri [
27].
2. Experimental Section
The studies reported here were carried out at the former Juba Sugar Project (JSP), south-western Somalia [
23]. The JSP was established and operated by Booker Agriculture International (now part of Booker Tate) to grow sugar for local consumption and export, but was closed down by 1991 with the outbreak of civil war in Somalia. It was located on the flood plain west bank of the Juba River, just south of the town of Jilib, from approximately 0.30N to 0.51N and 42.68E to 42.77E (Annex Figure 3.14 in [
28]). The JSP was destroyed in Somalia’s civil war and the land reverted to pastoralists [
29]. Examination of the JSP site using Google maps (2012) indicates that most of the area is now grazed, some is cultivated and some is inhabited. Because of these events over the last two decades, it has not been possible to assess changes in the white grub problem, or whether white grubs cause crop losses in the limited arable crops now present in the area.
The JSP soils were categorized as a mixture of fine, medium and coarse alluvial soils, on which the sugar cane was grown under irrigation throughout the year, and an area of marine plain unsuitable for cultivation. At the time of our visits three varieties made up the bulk of the sugar cane planted: Co 997, NCo 376, N53/216. The main rains (locally known as the gu rains) occur from the end of March to mid-July, and the minor rains (locally known as the deyr or der rains) from the end of September to early December, but rainfall is highly variable and periods of drought common.
At JSP, sugar cane was grown in blocks of about 10 hectares [
23], and each block was divided into about 12 fields. The northern half of the estate was termed Labadad South (LS) and comprised some 35 blocks, while the southern half was called Kamsuma North (KN) and covered 45 blocks. Abbreviations are used here (as they were at JSP) to designate particular fields, thus, KN06A/01 would be field 1 in block 6A of Kamsuma North
etc. The Management Compound (MC) was in the center of the estate, and some of the original “bush” (
Figure 1) was preserved within this area.
It should be pointed out that even at this time the infrastructure in Somalia was limited and many items taken for granted in most developing countries, e.g., plastic vials for medicines and kitchen containers that can be adapted for insect handling and rearing were simply not available. Hence, some of the materials used were based on what was available, e.g., oil drums and drink cans, rather than what would have been desirable.
2.1. The White Grub Species
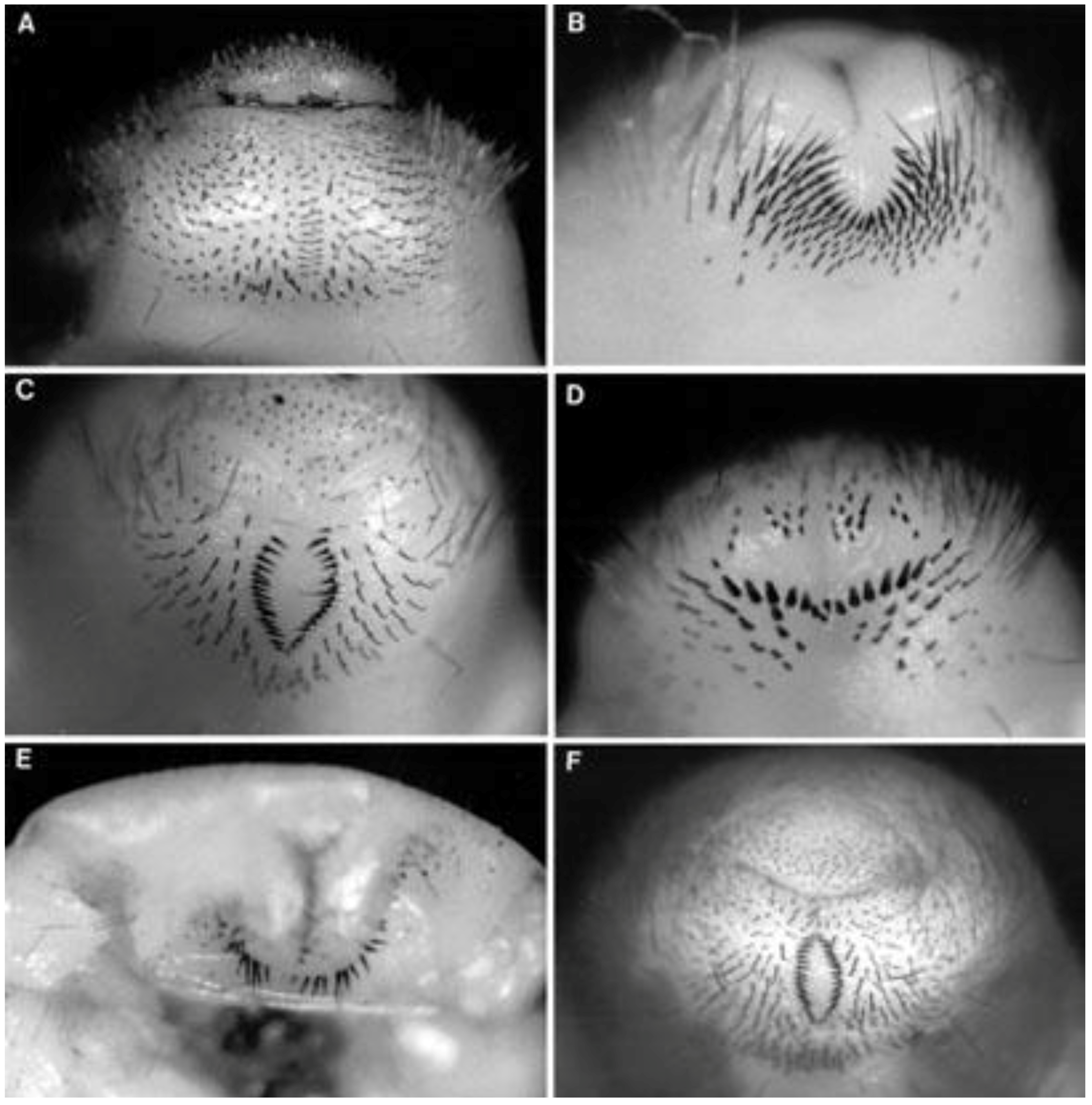
The taxonomy of beetles (like most insects) is based upon the adult stage. Hence, although it is the larvae of white grubs that cause most or all of the damage, the identification is mostly based on adults. Although larvae of different species are superficially similar, in Africa it has been found that many species can be distinguished by details of the raster (the arrangement of spines and hairs at the posterior end) which incorporates the pallidium (a compact arrangement of spines anterior to the anus), the shape of the anus, and characters of the mouthparts [
1,
