
Bee Community of Commercial Potato Fields in Michigan and Bombus impatiens Visitation to Neonicotinoid-Treated Potato Plants




Abstract
:1. Introduction
2. Materials and Methods
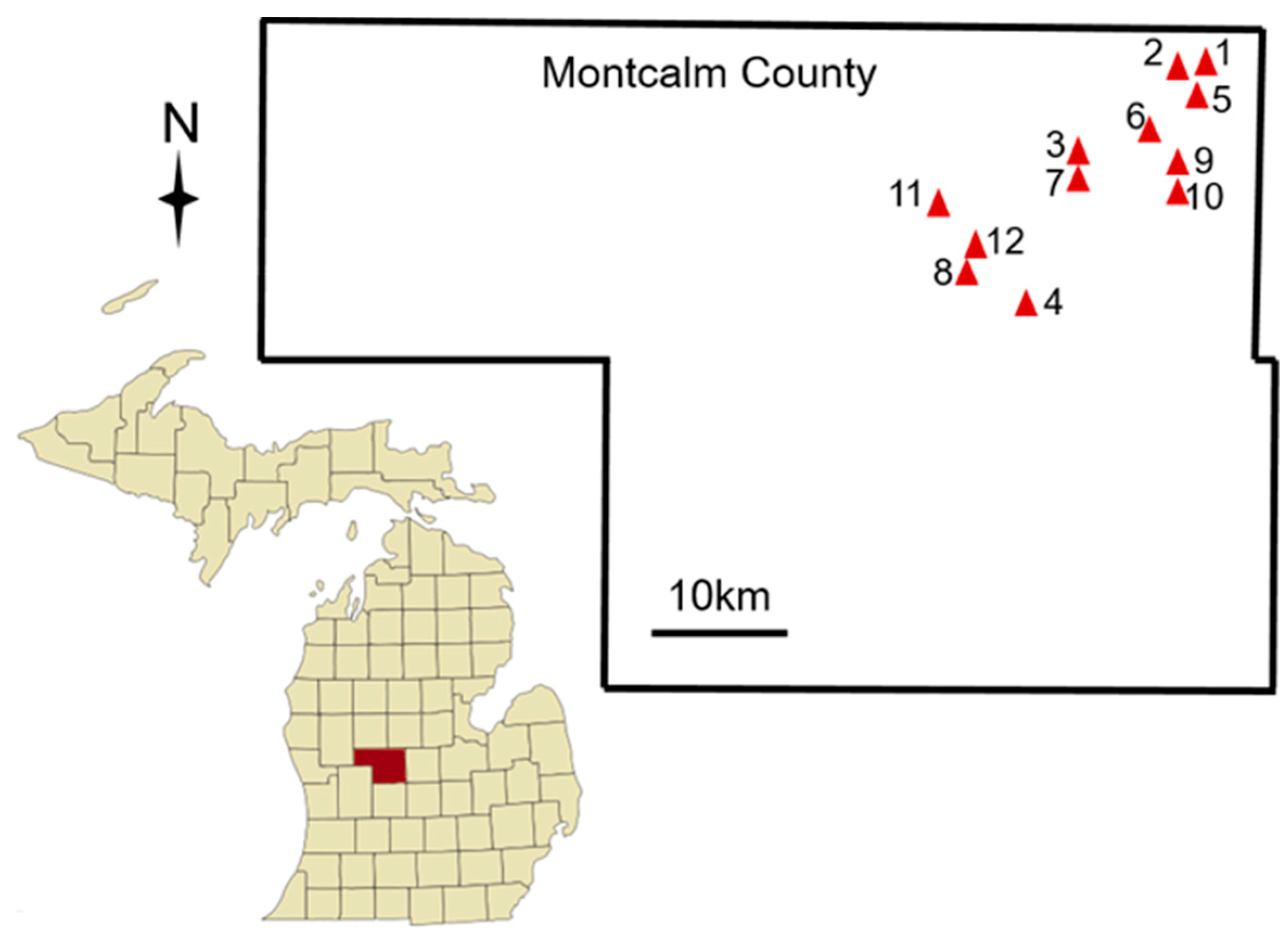
2.1. Bee Community Survey in Neonicotinoid-Treated Commercial Potato Fields
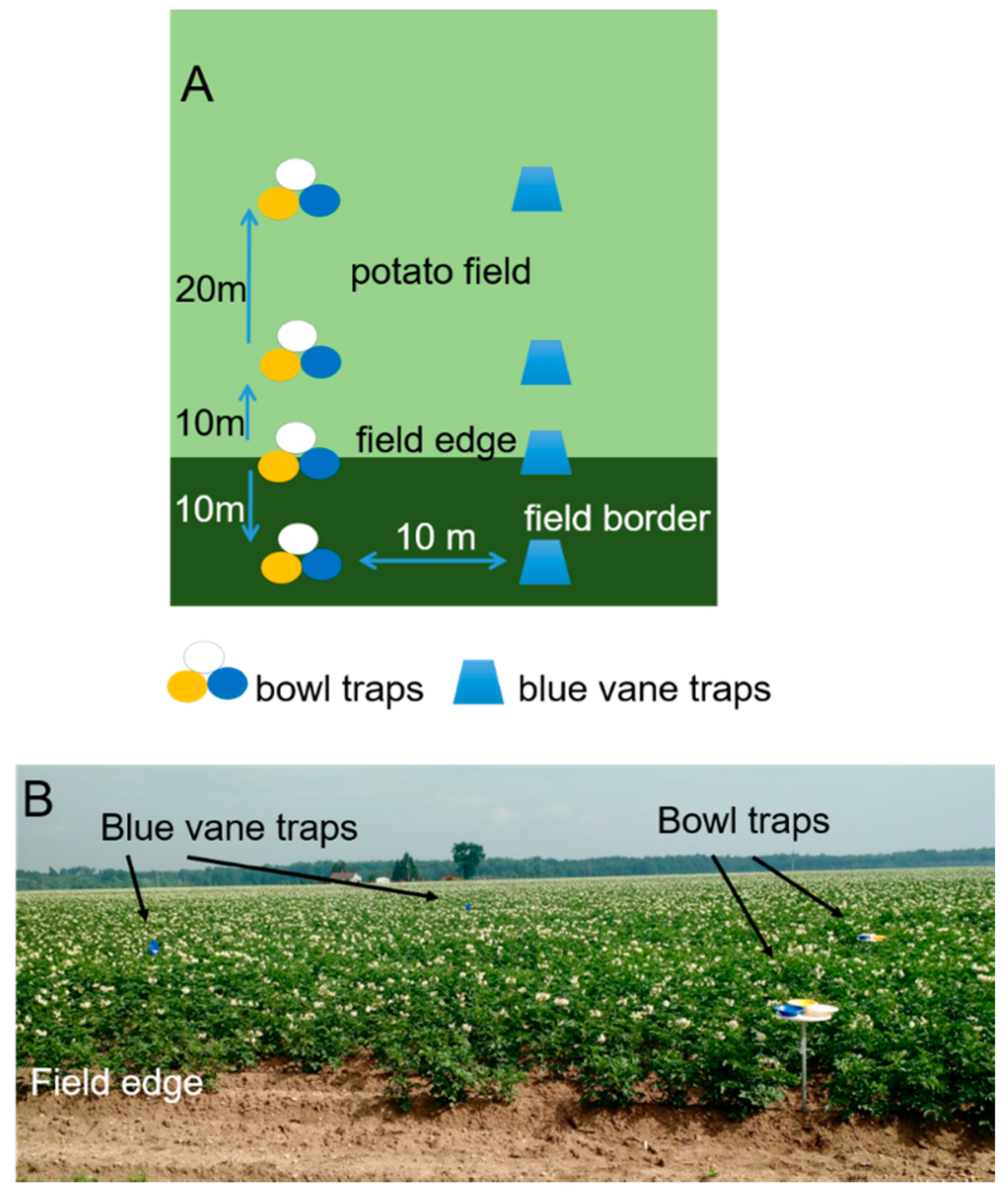
