
Effects of Rearing Density on Survival, Growth, and Development of the Ladybird Coleomegilla maculata in Culture



Abstract
:1. Introduction
2. Experimental Section
2.1. Research Subjects and Food Sources
2.2. Rearing Density
2.3. Statistical Analysis
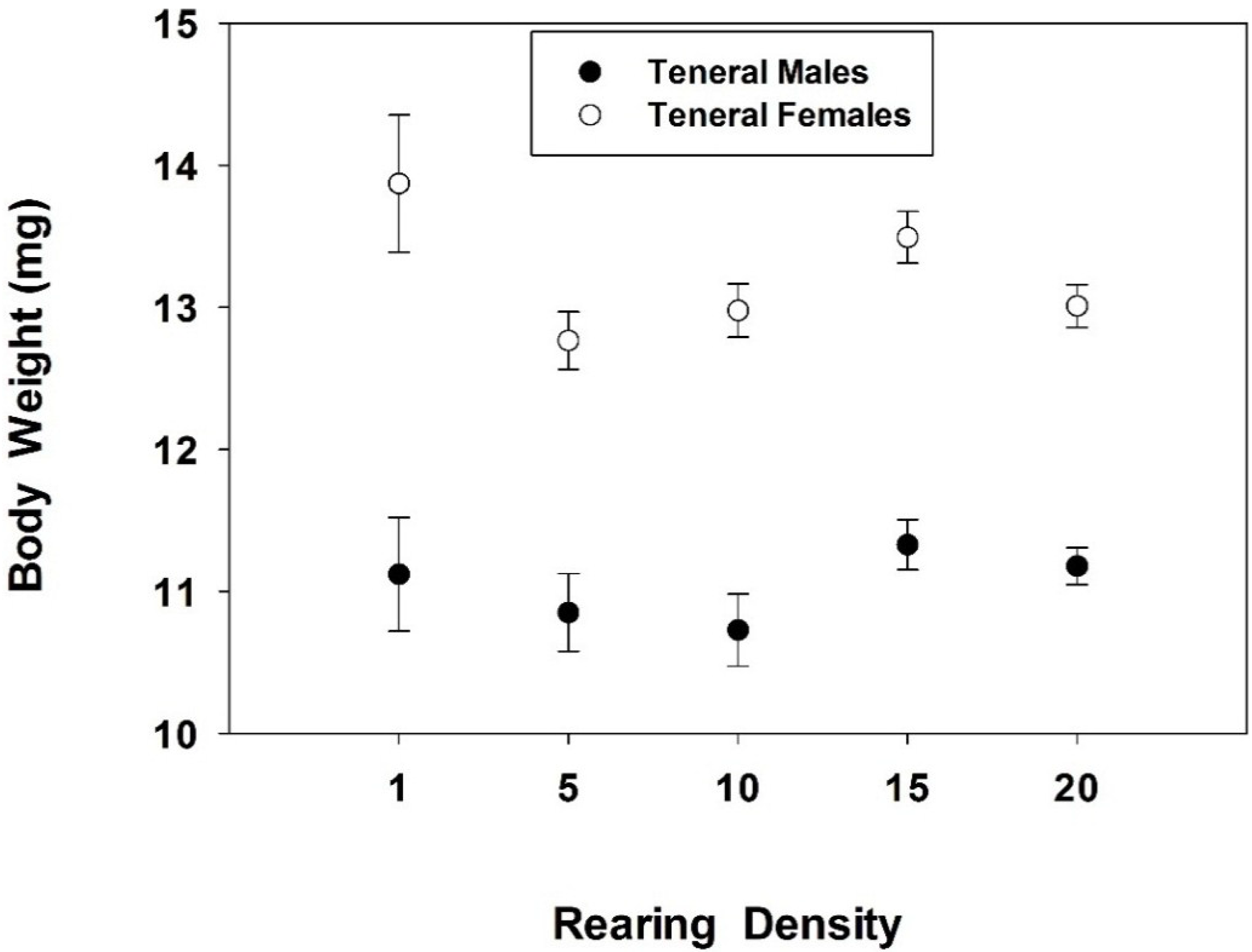
3. Results

{kind=link}
{kind=link}
| Life Parameter | Rearing Density | Median | 10% C.I. | 25% C.I. | 75% C.I. | 90% C.I. |
|---|---|---|---|---|---|---|
| Larval Survival (%) | 1 | 100 a | 100 | 100 | 100 | 100 |
| 5 | 100 ab | 80.0 | 100 | 100 | 100 | |
| 10 | 90.0 bc | 80.0 | 85.0 | 95.0 | 100 | |
| 15 | 93.3 bc | 80.0 | 88.3 | 100 | 95 | |
| 20 | 95.0 c | 85.0 | 92.5 | 95.0 | 100 | |
| Time as Larva (days) | 1 | 14.0 a | 11.0 | 11.5 | 14.0 | 14.0 |
| 5 | 13.0 a | 11.8 | 12.1 | 13.8 | 14.0 | |
| 10 | 13.7 a | 11.8 | 12.3 | 14.0 | 14.4 | |
| 15 | 13.6 a | 12.8 | 13.1 | 14.0 | 14.2 | |
| 20 | 13.2 a | 12.0 | 12.2 | 13.8 | 14.0 | |
| Time as Pupa (days) | 1 | 3.0 a | 2.0 | 2.0 | 4.0 | 5.0 |
| 5 | 4.0 a | 3.6 | 3.9 | 4.9 | 5.0 | |
| 10 | 3.9 a | 3.2 | 3.4 | 4.4 | 4.7 | |
| 15 | 3.8 a | 3.5 | 3.7 | 4.2 | 4.5 | |
| 20 | 4.0 a | 3.6 | 3.8 | 4.7 | 4.7 | |
| Pupal Survival (%) | 1 | 100 | 100 | 100 | 100 | 100 |
| 5 | 100 | 100 | 100 | 100 | 100 | |
| 10 | 100 | 100 | 100 | 100 | 100 | |
| 15 | 100 | 100 | 100 | 100 | 100 | |
| 20 | 100 | 100 | 100 | 100 | 100 | |
| Sex Ratio (% ♀♀) | 1 | 100 a | 0 | 0 | 100 | 100 |
| 5 | 50.0 a | 40.0 | 40.0 | 60.0 | 80.0 | |
| 10 | 55.6 a | 33.3 | 47.2 | 64.6 | 66.7 | |
| 15 | 50.0 a | 42.9 | 46.7 | 53.6 | 53.8 | |
| 20 | 52.6 a | 42.1 | 45.7 | 55.3 | 63.2 |


4. Discussion
| Predator | Larvae per Arena at 70%+ Survival Rate * | Arena Height (h) × Diameter (d), in cm | Arena Volume (cm3) 1 | Successful Rearing Density (Larvae/cm3) | Body Weight (mg), at 70%+ * | Reference |
|---|---|---|---|---|---|---|
| Coleomegilla maculata | 20 | 2.5 × 9.0 | 158.96 | 0.126 | 13.0 | This study |
| Brumoides suturalis | 10 | 1.5 × 14.0 | 230.79 | 0.043 | - | [26] |
| Adalia bipunctata | 4 | 20 × 15 | 3,532 | 0.001 | - | [24] |
| Propylea dissecta | 4 | 11 × 9 | 699.43 | 0.006 | 11.2 | [25] |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gordon, R.D. The Coccinellidae (Coleoptera) of America north of Mexico. J. N.Y. Entomol. Soc. 1985, 93, 1–912. [Google Scholar]
- Coll, M.; Mendoza, L.C.; Roderick, G.K. Population structure of a predatory beetle: The importance of gene flow for intertrophic level interactions. Heredity 1994, 72, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Munyaneza, J.; Obrycki, J.J. Development of three populations of Coleomegilla maculata (Coleoptera: Coccinellidae) feeding on eggs of Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 1998, 27, 117–122. [Google Scholar] [CrossRef]