14]. Thus, sampling the cane fields yielded at least seven recognizable morphospecies of white grub larvae based on examination of their rasters, while collecting at light produced rather more adults of genera known to have white grub larvae. Some of these were matched to adults during this study, others remain to be associated. As each morphospecies was recognized it was allocated a code letter: A, B, C,
etc. for larvae, and Z, Y, X, W,
etc. for adults. All morphospecies were checked by taxonomists of the CAB International Institute of Entomology at the Natural History Museum, London, the larvae by M.L. Cox and the adults by R. Madge. As a result, one additional uncommon ?
Schizonycha sp. larval morphospecies was recognized (
Section 3.1.7), and several additional adult
Schizonycha spp. (
Section 3.1.16). In preparing this paper, the identities of the two largest species were reassessed based on recent taxonomic reviews (
Section 3.1).
At JSP, the known white grub pests are all Melolonthinae, but representatives of Rutelinae and Dynastinae are also common at light and could have larvae attacking sugar cane. For example,
Heteronychus abyssinica Jack was quite common at lights at JSP, but commoner in the Management Compound than in the cane; the congeneric
H. licas is a known pest of sugar cane and other crops [
30] and reported from sugar cane in Somalia [
4]. As all studies were on-going simultaneously, understanding of the species and the correlation between larvae and adults improved over time. Each morphospecies is briefly introduced here to establish the species of concern, although the information presented is derived from the studies and observations that follow. Voucher specimens of all recognized larvae and adults were placed in the Natural History Museum, London.
There are three instars in the development of all white grubs [
1]. During each instar the head capsule, which is chitinized and rigid, remains the same size, while the body grows as the initially wrinkled skin is stretched to its maximum expansion. Hence, recognition of the different larval instars was based on head capsule size. After the October 1986 visit, MJWC examined preserved larvae, and measured the width of each head capsule. Based on this measurement, observations made with a hand lens in the field were confirmed and larvae of each species could be divided into instars. Not all instars were present, and where only one or two instars were present the allocation of instar number was provisional (e.g., larvae of
Schizonycha spp., unidentified Melolonthinae sp. and unidentified Cetoniinae sp.).
If this white grub problem were studied today, DNA barcoding [
31] could have been used in combination with traditional taxonomy tools to rapidly match white grub larvae with adults, help clarify any cryptic species, and help identify all stages. Recent work in South Africa has begun to develop this approach [
32].
2.2. Distribution of White Grub Larvae at JSP
Samples were taken from various sugar cane blocks and fields before, during and after the visits of MJWC and GBA, under the direction of JSP and BAI staff. For each field, 20 standard JSP 0.5 × 0.5 × 0.5 m pits were dug. These were arranged along the four equally spaced lateral irrigation lines, five sites per line, paced out at 10, 80, 160, 240 and 310 m from the edge of the field adjacent to the hydrant line. The quadrat was placed in the cane row, four rows across from the lateral line. The objective was to obtain samples from different fields in the blocks affected by white grubs, to assess the spatial and temporal distribution.