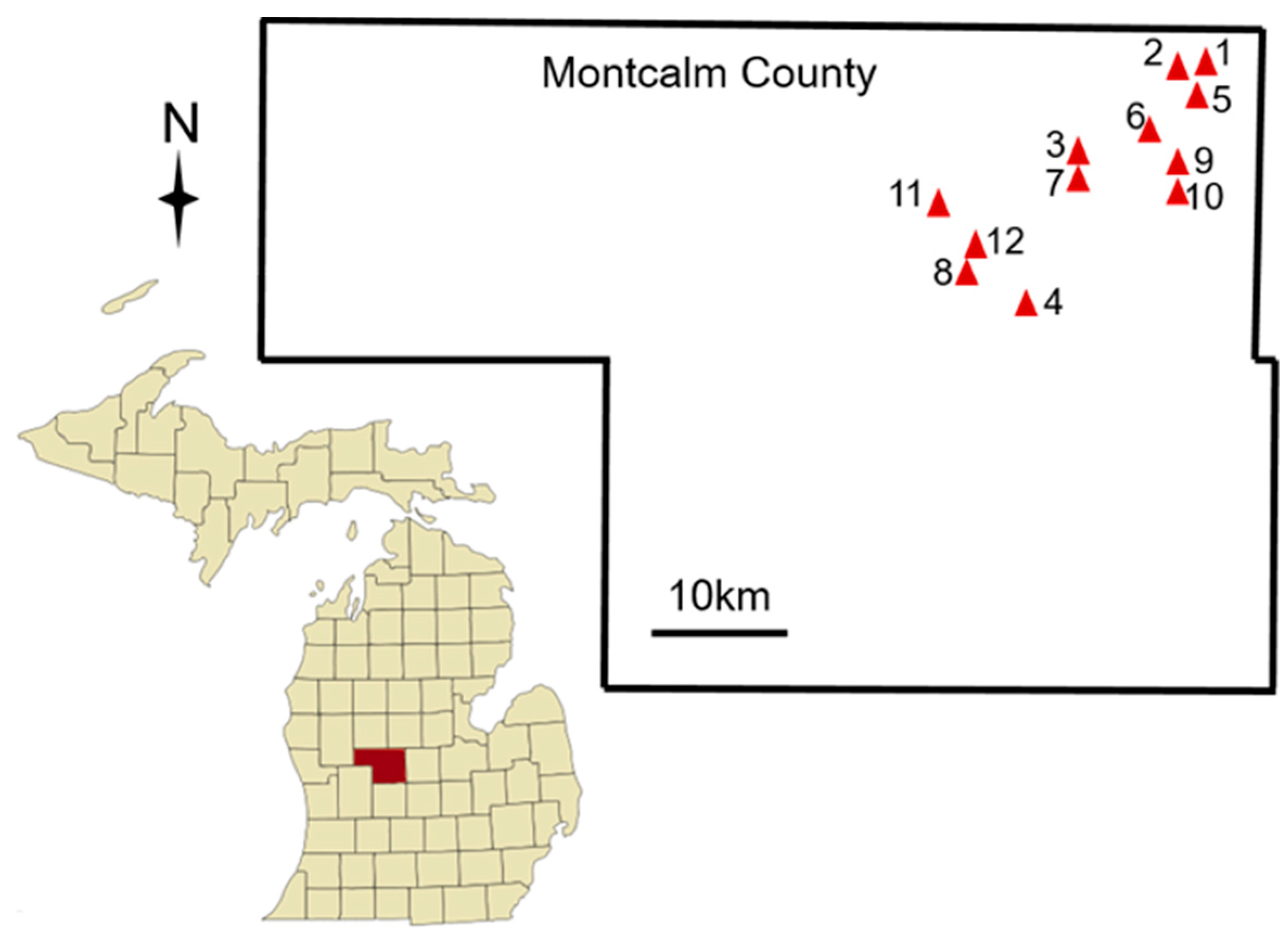
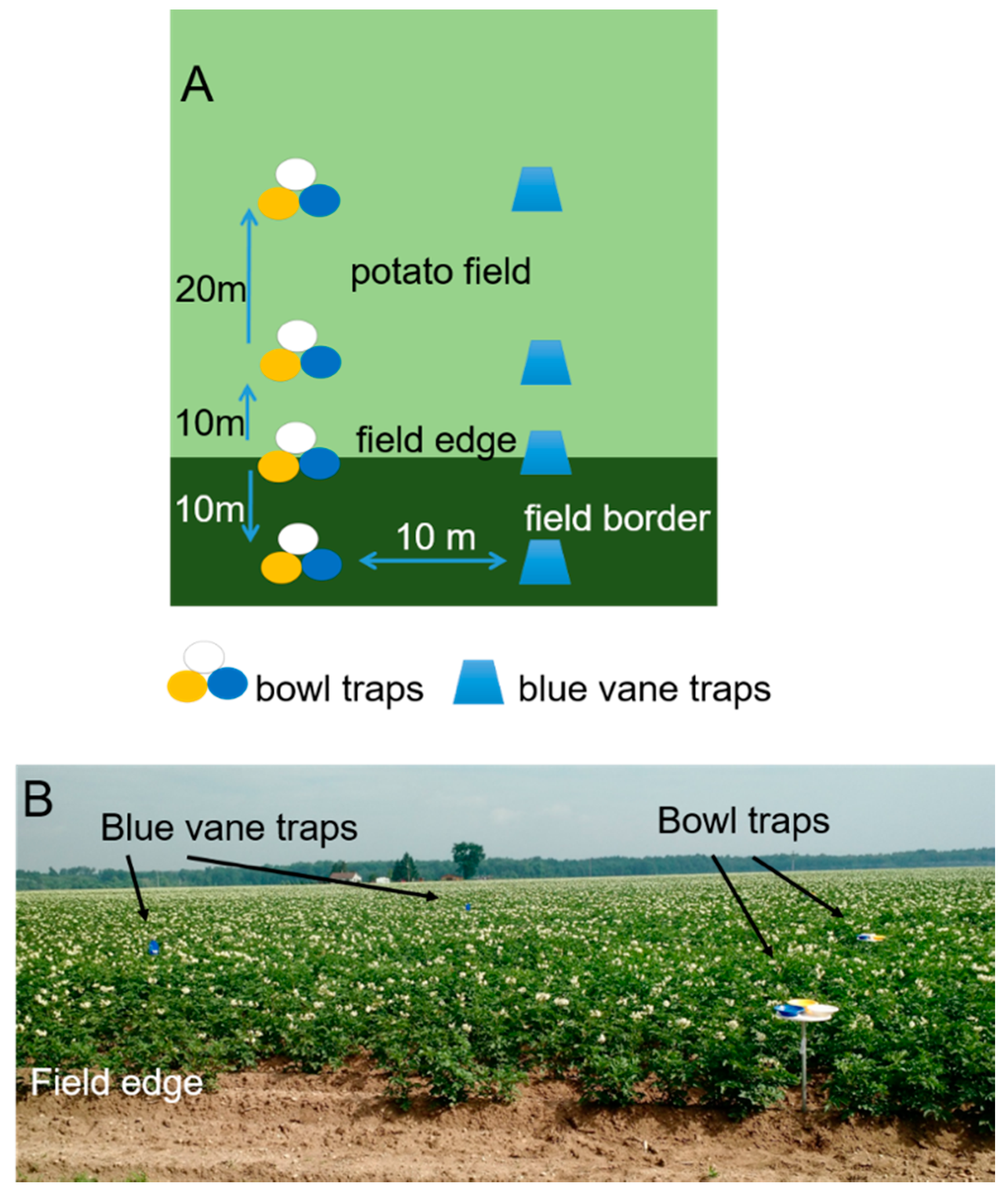
2.1.1. Data Collection
2.1.2. Statistical Analysis