- Krafsur, E.S.; Obrycki, J.J. Coleomegilla maculata (Coleoptera: Coccinellidae) is a species complex. Ann. Entomol. Soc. Am. 2000, 93, 1156–1163. [Google Scholar] [CrossRef]
- Conrad, M.S. The spotted lady beetle, Coleomegilla maculata (De Geer) as a predator of European corn borer eggs. J. Econ. Entomol. 1959, 52, 843–847. [Google Scholar] [CrossRef]
- Michaud, J.P.; Jyoti, J.L. Dietary complementation across life stages in the polyphagous lady beetle Coleomegilla maculata. Entomol. Exp. Appl. 2008, 126, 40–45. [Google Scholar]
- Hodek, I.; Evans, E.W. Food relationships, Chapter 5. In Ecology and Behaviour of Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Blackwell Publishing Ltd.: Chichester, West Sussex, UK, 2012; pp. 141–274. [Google Scholar]
- Smith, B.C. A technique for rearing coccinellid beetles on dry foods, and influence of various pollens on the development of Coleomegilla maculata lengi Timb. Can. J. Zool. 1960, 38, 1047–1049. [Google Scholar] [CrossRef]
- Smith, B.C. Growth and development of coccinellid larvae on dry foods (Coleoptera: Coccinellidae). Can. Entomol. 1965, 97, 760–768. [Google Scholar] [CrossRef]
- Andow, D.A.; Risch, S.J. Predation in diversified agroecosystems: Relations between a coccinellid predator Coleomegilla maculata and its food. J. Appl. Ecol. 1985, 22, 357–372. [Google Scholar] [CrossRef]
- Phoofolo, M.W.; Obrycki, J.J.; Lewis, L.C. Quantitative assessment of biotic mortality factors of the European corn borer (Lepidoptera: Crambidae) in field corn. J. Econ. Entomol. 2001, 94, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Musser, F.R.; Shelton, A.M. Predation of Ostrinia nubilalis (Lepidoptera: Crambidae) eggs in sweet corn by generalist predators and the impact of alternative foods. Environ. Entomol. 2003, 32, 1131–1138. [Google Scholar] [CrossRef]
- Li, Y.; Ostrem, J.; Romeis, J.; Chen, M.; Liu, X.; Hellmich, R.L.; Shelton, A.M.; Peng, Y. Development of a tier-1 assay for assessing the toxicity of insecticidal substances against Coleomegilla maculata. Environ. Entomol. 2011, 40, 496–502. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Rojas, M.G. Is Tetranychus urticae suitable prey for development and reproduction of naïve Coleomegilla maculata? Insect Sci. 2014, 21, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Riddick, E.W.; Wu, Z.; Rojas, M.G. Potential utilization of Artemia franciscana eggs as food for Coleomegilla maculata. BioControl 2014, 59, 575–583. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Rojas, M.G.; Morales-Ramos, J.A. Potential utilization of Spirulina microalga as a dietary supplement for the ladybird beetle Coleomegilla maculata. Trends Entomol. 2014, 10, 39–48. [Google Scholar]
