
Dynamic Responses in a Plant-Insect System to Fertilization by Cormorant Feces

Abstract
:1. Introduction
2. Study Organism
3. Results
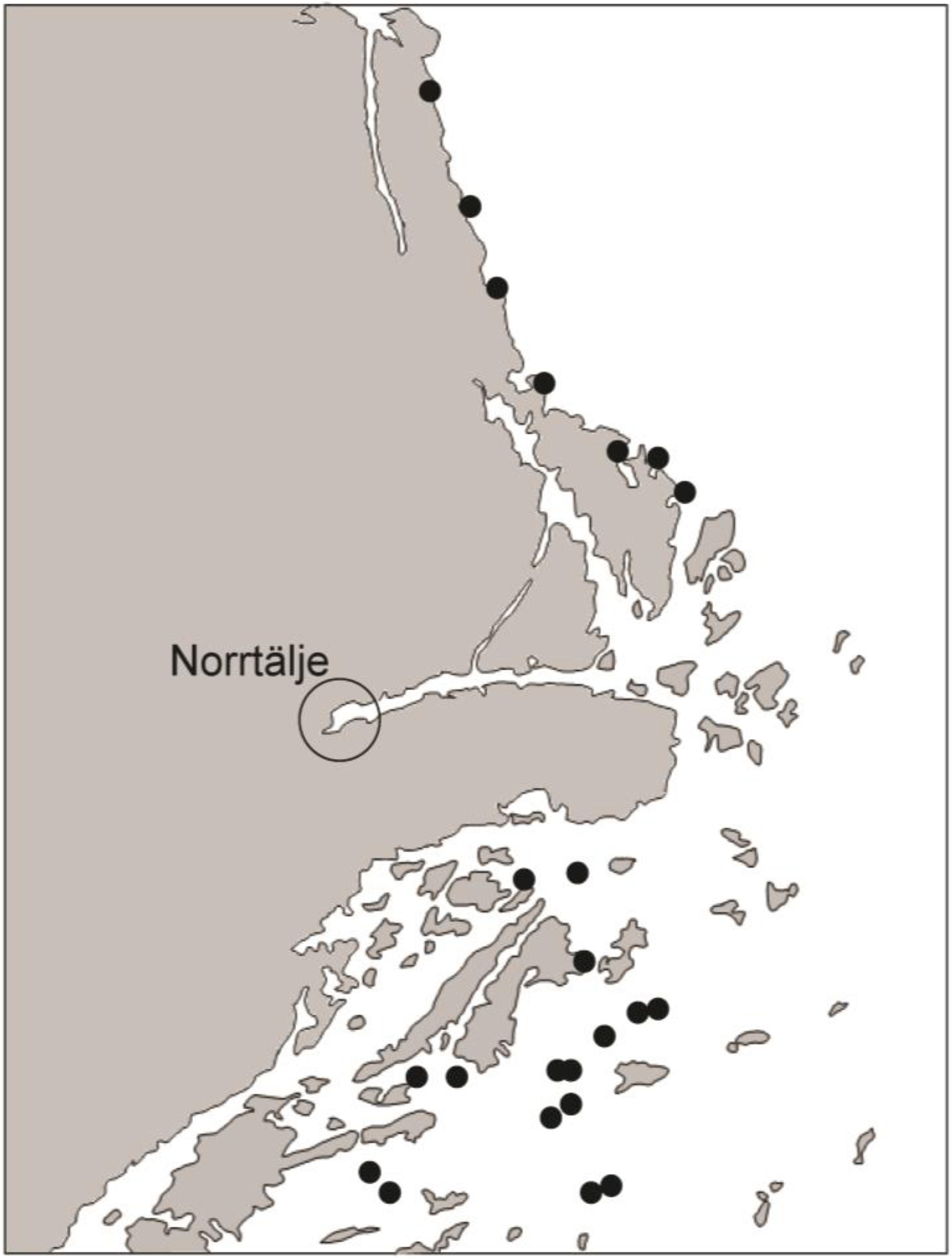
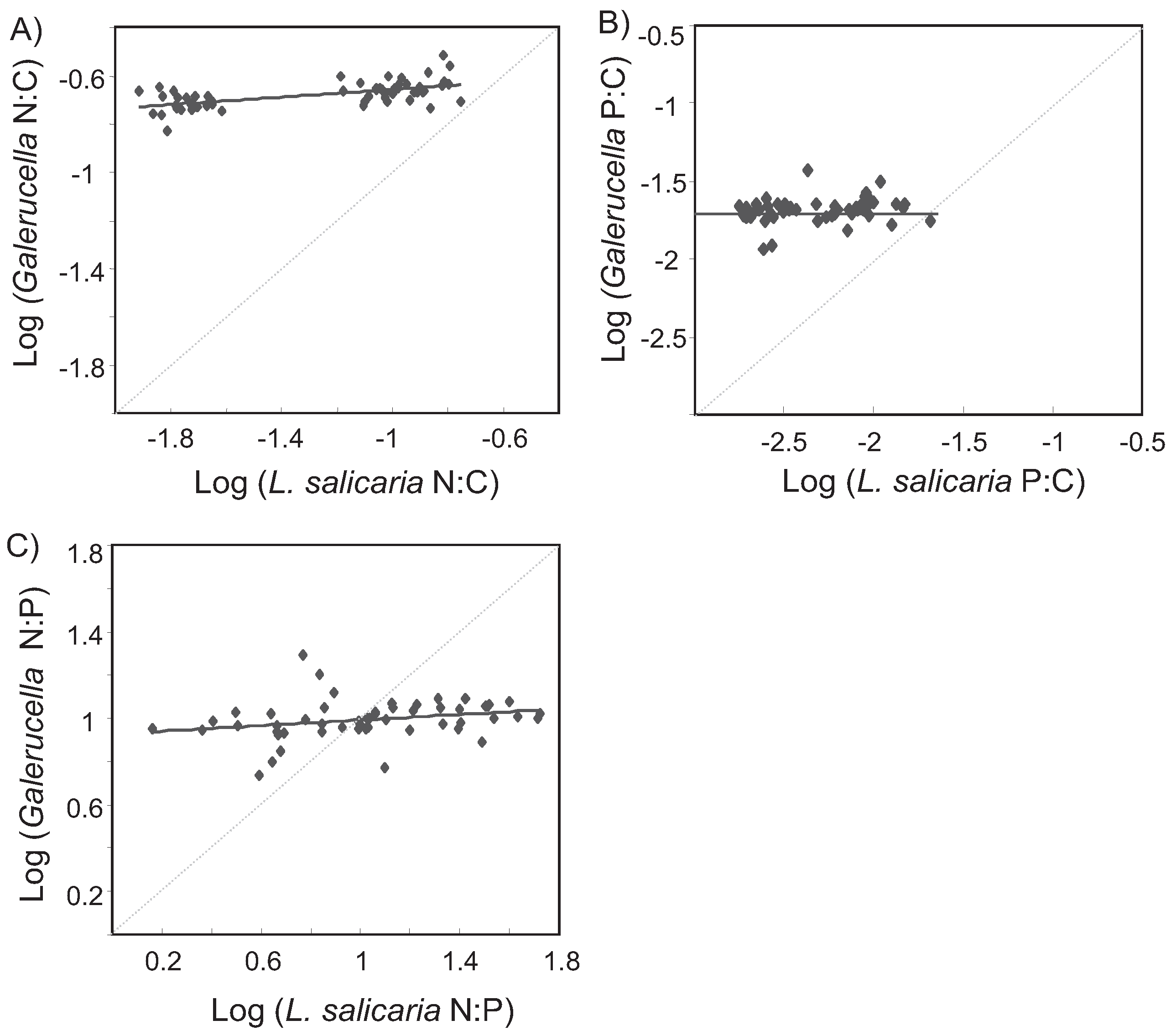
3.1. Field Study



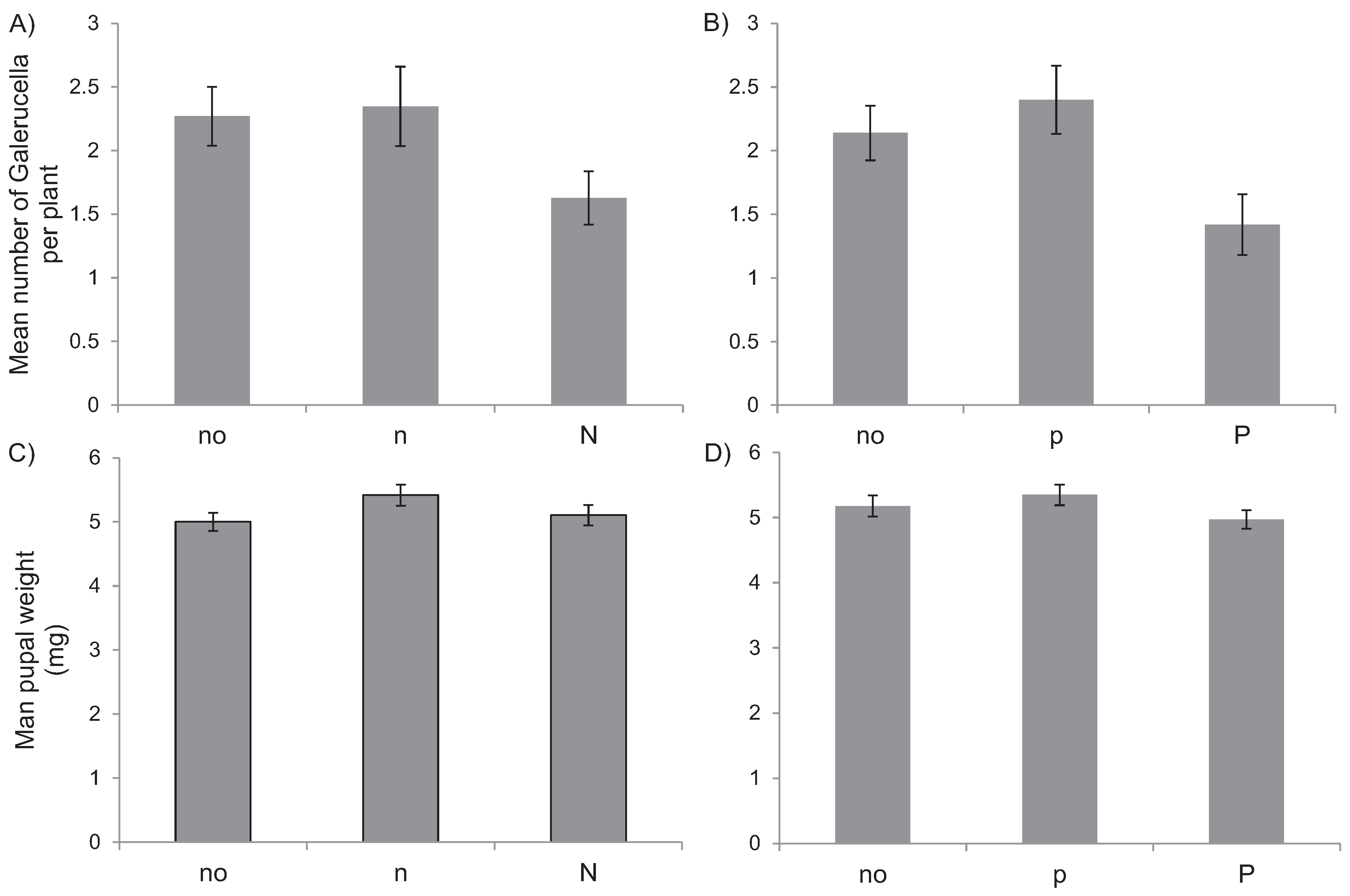
3.2. Pot Experiment



4. Discussion
5. Methods
5.1. Field Study
5.1.1. Soil and Plant Analyses
5.1.2. Statistics
5.2. Pot Experiment
Statistics
6. Conclusions
Acknowledgments
Author Contributions
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | np | nP | N | Np | NP | p | P | no | |
|---|---|---|---|---|---|---|---|---|---|
| Na2HPO4 (g/L) | 0 | 0.94 | 2.51 | 0 | 0.94 | 2.51 | 0.94 | 2.51 | 0 |
| H4N2O3 (g/L) | 3.34 | 3.34 | 3.34 | 8.91 | 8.91 | 8.91 | 0 | 0 | 0 |
Conflicts of Interest
References
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology; OUP: Oxford, UK, 2005. [Google Scholar]
- Gratton, C.; Denno, R.F. Seasonal shift from bottom-up to top-down impact in phytophagous insect populations. Oecologia 2003, 134, 487–495. [Google Scholar] [CrossRef]
- Haddad, N.M.; Haarstad, J.; Tilman, D. The effects of long-term nitrogen loading on grassland insect communities. Oecologia 2000, 124, 73–84. [Google Scholar] [CrossRef]
- White, T.C.R. The Inadequate Environment; Springer-Verlag: Berlin, Germany, 1993. [Google Scholar]
- Kolb, G.S.; Jerling, L.; Hambäck, P.A. The impact of cormorants on plant-arthropod food webs on their nesting islands. Ecosystems 2010, 13, 353–366. [Google Scholar] [CrossRef]