2.2. Bumblebee Potato Flower Visitation
2.2.1. Data Collection
2.2.2. Statistical Analysis
3. Results
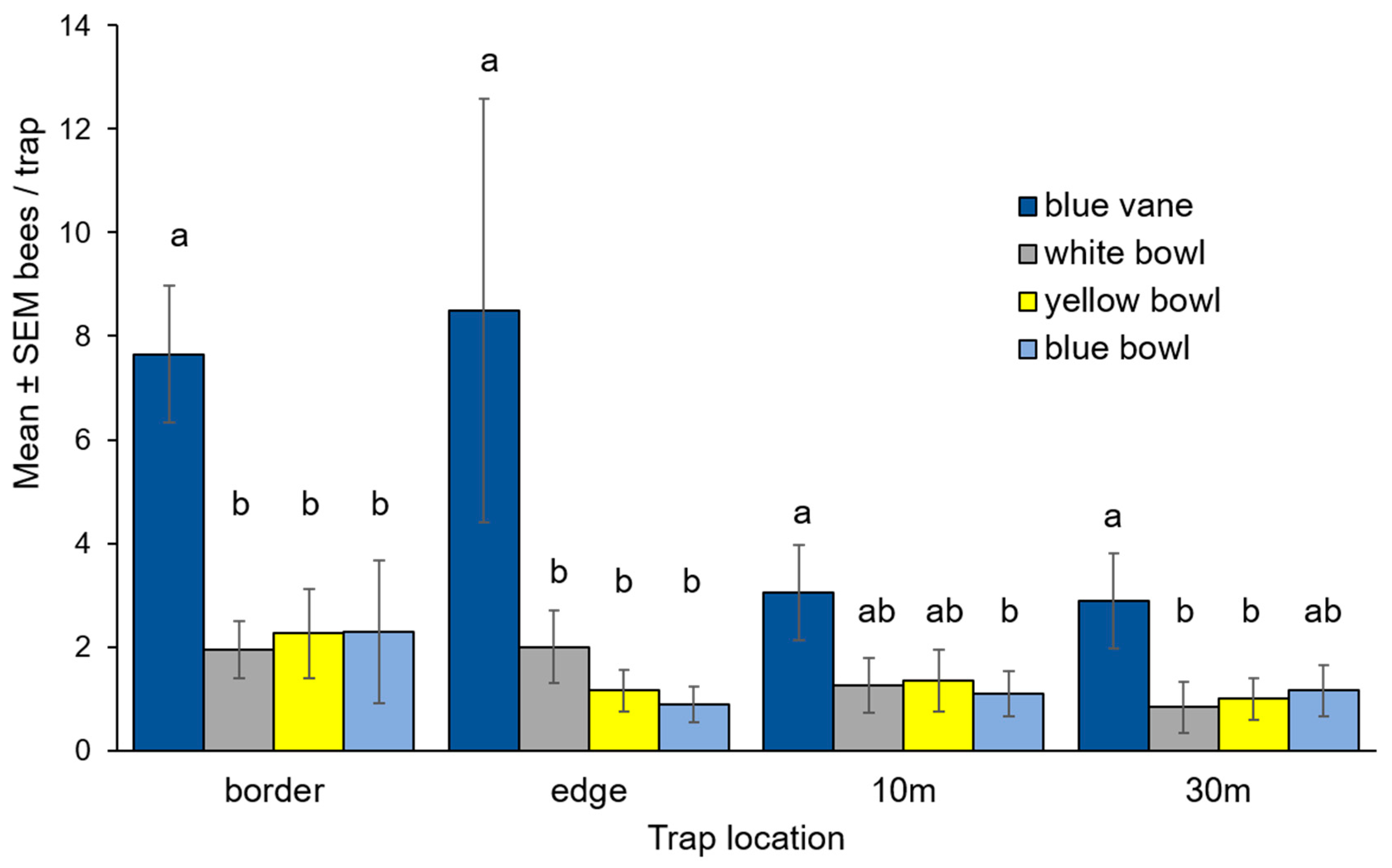
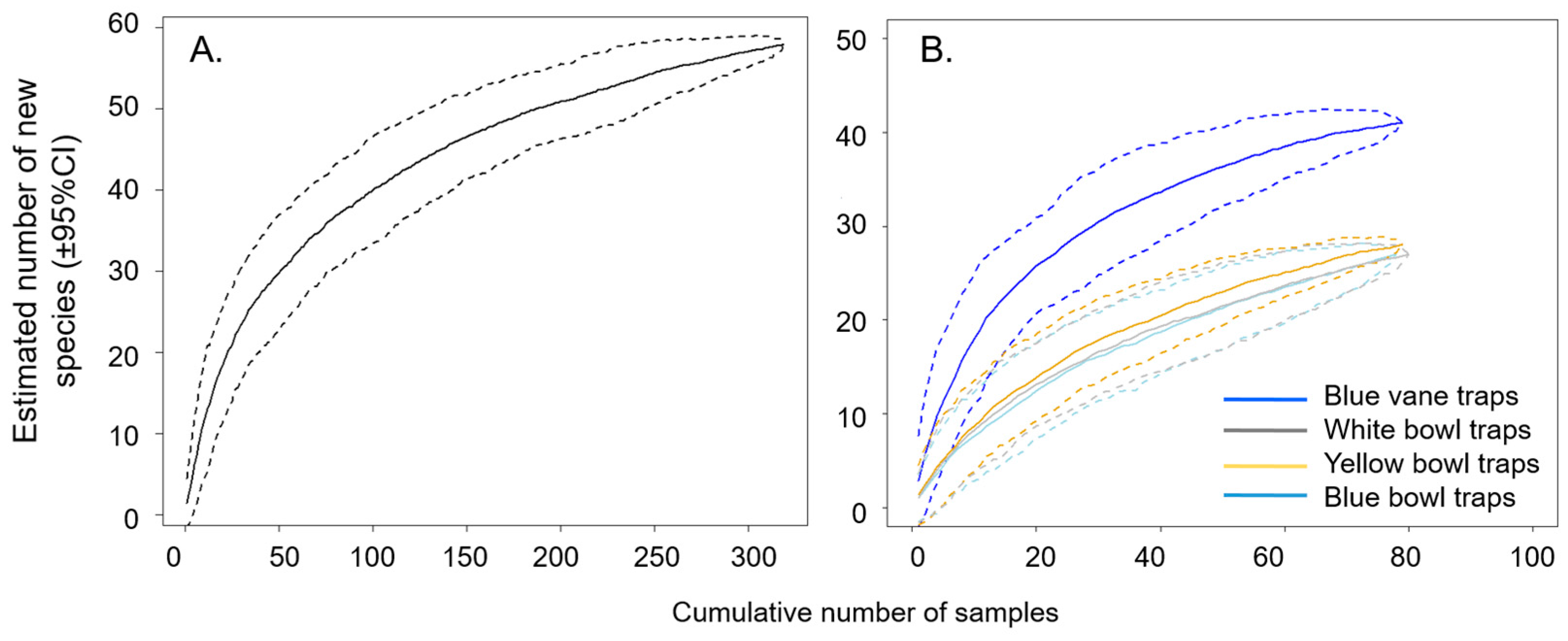