Because the diversity of white grub species was not recognized prior to October 1986, all white grubs were pooled prior to this date. However, it seems likely that most of the white grubs counted belonged to the two large, priority species, C. melolonthoides in the north of JSP and B. werneri LaCroix in the south. For samples taken after October 1986, the two priority species were distinguished together with their three larval instars.
There were other problems with some of the early samples. Pits were of irregular dimension, up to 1 m on one side; a measuring stick was introduced to produce regular quadrats (including depth). Not all pits were being dug in the cane row; some were in the inter-row but this was corrected.
2.3. Distribution of White Grub Larvae in the Soil in Relation to the Cane Row
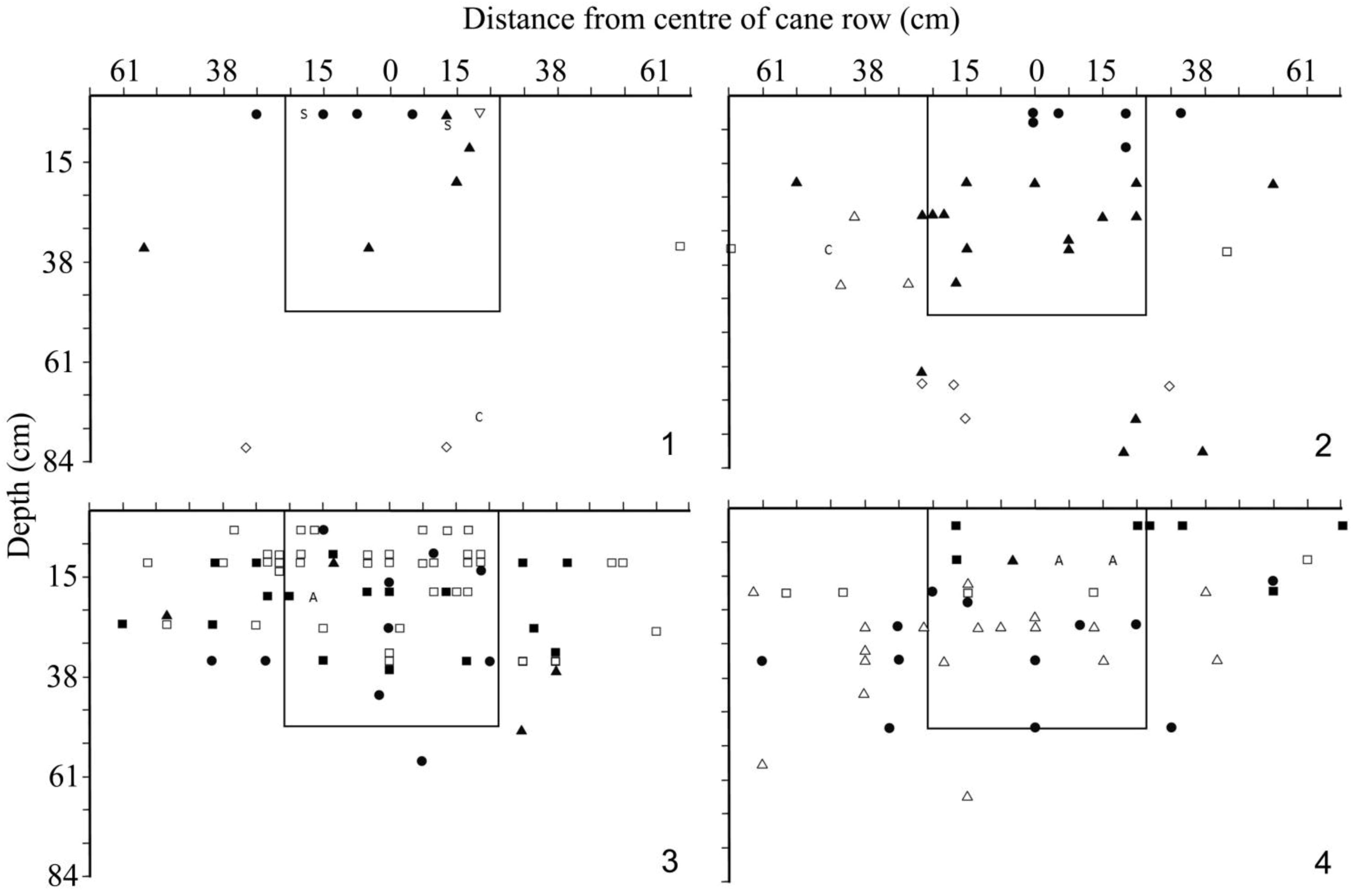
Jepson [
7] describes, and illustrates an example of, a method of making transects across the cane furrow to record the distance from the center of the furrow, and depth of all larvae found (in inches). This method was used to (1) obtain comparable data for the situation in Somalia, (2) to assess the effectiveness of the 0.5 × 0.5 × 0.5 m quadrat samples being used in the general survey at JSP (
Section 2.2), (3) to check for the presence of pupae of
C. melolonthoides, and (4) obtain material of early stages of all white grubs for evaluation and identification. Four similar transects were dug in JSP in December 1986: Two in KN06A and two in LS10/09.
