
Diffusion Rates and Dispersal Patterns of Unfed versus Recently Fed Bed Bugs (Cimex lectularius L.)

Abstract
:1. Introduction
2. Methods
2.1. Bed Bugs, Arena, and Substrates


2.2. Protocol
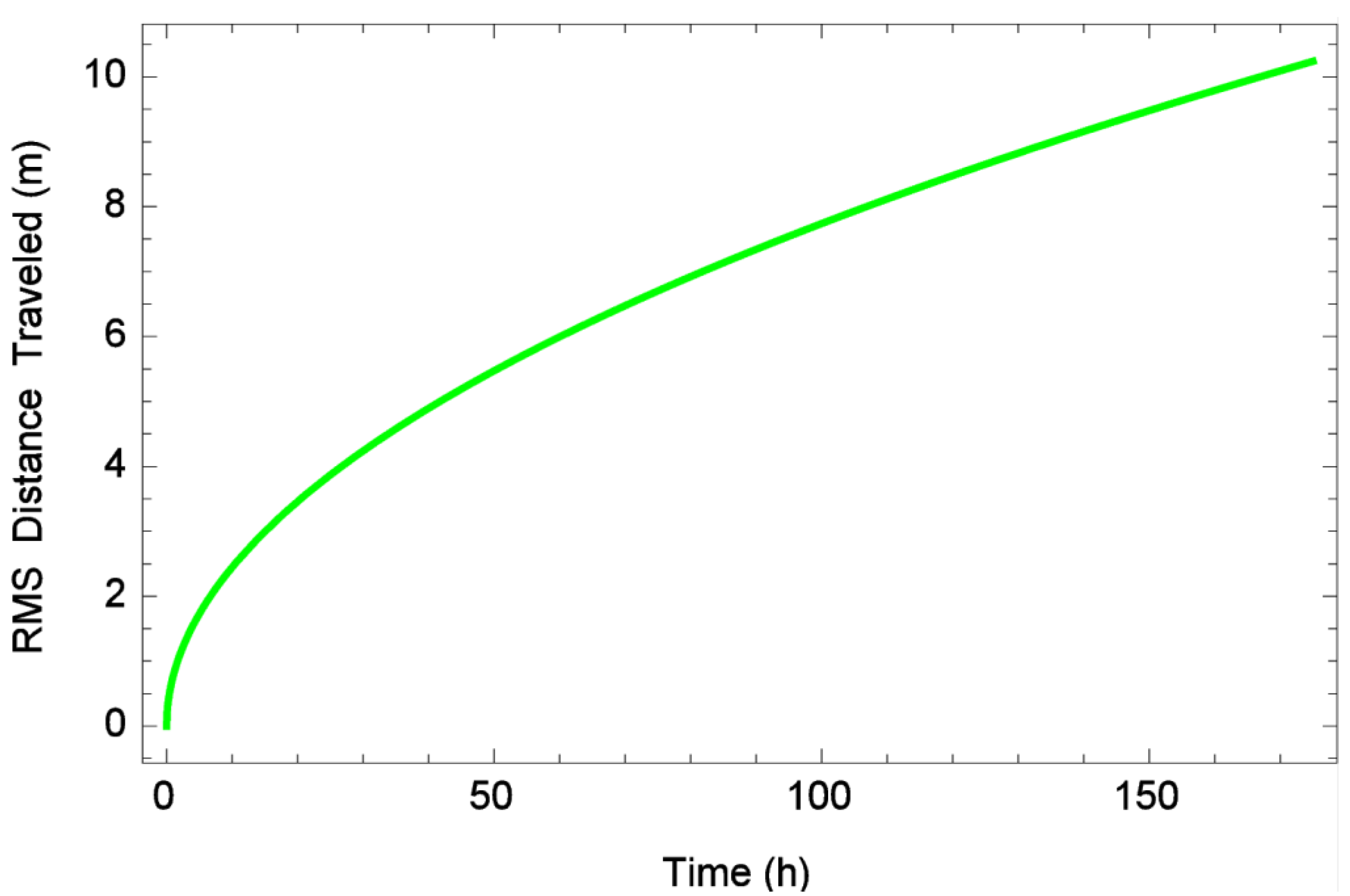
2.3. Calculation of Diffusion Rates
- (1)
- selecting a search range for estimates of the diffusion rate, and (for the initial range for all reps, to was chosen based on a priori observations that indicated this was the optimal range for possible diffusion rates)
- (2)
- creating a mesh of the interval with 100 estimates of the diffusion rate, i.e., , (with the initial case giving for and );
- (3)
- finding the smallest SS difference, say , and refining the search range to and ;
- (4)
- repeating steps (2)–(3) until .