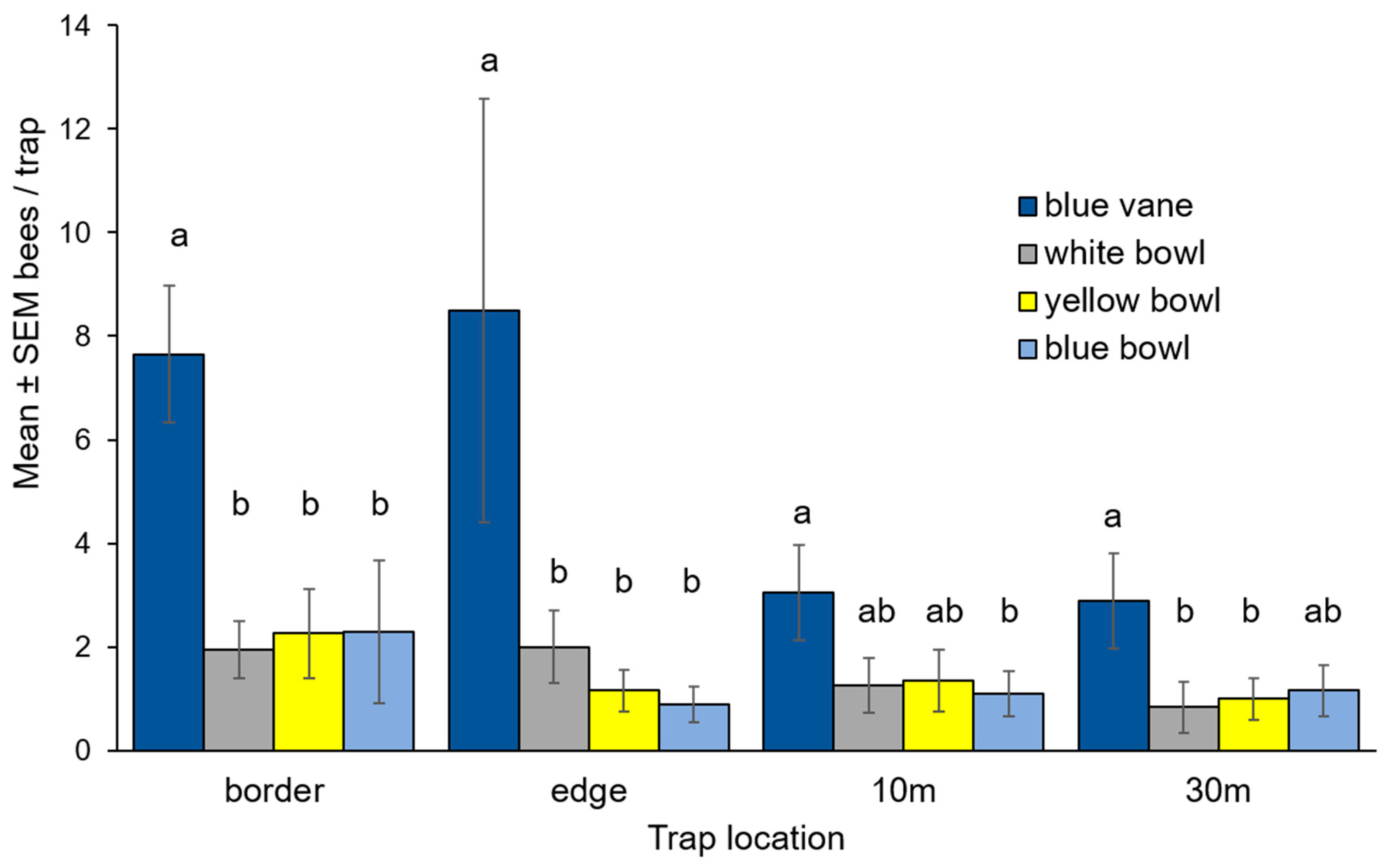
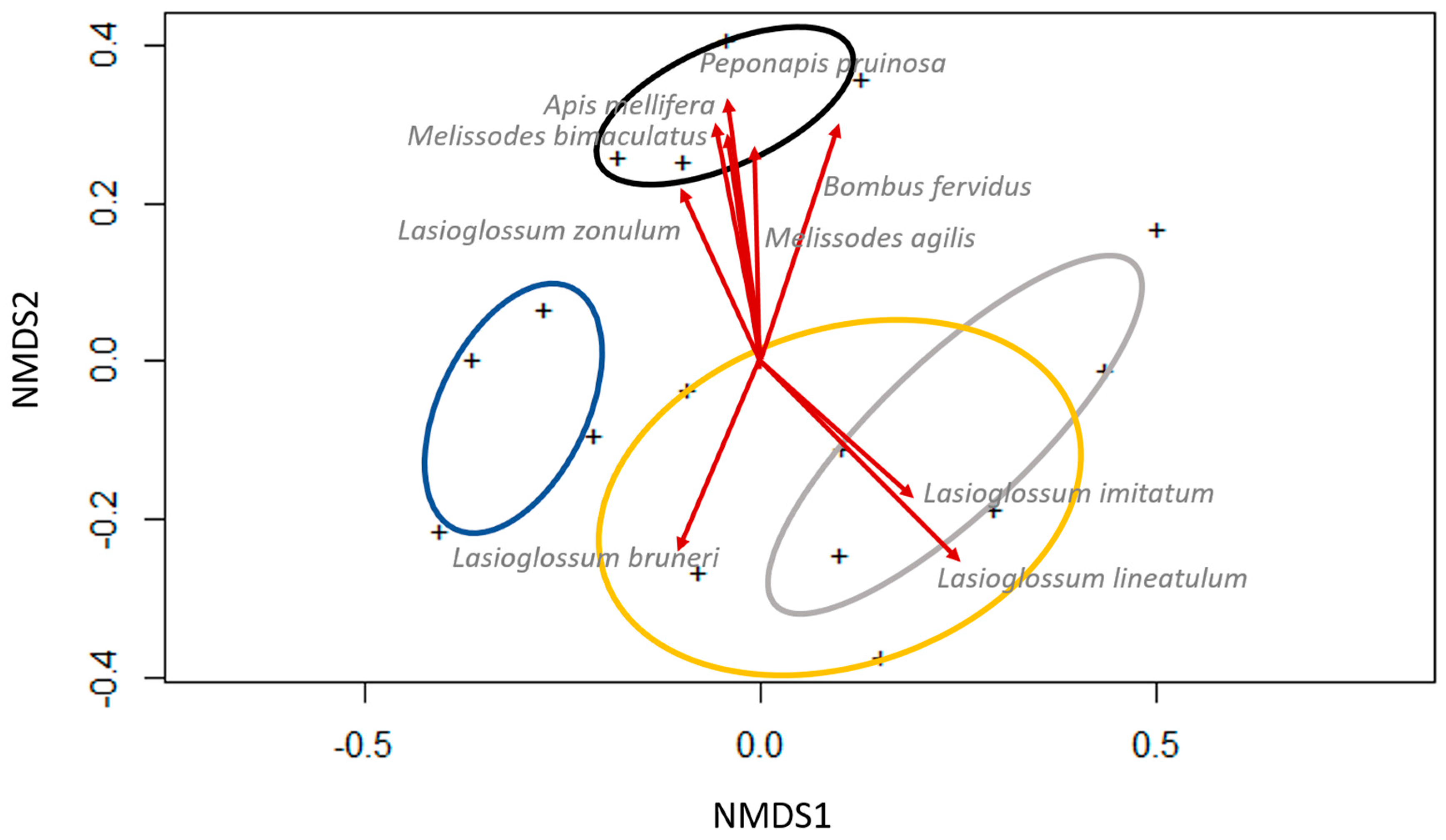
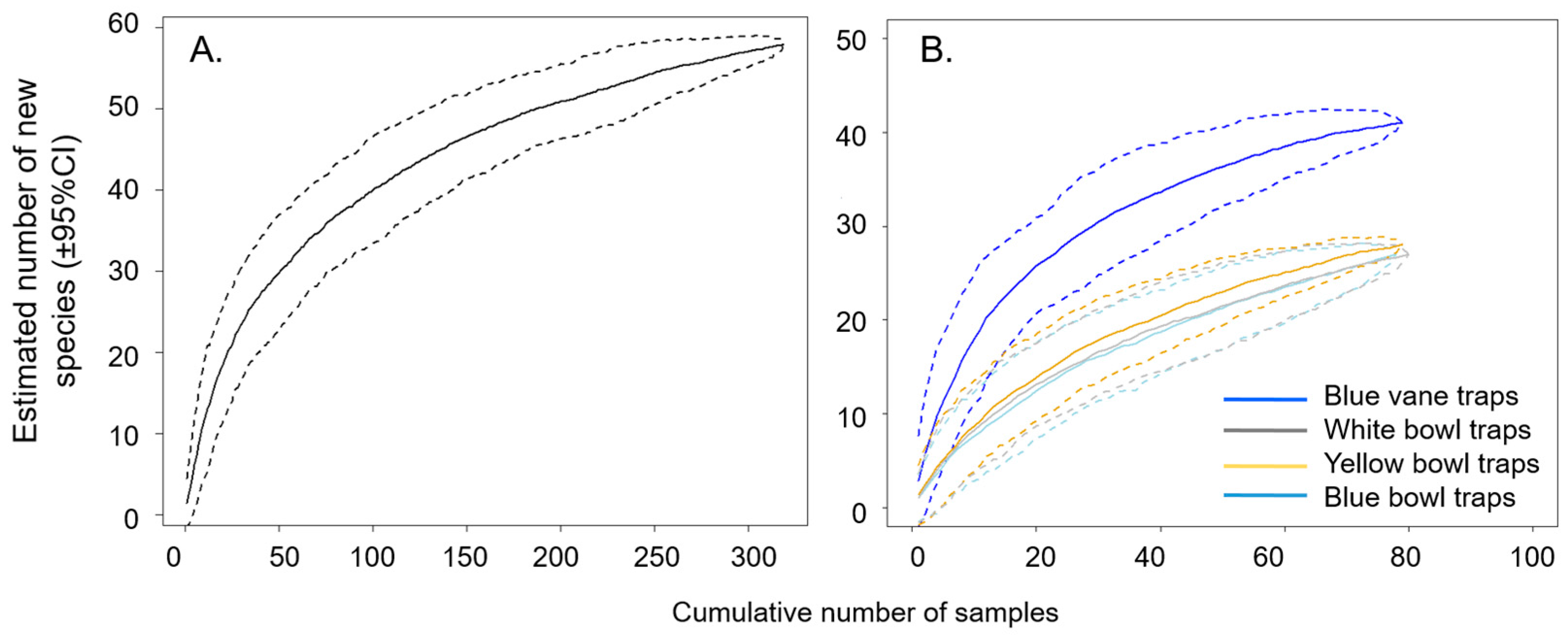