A section of furrow of length 91 cm (36 inches) was selected so as to incorporate the center of a stool or group of stools, and the width of the transect to 61 cm (24 inches) on each side, marked out. A pit of 131 cm (51 inches) depth was then excavated in successive layers of 7.5 cm (3 inches). Within each layer, the distance was recorded at which each larva occurred from the center, and the larvae were categorized into instars and types (A, B, C, D etc.) based on the form of the raster. Although care was taken not to move the soil as it was searched, there was doubtless some movement of larvae during this process; however, because excavation was carried out in successive layers, the depth records should be accurate.
2.4. Sampling Adult Scarabaeidae Attracted to Light
A 45 gallon oil drum was converted to make a mercury vapor light trap similar to the Robinson trap [
33], but much deeper. It was operating with a 125 watt mercury vapor bulb and ballast. Trap localities were either in the JSP Management Compound where electricity was available throughout the night, or in the sugar cane blocks, where electricity was provided from a trailer mounted generator. The Management Compound contained stands of original riverine forest and was considered to represent the local natural or “bush” habitat. The trap was emptied by tipping the contents into a plastic box, which was sealed, labeled and placed in a freezer until the contents could be processed. Adult Scarabaeidae were identified to species or genus and counted.
Preliminary observations using the light trap in the JSP Management Compound supported earlier reports [
7] that Scarabaeidae beetles fly primarily in the early evening. Hence, for the first visit in December 1986, it was decided that the trap need be run only from dusk until 22:00 h. During three nights in April 1987, this was assessed more rigorously by emptying the trap at four intervals during the night, and recording the numbers of
C. melolonthoides caught for each time period separately. These numbers are not independent (beetles trapped in the first part of the night might otherwise have continued flying and been caught later) and so are not analysed further.
In October 1986, at the beginning of the minor rains, the light trap was run on four evenings in the cane fields and two nights in the Management Compound. On the second visit in April–May 1987, heavy rain at the belated onset of the main rains meant that access to the sugar cane by vehicle was very difficult, and the trap was mostly operated in the Management Compound. Between our two visits and for several nights after the second visit, the trap was run by JSP staff. The numbers of Scarabaeidae beetles captured were very variable and sometimes large, which meant that on some occasions, recording all white grubs captured was impractical, so counting focused on
C. melolonthoides and
B.
werneri, the two larger, individually more damaging species, which were assumed to be of greater economic importance because of their much lower damage threshold (
Section 3.9). Given the apparent changes in flight activity during the night and the diversity of trapping periods during the night (
Section 3.4), the results are not analysed or condensed further.
2.5. Reproductive Phenology of Cochliotis melolonthoides Based on Light Trap Samples
Adults of C. melolonthoides were present from the onset of the main rains in April and May, when large numbers were obtained from the light trap, but not in the minor rains in October. An analysis of the C. melolonthoides adults captured April–May 1987 was made. For either the entire sample, or taking a subsample when the number caught was large, each individual was sexed, and females were dissected to assess whether they were gravid or not, and on some occasions the number of mature eggs was counted.
2.6. Reproductive Phenology of Cochliotis melolonthoides and Brachylepis werneri Based on Field Samples
Irregular pit samples were dug in April and May 1987 to look for pathogens of white grubs. The number and stage of the two principal species were also recorded. Sample sites were selected under badly damaged cane and the fields sampled had previous records of high infestation. Soils were mainly sandy loams and bare patches were evident (
Figure 1).
2.7. Rearing Field Collected Larvae
At the beginning of this study, only one larva had been associated with its adult and named: C. melolonthoides. Hence, in parallel with other studies, an attempt was made to rear through field collected larvae of types B–F in order to link them to the corresponding adults, which could be identified. The following approach was used as most likely to produce results with minimal technical supervision between our visits.
Four 45 gallon oil drums were cut in half to give eight cylindrical half drums. Small holes were made in the base for drainage. Once the inside of the drum had been thoroughly cleaned, the base was covered inside with insect mesh and drums were filled with soil and a stool of sugar cane planted in each. In this process, soil and stools were taken from a field where no white grubs had been found in surveys, and the soil was first sifted by hand to check for the presence of white grubs. It was then necessary to cover the tubs with the cane stools planted so that the tub was ventilated and light, but no access or egress for Scarabaeidae adults was possible. Mosquito nets were cut up and made into cones; the base of the cone was then tied tightly around the top of the drum and the top tied to a prepared framework. The drums were placed against a south-east facing wall so that they received direct light for the first part of the morning only. The half-drums were set up on 1 November 1986 as shown in
Table 1.
Table 1.
Rearing trials studies set up in half-drums, 1 November 1986.
Table 1.