- Meyer, G.A.; Root, R.B. Influence of feeding guild on insect response to host plant fertilization. Ecol. Entomol. 1996, 21, 270–278. [Google Scholar] [CrossRef]
- McCauley, E.; Nisbet, R.M.; Murdoch, W.W.; de Roos, A.M.; Gurney, W.S.C. Large-amplitude cycles of Daphnia and its algal prey in enriched environments. Nature 1999, 402, 653–656. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Paradox of enrichment: destabilization of exploitation ecosystems in ecological time. Science 1971, 171, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Chattopadhyay, J. The stability of ecosystems: A brief overview of the paradox of enrichment. J. Biosci. 2007, 32, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Arditi, R.; Ginzburg, L.R. Coupling in predator-prey dynamics: Ratio-Dependence. J. Theor. Biol. 1989, 139, 311–326. [Google Scholar] [CrossRef]
- Gurney, W.S.C.; Nisbet, R.M. The regulation of inhomogeneous populations. J. Theor. Biol. 1975, 52, 441–457. [Google Scholar] [CrossRef] [PubMed]
- Hultén, E.; Fries, M. Atlas of North European vascular plants north of the tropic of cancer. I-III; Koeltz: Königstein, Germany, 1986. [Google Scholar]
- Hambäck, P.A.; Ågren, J.; Ericson, L. Associational resistance: Insect damage to purple loosestrife reduced in thickets of sweet gale. Ecology 2000, 81, 1784–1794. [Google Scholar] [CrossRef]