3.1. Bee Community Survey in Commercial Potato Fields
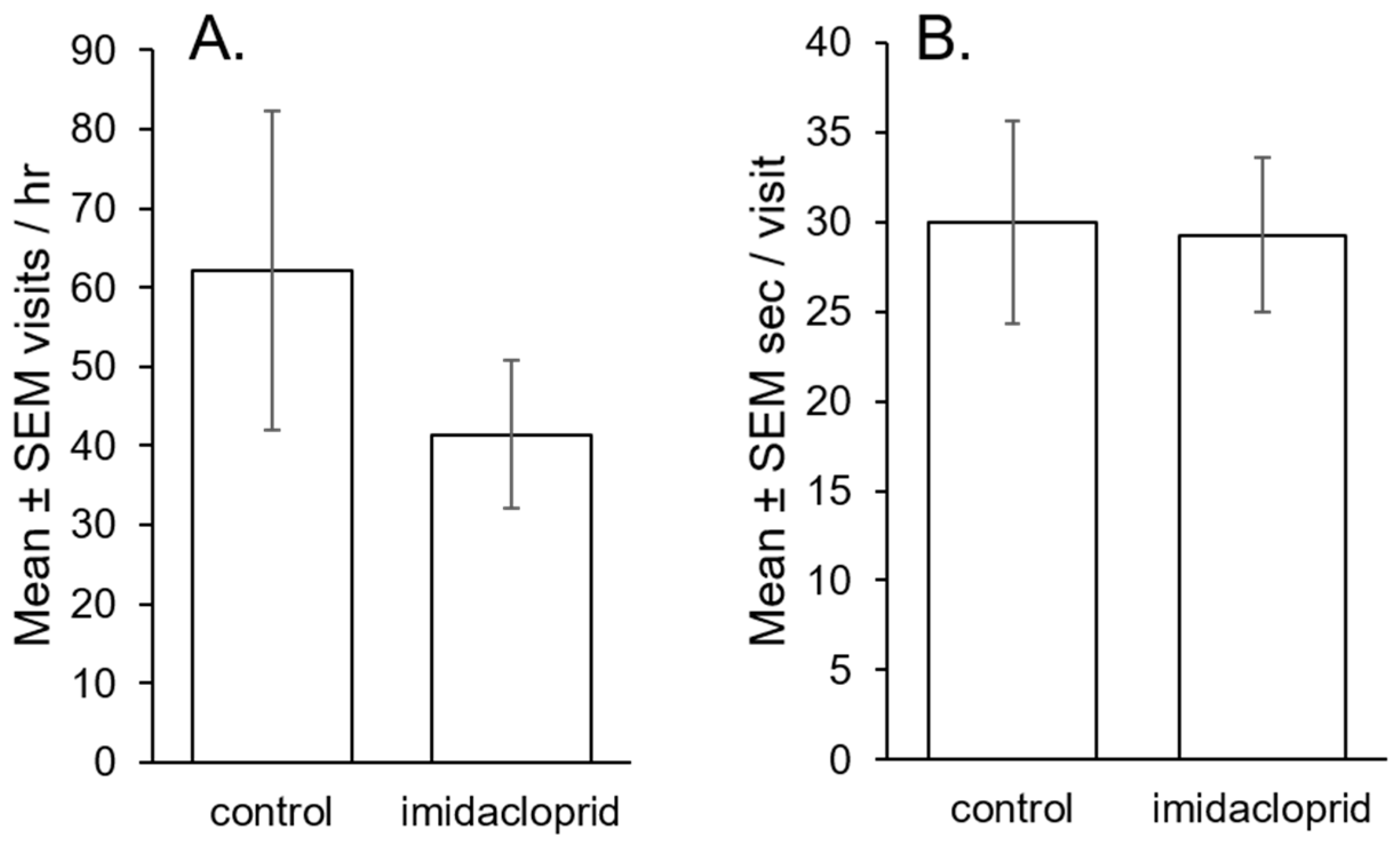
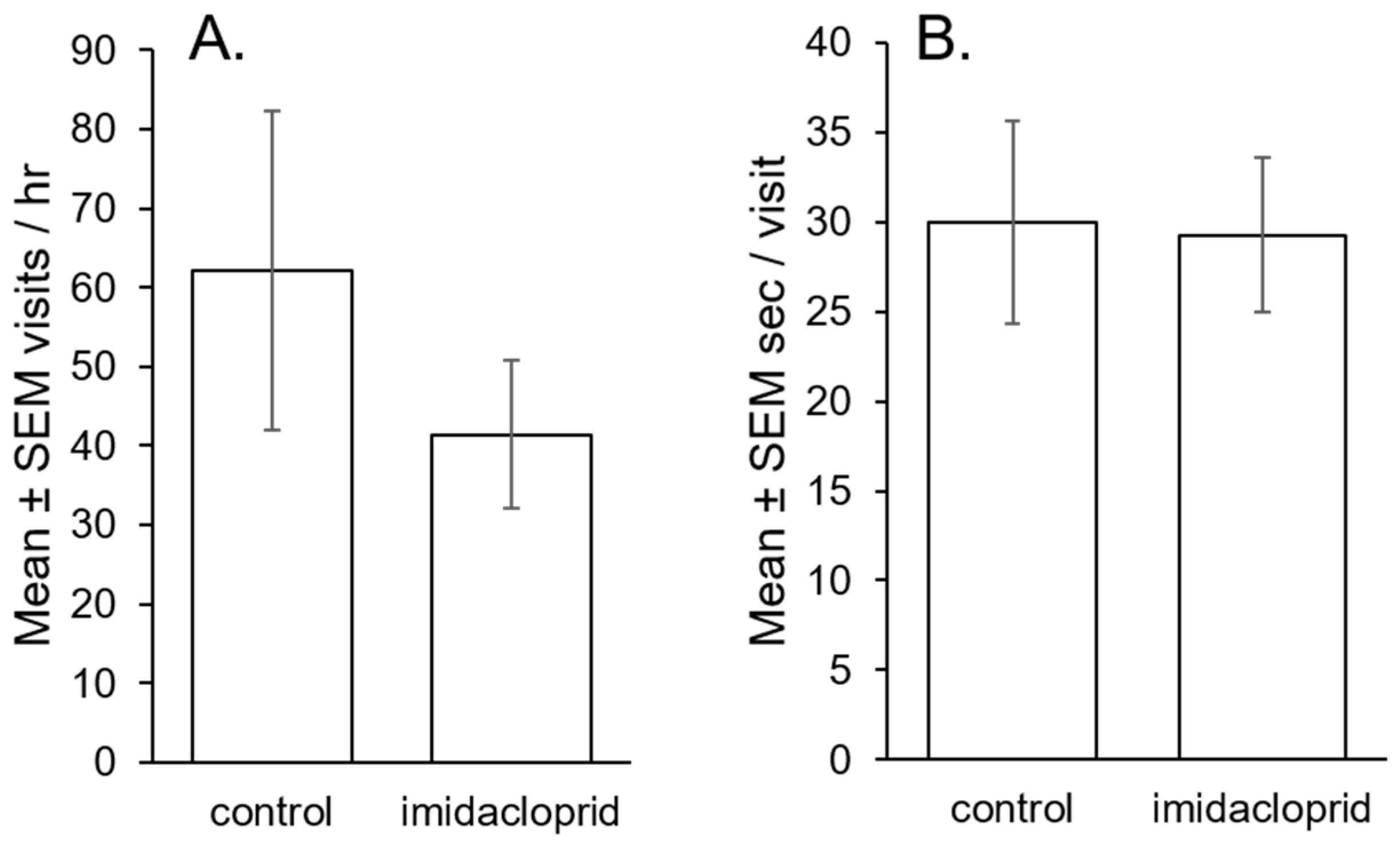
3.2. Bumblebee Potato Flower Visitation
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO International Year of the Potato. Available online: www.fao.org/potato-2008 (accessed on 11 November 2016).