Rearing trials studies set up in half-drums, 1 November 1986.
| Drum | Larva species andsource | Number and stage |
|---|
| 1,2 | Brachylepis werneri (larva F) from LS10 | 10 instar III larvae |
| 3 | Unknown Melolonthinae sp. (larva D) from KN13 | 5 instar II and III larvae |
| 6 | ?Schizonycha sp. (larva C) from KN06A, KN13 and LS10 | 6 instar III larvae |
| 4,5 | Anomala ancilla (adult W) from light trap | 5 adult females with developed ovaries |
| 7 | ?Triodontella sp. A (larva B) from KN06A and KN13 | 26 instars II and III larvae |
| 8 | ?Triodontella sp. A (larva B) from KN06A and KN13 | 28 instars II and III larvae |
The larvae used for these field trials were taken from field samples, and although the larvae used appeared healthy, it is almost inevitable that, in spite of the care taken, the process of excavation and transport to the laboratory would have caused damage and trauma, leading to early death of some larvae.
The maintenance procedures anticipated included the following. The temperature in the drums was to be monitored and if high (>25 °C around cane roots) additional shading would be provided. The set up with these nets was somewhat insecure and if found to be adversely affected by high winds, a more secure arrangement would need to be devised. The drums were to be watered two times a week, by using a hose which can add water through the mesh tent, so that it need not be opened. At the same time a check should be made for any adult beetles. Any found should be removed (except for the adults already present in drums 4 and 5), preserved and labeled as to which drum they emerged from and when. Emergence of adults was considered most likely to be at the beginning of the next main rains in May. BAI/JSP staff excavated Drums 1, 4 and 8 to check on progress in November 1986, and the remainder was examined by GBA at the beginning of the main rains at the end of April.
Disappointing results were obtained using the half drums (see
Section 3.7), so a small scale rearing method was tested and used for bioassays. Field collected eggs of
C. melolonthoides were set up in individual containers (drink cans with the tops removed). The eggs were placed in soil and covered with moistened tissue. A rearing system for ?
Triodontella sp. A (larva B), unknown Melolonthinae sp. (larva D) and unknown Cetoniinae sp. (larva E) in similar plastic containers containing soil and grass were not successful, due to an abundance of nuisance ants,
Paratrechina sp. which adversely affected the larvae.
2.8. Alternative Hosts for Larvae
White grubs are usually considered polyphagous, at least amongst Poaceae. Local crops such as maize, millet and sorghum may be vulnerable, and wild grasses are likely hosts. In a clearing in LS10/11 (
Figure 1), three quadrats were dug amongst grasses and compared with quadrats from the adjacent cane row. The quadrats in the former area were selected by eye as representative, while those in the cane row were selected at random from within the line of the cane row.
2.9. Extrapolating Population Density—Damage Relationships
It was apparent from the samples taken by JSP staff, and the observations reported above that the distribution of white grubs within a field is highly contagious. Large differences can occur between apparently similar sites very close to each other, e.g., in
Section 3.3, pits 3 and 4 were excavated under almost identical stools yet one had more than twice as many white grubs as the other. Similarly, the population of white grubs may not be closely linked to the apparent damage, e.g., in