- Hight, S.D.; Blossey, B.; Laing, J.; Declerck-Floate, R. Establishment of insect biological control agents from Europe against Lythrum salicaria in North America. Environ. Entomol. 1995, 24, 967–977. [Google Scholar] [CrossRef]
- Hambäck, P.A. Density-dependent processes in leaf beetles feeding on purple loosestrife: Aggregative behaviour affecting individual growth rates. Bull. Entomol. Res. 2010, 100, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, J.A.; Hambäck, P.A.; Ericson, L. Herbivore-induced “rent rise” in the host plant may drive a diet breadth enlargement in the tenant. Ecology 2008, 89, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.R.; Faeth, S.H. The consequences of larval aggregation in the butterfly Chlosyne lacinia. Ecol. Entomol. 1997, 22, 408–415. [Google Scholar] [CrossRef]
- Wise, M.J.; Kieffer, D.L.; Abrahamson, W.G. Costs and benefits of gregarious feeding in the meadow spittlebug, Philaenus spumarius. Ecol. Entomol. 2006, 31, 548–555. [Google Scholar] [CrossRef]
- Kolb, G.; Palmborg, C.; Hambäck, P.A. Ecological stoichiometry and density responses of plant-arthropod communities on cormorant nesting islands. PLOS ONE 2013, 8, e61772. [Google Scholar] [CrossRef] [PubMed]
- Kagata, H.; Ohgushi, T. Nitrogen homeostasis in a willow leaf beetle, Plagiodera versicolora, is independent of host plant quality. Entomol. Exp. Appl. 2006, 118, 105–110. [Google Scholar] [CrossRef]