- Peters, T.M.; Barbosa, P. Influence of population density on size, fecundity, and developmental rate of insects in culture. Annu. Rev. Entomol. 1972, 22, 431–450. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Andow, D.A. Mortality of coccinellid (Coleoptera: Coccinellidae) larvae and pupae when prey become scarce. Environ. Entomol. 1999, 28, 1092–1100. [Google Scholar] [CrossRef]
- Michaud, J.P. A comparative study of larval cannibalism in three species of ladybird. Ecol. Entomol. 2003, 28, 92–101. [Google Scholar] [CrossRef]
- Pervez, A.; Gupta, A.K.; Omkar. Larval cannibalism in aphidophagous ladybirds: Influencing factors, benefits and costs. Biol. Control 2006, 38, 307–313. [Google Scholar] [CrossRef]
- Hesler, L.S.; McNickle, G.; Catangui, M.A.; Losey, J.E.; Beckendorf, E.A.; Stellwag, L.; Brandt, D.M.; Bartlett, P.B. Method for continuously rearing Coccinella lady beetles (Coleoptera: Coccinellidae). Open Entomol. J. 2012, 6, 42–48. [Google Scholar] [CrossRef]
- Allen, M.A.; Riddick, E.W. A system for harvesting eggs from the pink-spotted lady beetle. Psyche 2012. [Google Scholar] [CrossRef]
- Riddick, E.W.; Chen, H. Production of coleopteran predators, Chapter 2. In Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; Morales-Ramos, J.A., Rojas, M.G., Shapiro-Ilan, D.I., Eds.; Academic Press: London, UK, 2014; pp. 17–55. [Google Scholar]
- Dimetry, N.Z. The role of predator and prey density as factors affecting behavioural and biological aspects of Adalia bipunctata (L.) larvae. Z. Ang. Entomol. 1976, 81, 386–392. [Google Scholar] [CrossRef]
- Omkar; Pathak, S. Crowding affects the life attributes of an aphidophagous ladybird beetle, Propylea dissecta. Bull. Insectol. 2009, 62, 35–40. [Google Scholar]
- Bista, M.; Mishra, G.; Omkar. Influence of crowding and diet on the development and survival of the ladybird Brumoides suturalis (Coleoptera: Coccinellidae) reared on two aphid species. Int. J. Trop. Insect Sci. 2012, 32, 64–68. [Google Scholar] [CrossRef]
- Bonte, M.; de Clercq, P. Influence of predator density, diet and living substrate on developmental fitness of Orius laevigatus. J. Appl. Entomol. 2011, 135, 343–350. [Google Scholar] [CrossRef]
- Grundy, P.R.; Maelzer, D.A.; Bruce, A.; Hassan, E. A mass-rearing method for the assassin bug Pristhesancus plagipennis (Hemiptera: Reduviidae). Biol. Control 2000, 18, 243–250. [Google Scholar] [CrossRef]