Section 3.3, pits 1 and 2 represent heavily damaged and relatively healthy stools a few yards apart, yet sample 2 actually has more large white grubs (instar 3
C. melolonthoides) than sample 1.
Clearly any attempt to link white grub populations with the damage caused will have to be based upon extensive studies. Any economic threshold value will be dependent upon the age of the crop, soil types and water stress as well as white grub species, stage and season. An alternative strategy is to extrapolate from published findings for the same or other white grubs attacking sugar cane elsewhere. Thus, Jepson's [
7] figure of 2–2.5 larvae per stool (or 8000–10,000/acre = 20,000–25,000/ha) for
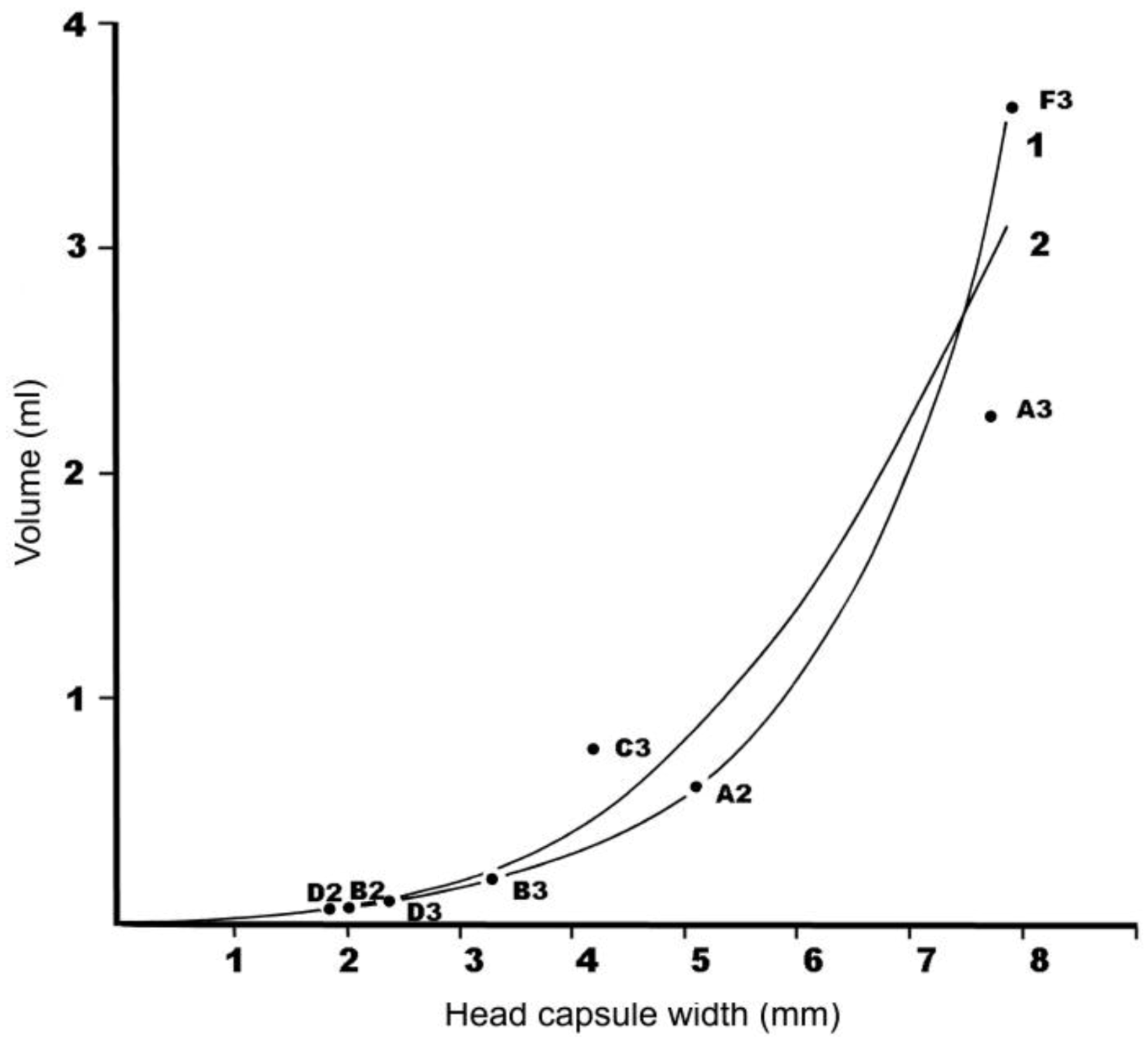
C. melolonthoides in Tanzania provides a standard. An approach was developed to estimate how this threshold might need to be modified to allow for the body size of the larvae of different white grub species. This was based on the assumptions that the amount of damage each larva does is a function of its body mass, and that body mass is a linear function of body volume. The former is a simplification of a complex interaction between white grubs and the sugar cane, but the later has been subsequently demonstrated, at least for Neotropical white grubs [
34].
During the October 1986 visit, field collected white grubs were preserved by immersing them in boiling water for 30 seconds for small larvae or up to two minutes for large larvae, and then placing them in 70% ethanol. A total of 70 larvae of six different species were thus preserved.
For each, the head capsule width was measured using a binocular microscope and a calibrated eye-piece graticule. The volume was measured by noting the change in level when a larva having been dried off with tissue paper was immersed in a partially filled measuring cylinder. In the case of the smallest larvae, they were measured in batches and the volume averaged. The bodies of the preserved larvae were more or less fully distended, due to the preservation method, so the volume is likely to be comparable to the final instar size. Larvae that were damaged or distorted were not included in the volume measurements.
The resultant volumes were plotted against head capsule width, and two models fitted to the relationship by linear least squares regression analysis (using the data analysis add-in in Excel): One based on a log-linear relationship, and the other on a cubed power. The equations were then used to calculate the economic threshold equivalence for third instar mature larvae of the different species. Since the results are compared with the results obtained in the same way for third instar C. melolonthoides, the approach should be valid.
2.10. Field Observations on Population Density—Damage Relationships
As noted in the introduction to
Section 2.9, the distribution of white grub larvae is very patchy, and the correlation between larval density and visible damage was not obvious in casual observations. To test this more objectively a scoring method for damage was prepared, so that larval densities in soil samples could be taken from stools different categories of damage.
In consultation with Mr. Peter Drew, BAI agronomist, a damage index was drawn up as follows:
1 = all tillers are healthy;
2 = up to 1/4 of tillers are yellow and stunted;
3 = 1/4 to all tillers are yellow and stunted;
4 = all of stool stunted, some dead or dying tillers; and
5 = stools either completely dead or only a few green tillers which are small and unhealthy.