- Hambäck, P. Why purple loosestrife in sweet gale shrubs are less attacked by herbivorous beetles? (In Swedish with English abstract). Entomol. Tidskr. 2004, 125, 93–102. [Google Scholar]
- Hesse, P.R. A Textbook on Soil Chemical Analysis; Chemical Publishing Co., Inc.: New York, NY, USA, 1971. [Google Scholar]
- Application Note (AN 50/84). Determination of Ammonia Nitrogen by Flow Injection Analysis and Gas Diffusion; Foss Tecator AB: Höganäs, Sweden, 1984.
- Application Sub Note (ASN 50-01/92). Ammonia; Foss Tecator AB: Höganäs, Sweden, 1992.
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Analysis of Water and Wastewater; American Water Works Association; Water Pollution Control Federation: Washington, DC, USA, 1998. [Google Scholar]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolb, G.; Hambäck, P.A. Dynamic Responses in a Plant-Insect System to Fertilization by Cormorant Feces. Insects 2015, 6, 419-431. https://doi.org/10.3390/insects6020419
Kolb G, Hambäck PA. Dynamic Responses in a Plant-Insect System to Fertilization by Cormorant Feces. Insects. 2015; 6(2):419-431. https://doi.org/10.3390/insects6020419
Chicago/Turabian StyleKolb, Gundula, and Peter A. Hambäck. 2015. "Dynamic Responses in a Plant-Insect System to Fertilization by Cormorant Feces" Insects 6, no. 2: 419-431. https://doi.org/10.3390/insects6020419
APA StyleKolb, G., & Hambäck, P. A. (2015). Dynamic Responses in a Plant-Insect System to Fertilization by Cormorant Feces. Insects, 6(2), 419-431. https://doi.org/10.3390/insects6020419