2.4. Statistical Analyses
3. Results and Discussion

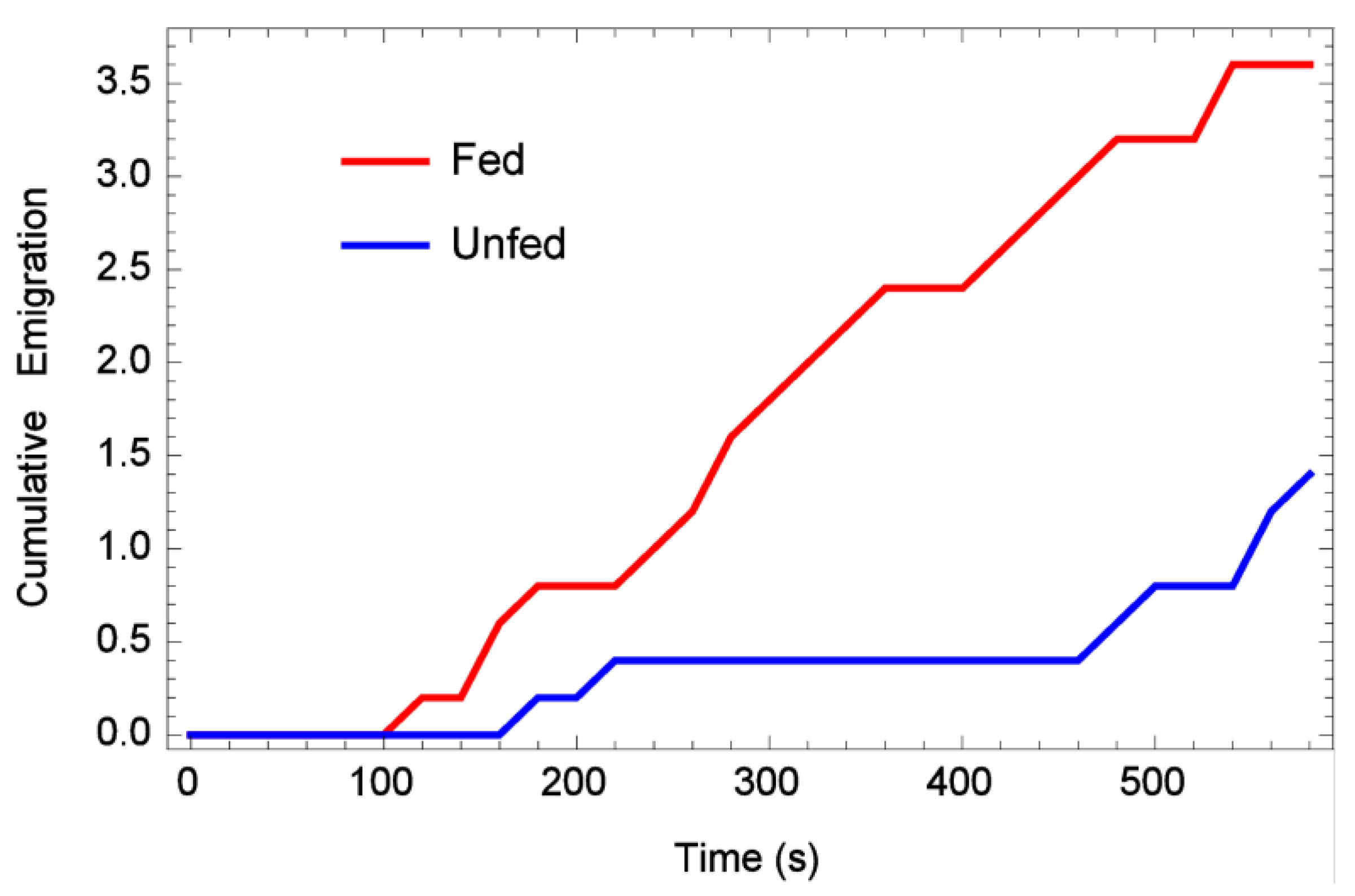
3.1. One-Dimensional Dispersal

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number that Moved Very Little (<4 cm) | ||
| ARENA TYPE | FED | UNFED |
| one-dimensional | 9♀ 16♂ (50%) | 14♀ 13♂ (54%) |
| two-dimensional | 5♀ 8♂ (26%) | 4♀ 8♂ (24%) |
| Number that moved as far as possible (22.5–25 cm one-dimensional or 34–41 cm two dimensional) * | ||
| ARENA TYPE | FED | UNFED |
| one-dimensional | 10♀ 5♂ (30%) | 7♀ 5♂ (24%) |
| two-dimensional | 6♀ 5♂ (22%) | 2♀ (4%) |

3.2. Two-Dimensional Dispersal


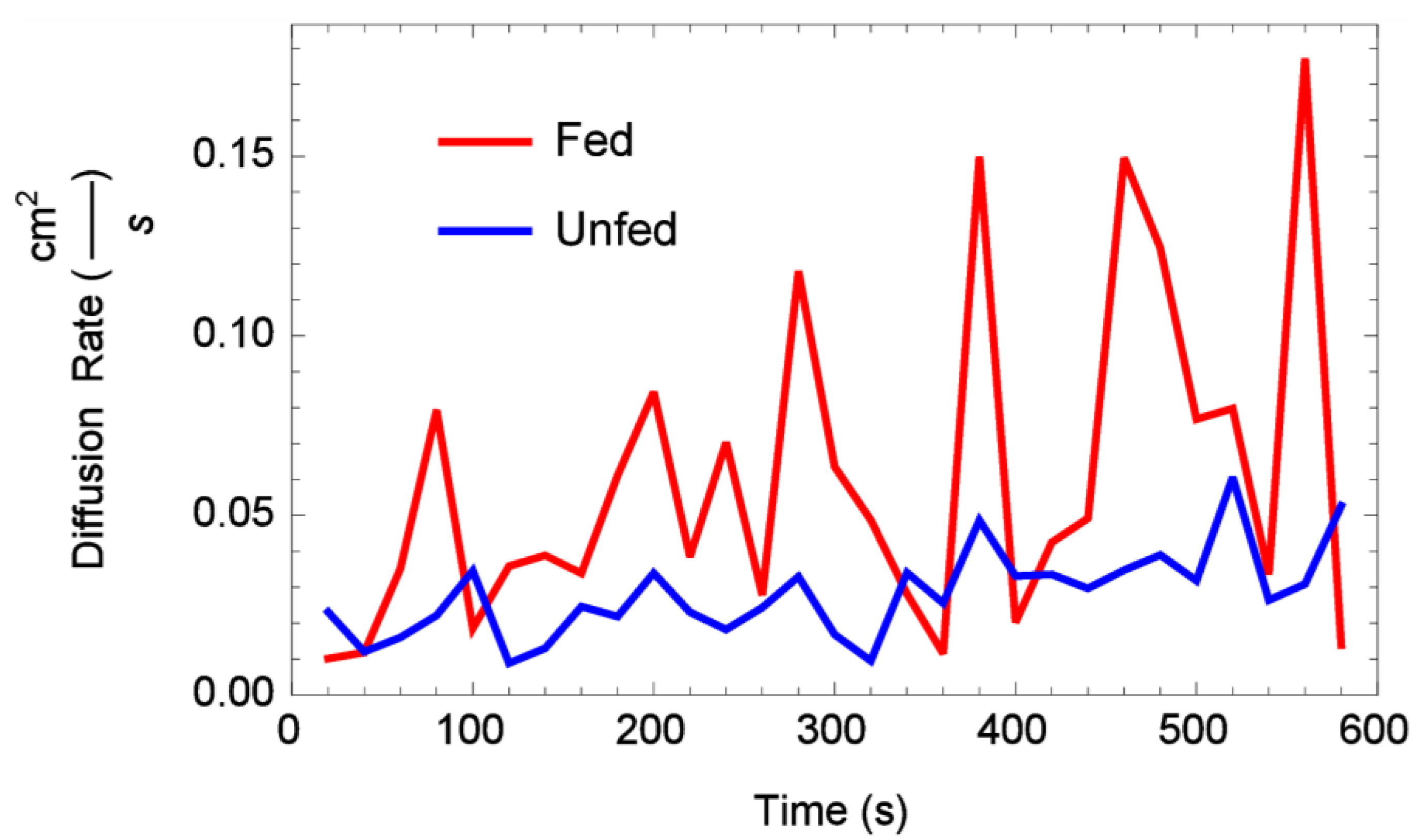
3.3. Diffusion Rates
| Status | Rep 1 | Rep 2 | Rep 3 | Rep 4 | Rep 5 | Overall |
|---|---|---|---|---|---|---|
| one-dimensional fed | 0.040 (0.008) | 0.006 (0.004) | 0.071 (0.076) | 0.079 (0.050) | 0.138 (0.140) | 0.070 (0.051) |
| one dimensional unfed | 0.014 (0.008) | 0.022 (0.011) | 0.060 (0.079) | 0.009 (0.005) | 0.013 (0.008) | 0.024 (0.021) |
| two-dimensional fed | 0.143 (0.146) | 0.006 (0.007) | 0.006 (0.012) | 0.055 (0.063) | 0.059 (0.076) | 0.054 (0.056) |
| two-dimensional unfed | 0.051 (0.068) | 0.017 (0.024) | 0.004 (0.011) | 0.062 (0.055) | 0.023 (0.019) | 0.032 (0.024) |



4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Little, S.E.; West, M.D. Home infestation with Cimex lectularius, the common bed bug, affecting both dog and client (Abstract No. 61). In Proceedings of the American Association of Veterinary Parasitologists Annual Meeting, New Orleans, LA, USA, 19–22 July 2008.