- Sahayaraj, K. Small scale laboratory rearing of a reduviid predator, Rhynocoris marginatus Fab. (Hemiptera: Reduviidae) on Corcyra cephalonica Stainton larvae by larval card method. J. Central Eur. Agric. 2002, 3, 137–148. [Google Scholar]
- Yu, J.-Z.; Chen, B.-H. Effect of concealment and rearing density on the development and survival of Lemnia biplagiata (Coleoptera: Coccinellidae). J. Agric. Res. China 2001, 50, 68–74. (In Chinese) [Google Scholar]
- Shands, W.A.; Shands, M.K.; Simpson, G.W. Techniques for mass-producing Coccinella septempunctata. J. Econ. Entomol. 1966, 59, 1022–1023. [Google Scholar] [CrossRef]
- Ito, K. A simple mass-rearing method for predaceous Orius bugs in the laboratory. Appl. Entomol. Zool. 2007, 42, 573–577. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z. Does a change from whole to powdered food (Artemia franciscana eggs) increase oviposition in the ladybird Coleomegilla maculata? Insects 2015, 6, 815–826. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Dixon, A.F.G. Insect Predator-Prey Dynamics: Ladybird Beetles and Biological Control; Cambridge Univ. Press: Cambridge, UK, 2000. [Google Scholar]
- Riddick, E.W.; Wu, Z.; Jamie Whitten Delta States Research Center, ARS-USDA, Stoneville, MS, USA. Unpublished data. 2015.
- De Clercq, P.; Bonte, M.; van Speybroeck, K.; Bolckmans, K.; Deforce, K. Development and reproduction of Adalia bipunctata (Coleoptera: Coccinellidae) on eggs of Ephestia kuehniella (Lepidoptera: Phycitidae) and pollen. Pest Manage. Sci. 2005, 61, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, D. Capacity and quality: Keys to success in the mass rearing of biological control agents. Nat. Enemies Ins. 1998, 20, 169–179. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riddick, E.W.; Wu, Z. Effects of Rearing Density on Survival, Growth, and Development of the Ladybird Coleomegilla maculata in Culture. Insects 2015, 6, 858-868. https://doi.org/10.3390/insects6040858
Riddick EW, Wu Z. Effects of Rearing Density on Survival, Growth, and Development of the Ladybird Coleomegilla maculata in Culture. Insects. 2015; 6(4):858-868. https://doi.org/10.3390/insects6040858
Chicago/Turabian StyleRiddick, Eric W., and Zhixin Wu. 2015. "Effects of Rearing Density on Survival, Growth, and Development of the Ladybird Coleomegilla maculata in Culture" Insects 6, no. 4: 858-868. https://doi.org/10.3390/insects6040858
APA StyleRiddick, E. W., & Wu, Z. (2015). Effects of Rearing Density on Survival, Growth, and Development of the Ladybird Coleomegilla maculata in Culture. Insects, 6(4), 858-868. https://doi.org/10.3390/insects6040858