In November 1986, three fields in block KN13 were selected by Mr. Drew, where all levels of damage index were present. Under his supervision, two sets of twenty standard pits of 0.5 × 0.5 × 0.5 m were subjectively located under stools in each damage category, and the number of instar 3 C. melolonthoides counted in each. The numbers were pooled for each set of 20 samples, giving 30 data points. Of the three fields sampled, KN13/08 was var. NS3-216 and the other two were C0997. The soils of these fields were fine grain with a very high percentage of silts, and the block was previously cut on 3 August 1986. The mean of each set of 20 quadrats was used to calculate a linear regression against damage index using the data analysis add-in in Excel.
2.11. Natural Enemies of White Grubs
The action of natural enemies can provide the foundation of an integrated pest management approach to pest control, while individual components of the natural enemy complex can be manipulated to augment their impact. Accordingly, evidence for the presence of natural enemies of white grubs was collected at JSP.
2.11.1. Field Observations
During field work at JSP, observations were made to detect predators, parasitoids or pathogens acting on white grubs. Common predators would have been observed in the course of other work; if these were predators of white grubs, predation events might be observed, such as ants actively feeding on larvae. Larval remains that may have been killed by predators would also have been found, especially when sieving the soil in quadrat sampling. To detect parasitoids, larvae were checked for evidence of ectoparasitism, the soil from the transect samples was sieved for pupating parasitoids, and samples of white grub larvae were dissected in the laboratory for evidence of internal parasitism. In addition, the dissection of adults to assess reproductive state (above) would have revealed well developed larvae of parasitoids of adults. Pathogens would have been detected as cadavers killed by bacteria or viruses, or mummified corpses with fungal fruiting bodies, which in the case of Ophiocordyceps spp. would be rather conspicuous.
2.11.2. Survey for Ophiocordyceps barnesii
In field observations and samples, Ophiocordyceps barnesii was found infecting third instar B. werneri in Labadad South only. Further observations were made by digging sample holes in the fields known to be infected with this pathogen (LS10/09 and LS10/08). Samples were not standardized, extending as appropriate to include the whole root system or until no material was being found, but the size of each hole was recorded. The fields sampled were bare and patchy in places, and densely overgrown with sorghum grass.
2.11.3. Sampling for entomopathogenic nematodes (EPNs)
Soil from block KN13/08 collected on 27 April 1987 was placed in individual drink cans and one first instar larva of C. melolonthoides was added to each container. At this time only first instar larvae were available, otherwise third instar larvae would have been used. After one week, on 4 May 1987, the larvae were checked for evidence of infection by EPNs, although if time had not been limiting this would have been done after ten days. In the one case where this was found, five third instar B. werneri larvae were added to the soil sample, and checked for evidence of EPNs after three days.
4. Discussion and Conclusions
During two brief visits to Somalia, we have accumulated information on the biology, phenology, pest incidence and management options for white grubs in south-west Somalia. Given the time limitations, much of this information is of a preliminary nature, but is sufficient to give clear indications as to which are the economically damaging species, their biology and phenology. Useful indications have been documented regarding sampling methods and economic thresholds. Biological control options have been identified but would need further development and testing. However, bearing in mind that these observations were made 25 years ago, some conclusions are still valid, but others, particularly those relating to scarabaeid incidence and damage, can only be considered snapshots from the past, and new studies would be needed to assess the current situation.
The results show that a large proportion of the larvae found in the transect would have been missed in a standard quadrat: 42, 55, 38 and 52% respectively for samples 1–4 (
Figure 5). If the quadrat size was changed to 1 × 0.5 × 0.5 m across the cane row the percentage missed would be reduced to 10, 21, 31 and 30 for the four holes. If, however only larvae of
C. melolonthoides and
B.
werneri are considered, the standard quadrat would have missed 30, 50, 25 and 17% while a 1m wide quadrat would only miss 10, 17, 0 and 0% respectively. However, it needs to be asked, how reliable are the records of distance from the center of the cane row? As suggested above, there was undoubtedly some displacement of the larvae during excavation, but overall this should have evened out, and in any event many larvae were discovered in situ including almost all large individuals (
C. melolonthoides and
B.
werneri). Hence, the recorded position of individual larvae may be in error but the pattern is probably reliable. This could be confirmed by excavating additional transects using Jepson's [
7] method in which an initial trench was dug in the inter row parallel to the cane row, and then extended 7.5 cm (3 inches) at a time across the row. In this way a reliable measure of the distance of each larva from the cane row would be obtained (at the cost of potential inaccuracies in the depth measurements). If, in future, absolute measures of population density are needed, it would be appropriate to use a quadrat at least 1 m wide across the cane row. However, the advantage of smaller quadrat size is clear for logistic reasons, and this is still appropriate, providing it is recognized that a proportion of the white grub larvae will be missed as they are further from the middle of the cane row than the quadrat extends. Further sampling would be needed to assess whether some consistent conversion factors could be derived and applied.