- Reinhardt, K.; Siva-Jothy, M.T. Biology of the bed bugs. Ann. Rev. Entomol. 2007, 52, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Ryckman, R.E.; Bently, D.G.; Archbold, E.F. The Cimicidae of the Americas and oceanic islands, a checklist and bibliography. Bull. Soc. Vector Ecol. 1981, 6, 93–142. [Google Scholar]
- Thomas, I.; Kihiczak, G.G.; Schwartz, R.A. Bedbug bites: A review. Int. J. Dermatol. 2004, 43, 430–459. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.F. The perfect storm: An extension view on bed bugs. Am. Entomol. 2006, 52, 102–104. [Google Scholar] [CrossRef]
- Goddard, J.; de Shazo, R.D. Rapid rise in bed bug populations: The need to include them in the differential diagnosis of mysterious skin rashes. South. Med. J. 2008, 101, 854–855. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.L.; Leffler, K. Bedbug infestations in the news: A picture of an emerging public health problem in the United States. J. Environ. Health 2008, 70, 24–27. [Google Scholar] [PubMed]
- Cleary, C.J.; Buchanan, D. Diagnosis and management of bedbugs: An emerging United States infestation. Nurse Pract. 2004, 29, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Gangloff-Kauffmann, J.; Hollingsworth, C.; Hahn, J.; Hansen, L.; Kard, B.; Waldvogel, M. Bed bugs in America: A pest management industry survey. Pest Control Technol. Mag. 2006, 52, 46–60. [Google Scholar] [CrossRef]
- Bircher, A.J. Systemic immediate allergic reactions to arthropod stings and bites. Dermatology 2005, 210, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Brasch, J.; Schwarz, T. 26-year-old male with urticarial papules. J. Dtsch. Dermatol. Ges. 2006, 4, 1077–1079. [Google Scholar] [CrossRef] [PubMed]
- Churchill, T.P. Urticaria due to bed bug bites. J. Am. Med. Assoc. 1930, 95, 1975–1976. [Google Scholar] [CrossRef]
- De Shazo, R.D.; Feldlaufer, M.F.; Mihm, M.C.; Goddard, J. Bullous reactions to bed bug bites reflect cutaneous vasculitis. Am. J. Med. 2012, 125, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Elston, D.M.; Stockwell, S. What’s eating you? Bed bugs. Cutis 2000, 65, 262–264. [Google Scholar] [PubMed]
- Hwang, S.W.; Svoboda, T.J.; de Jong, I.J.; Kabasele, K.J.; Gogosis, E. Bed bug infestations in an urban environment. Emerg. Infect. Dis. 2005, 11, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Castillo-Neyra, R.; Tustin, A.W.; Borrini-Mayori, K.; Naquira, C.; Levy, M.Z. Bed bugs (Cimex lectularius) as vectors of Trypanosoma cruzi. Am. J. Trop. Med. Hyg. 2015, 92, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J.; de Shazo, R.D. Bed bugs (Cimex lectularius) and clinical consequences of their bites. J. Am. Med. Assoc. 2009, 301, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Mellanby, K. The physiology and activity of the bed bug (Cimex lectularius L.) in a natural setting. Parasitology 1939, 31, 200–211. [Google Scholar] [CrossRef]
- Reis, M.D.; Miller, D.M. Host searching and aggregation activity of recently fed and unfed bed bugs (Cimex lectularius L.). Insects 2011, 2, 186–194. [Google Scholar] [CrossRef]
- Romero, A.; Potter, M.F.; Haynes, K.F. Circadian rhythm of spontaneous locomotor activity in the bed bug, Cimex lectularius. J. Insect Physiol. 2010, 56, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.F.; Moon, R.D.; Kells, S.A. Off-host aggregation behavior and sensory basis of arrestment by Cimex lectularius (heteroptera: Cimicidae). J. Insect Physiol. 2009, 55, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Suchy, J.T.; Lewis, V.R. Host-seeking behavior in the bed bug, Cimex lectularius. Insects 2011, 2, 22–35. [Google Scholar] [CrossRef]
- Turchin, P.; Omland, K. Quantitative analysis of insect movement. In Ecological Entomology, 2nd ed.; Turchin, P., Ed.; Wiley and Sons: New York, NY, USA, 1999; pp. 463–502. [Google Scholar]
- Cantrell, R.S.; Cosner, C. Spatial Ecology via Reaction-Diffusion Equations; Wiley: New York, NY, USA, 2003; p. 411. [Google Scholar]
- Fife, P.C. Mathematical Aspects of Reacting and Diffusing Systems; Springer-Verlag: New York, NY, USA, 1979. [Google Scholar]
- Holmes, E.E.; Lewis, M.A.; Banks, R.R. Partial differential equations in ecology: Spatial interactions and population dynamics. Ecology 1994, 75, 17–29. [Google Scholar] [CrossRef]
- Levin, S.A. Dispersion and population interactions. Am. Nat. 1974, 108, 207–228. [Google Scholar] [CrossRef]
- Levin, S.A. The role of theoretical ecology in the description and understanding of populations in heterogeneous environments. Amer. Zool. 1981, 21, 865–875. [Google Scholar] [CrossRef]
- Murray, J.D. An introduction. In Mathematical Biology, 3rd ed.; Springer-Verlag: New York, NY, USA, 2003; Volume I. [Google Scholar]
- Okubo, A. Diffusion and Ecological Problems: Mathematical Models; Springer: Berlin, Germany, 1980; Volume 10. [Google Scholar]
- Turchin, P. Quantitative Analysis of Movement: Measuring and Modeling Population Redistribution in Animals and Plants; Sinauer Associates Sunderland: Sunderland, MA, USA, 1998. [Google Scholar]
- Kareiva, P.M. Local movement in herbivorous insects: Applying a passive diffusion model to mark-recapture field experiments. Oecologia 1983, 57, 322–327. [Google Scholar] [CrossRef]
- Bowman, A.W.; Azzalini, A. Applied Smoothing Techniques for Data Analysis: The Kernal Approach, with S-Plus Illustrations; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Langlois, T.J.; Fitzpatrick, B.R.; Fairclough, D.V.; EWakefield, C.B.; Hesp, S.A.; McLean, D.L.; Harvey, E.S.; Meeuwig, J.J. Similarities between line fishing and baited stereo-video estimations of length-frequency: Novel application of kernal density estimates. PLoS ONE 2012, 7, e45973. [Google Scholar] [CrossRef] [PubMed]
- How, Y.F.; Lee, C.Y. Effects of life stages and feeding regimes on active movement behavior of the tropical bed bug, Cimex hemipterus (hemiptera: Cimicidae). J. Med. Entomol. 2010, 47, 305–312. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goddard, J.; Caprio, M.; Goddard, J., II. Diffusion Rates and Dispersal Patterns of Unfed versus Recently Fed Bed Bugs (Cimex lectularius L.). Insects 2015, 6, 792-804. https://doi.org/10.3390/insects6040792
Goddard J, Caprio M, Goddard J II. Diffusion Rates and Dispersal Patterns of Unfed versus Recently Fed Bed Bugs (Cimex lectularius L.). Insects. 2015; 6(4):792-804. https://doi.org/10.3390/insects6040792
Chicago/Turabian StyleGoddard, Jerome, Michael Caprio, and Jerome Goddard, II. 2015. "Diffusion Rates and Dispersal Patterns of Unfed versus Recently Fed Bed Bugs (Cimex lectularius L.)" Insects 6, no. 4: 792-804. https://doi.org/10.3390/insects6040792