As Carnegie and Leslie [
37] concluded in South Africa, the use of a light trap provides a cost effective way of monitoring adult populations of those Scarabaeidae species which fly by night. Although there may be white grubs which do not fly by night, there is no evidence that any of these occur in JSP, since the same species dug from sugar cane fields as adults were also caught in the light trap. Using a light trap also provides a very efficient method of collecting specimens for identification and taxonomic studies. For example in KN06A, a day's digging of 2 m
3 produced 2 adult
Triodontella sp. A; the next night the light trap caught 1,023 at the same place.
Based on the results obtained, it seems that C. melolonthoides and perhaps B. werneri (the two larger species) have an annual life cycle with the adults flying at the beginning of the main rains only. In contrast, at least some of the smaller species fly in both rains, although we cannot say whether this represents overlapping generations or two generations per year linked to the bimodal rains.
The method adopted here to extrapolate population density – damage relationships is far from precise and includes several untested assumptions, but it provides an approach to assess the relative damage that different sized white grubs may do to sugar cane. The indication that white grubs of the smaller species (e.g., ?Triodontella sp. A (larva B) and unknown Melolonthinae sp. (larva D)) can be tolerated at densities 10–30 times those of the two large species (C. melolonthoides and B.werneri) may be a useful guide.
Comparing the larval densities recorded at JSP with these economic thresholds, the only species likely to locally exceed the economic threshold at the time these observations were made are C. melolonthoides, B. werneri and possibly Schizonycha spp. Although sometimes more common, the smaller larvae of ?Triodontella sp. A (larva B), unknown Melolonthinae sp. (larva D) and unknown Cetoniinae sp, (larva E) did not reach their estimated economic thresholds.
This has implications for the most efficient survey strategy. If C. melolonthoides and B. werneri have annual life cycles, and October (and perhaps a few months following) is the period when they are full grown, then survey work should concentrate on this period when the only species of economic importance are present as large larvae and hence immediately distinguishable from small unimportant species.
Although the sample size was comparatively small and should be repeated in other fields with different soil types, the correlation of white grub population density with the damage index showed promise as a way to simply and quickly assess white grub populations and damage without the major effort of comprehensive soil sampling, although spot sampling should still be anticipated to confirm which species is present. For C. melolonthoides soil sampling would be restricted to the season when third instar larvae are present, October–December.
Several natural enemies of
Cochliotis melolonthoides have been reported from Tanzania [
7], but no evidence was found for natural enemies attacking this species at JSP. For example, although
Ophiocordyceps barnesii has been reported as a sometimes common pathogen of
Cochliotis melolonthoides in Tanzania [
9,
20,
21], in our studies we found it affecting
B.
werneri only, even though
C. melolonthoides was considerably more common in nearby blocks. Cross infectivity trials could not be attempted in May 1987 as inadequate numbers of third instar
C. melolonthoides larvae were available. In the rearing trails in half drums, set up in October 1986 using soil from white grub-free blocks, only
B.
werneri became infected with
O. barnesii. However, this provides only circumstantial evidence as most larvae died anyway, there were no trials with
C. melolonthoides and the
B.
werneri larvae may well have been infected before they were set up.
The EPNs and O. barnesii found in our surveys are both considered to have potential as biopesticides, if interventions were needed to reduce populations of white grubs. EPNs can be easily mass produced and are widely tested and used for the control of pests in restricted, moist places, including soil pests. Ophiocordyceps fungi cannot be mass produced at this time, so the immediately available option would be to field-collect the sporing bodies and redistribute the spores to areas where white grub damage occurs, but the fungus does not seem to be common.
The white grub incidence within individual fields is very heterogeneous, and outbreaks had been very localized. We considered to what extent such a pattern can be linked to physical factors such as soil type, cane variety, levels of irrigation, presence of alternative hosts etc. We concluded that the data to answer this question are not available, and extensive and prolonged surveys would be necessary to find any correlations and manipulative experiments to then demonstrate cause and effect
At the present time, although Somalia depends heavily on agriculture to sustain its economy [
38], the large scale growing of sugar cane is not being practiced due to the on-going state of unrest. This is starting to change, so that in the future, and it may be that sugar cane production will be developed again, in which case, these observations will be relevant to that future production system. On the other hand, other priorities may arise, so that food crops or other agricultural land-use is developed in south-west Somalia. However, given that white grubs are polyphagous, then many of the food crops likely to be grown will also be affected, so again these observations will be relevant. Finally, the observations, as they relate to crops grown under irrigation in an area of weak, irregular, bimodal rainfall, may be relevant to other parts of Africa, either now, or in the future as a result of climate change. Thus, although not immediately applicable in Somalia, we believe these observations and conclusions will be useful in the future.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