- USDA NASS National Agricultural Statistics Service. Available online: https://www.nass.usda.gov/ (accessed on 12 January 2017).
- Buchmann, S.L.; Cane, J.H. Bees assess pollen returns while sonicating Solanum flowers. Oecologia 1989, 81, 289–294. [Google Scholar] [CrossRef]
- Karlsson, M.F.; Birgersson, G.; Cotes Prado, A.M.; Bosa, F.; Bengtsson, M.; Witzgall, P. Plant odor analysis of potato: Response of Guatemalan Moth to above- and belowground potato volatiles. J. Agric. Food Chem. 2009, 57, 5903–5909. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.W.T. Male-fertile potato flowers are selectively buzz-pollinated only by Bombus terricola Kirby in Upstate New York. J. Kans. Entomol. Soc. 1993, 66, 252–254. [Google Scholar]
- Free, J.B. Insect Pollination of Crops; Academic Press: London, UK, 1993. [Google Scholar]
- White, J.W. Pollination of potatoes under natural conditions. CIP Circ.—Int. Potato Cent. 1983, 11, 1–2. [Google Scholar]
- Marfil, C.F.; Masuelli, R.W. Reproductive ecology and genetic variability in natural populations of the wild potato, Solanum kurtzianum. Plant Biol. 2014, 16, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Celis, C.; Scurrah, M.; Cowgill, S.; Chumbiauca, S.; Green, J.; Franco, J.; Main, G.; Kiezebrink, D.; Visser, R.G.F.; Atkinson, H.J. Environmental biosafety and transgenic potato in a centre of diversity for this crop. Nature 2004, 432, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Koh, I.; Lonsdorf, E.V.; Williams, N.M.; Brittain, C.; Isaacs, R.; Gibbs, J.; Ricketts, T.H. Modeling the status, trends, and impacts of wild bee abundance in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, M.J.; Rey, K.P.; O’Neal, M.E. Defining the insect pollinator community found in Iowa corn and soybean fields: Implications for pollinator conservation. Environ. Entomol. 2016, 45, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Gill, K.A.; O’Neal, M.E. Survey of soybean insect pollinators: Community identification and sampling method analysis. Environ. Entomol. 2015, 44, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, M.J.; O’Neal, M.E. Insect pollinators in Iowa cornfields: Community identification and trapping method analysis. PLoS ONE 2016, 11, e0143479. [Google Scholar] [CrossRef] [PubMed]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Kamel, A. Insecticide residues in pollen and nectar of a cucurbit crop and their potential exposure to pollinators. J. Agric. Food Chem. 2012, 60, 4449–4456. [Google Scholar] [CrossRef] [PubMed]
- Szendrei, Z. Results of the 2010 Michigan potato pest survey. Mich. Potato Newsline 2011, 22, 4–5. [Google Scholar]
- Walters, K.F.A. Neonicotinoids, bees and opportunity costs for conservation. Insect Conserv. Divers. 2016, 9, 375–383. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.A.; Raine, N.E. Chronic exposure to a neonicotinoid pesticide alters the interactions between bumblebees and wild plants. Funct. Ecol. 2016, 30, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.A.; Russell, A.L.; Morrison, S.J.; Rogers, C.; Raine, N.E. Investigating the impacts of field-realistic exposure to a neonicotinoid pesticide on bumblebee foraging, homing ability and colony growth. J. Appl. Ecol. 2016, 53, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Laycock, I.; Lenthall, K.M.; Barratt, A.T.; Cresswell, J.E. Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology 2012, 21, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Pilling, E.; Campbell, P.; Coulson, M.; Ruddle, N.; Tornier, I. A four-year field program investigating long-term effects of repeated exposure of honey bee colonies to flowering crops treated with thiamethoxam. PLoS ONE 2013, 8, e77193. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Plant Protection Products and their Residues (PPR). Scientific opinion on the science behind the development of a risk assessment of Plant Protection Products on bees (Apis mellifera, Bombus spp. and solitary bees): Risk assessment for bees. EFSA J. 2012. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D. A field study examining the effects of exposure to neonicotinoid seed-treated corn on commercial bumble bee colonies. Ecotoxicology 2014, 23, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Cutler, G.C.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L. A large-scale field study examining effects of exposure to clothianidin seed-treated canola on honey bee colony health, development, and overwintering success. PeerJ 2014. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.M.; Wilkins, S.; Harkin, S.; Milner, S.; Walters, K.F. Neonicotinoids and bumblebees (Bombus terrestris): Effects on nectar consumption in individual workers. Pest Manag. Sci. 2015, 71, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Elston, C.; Thompson, H.M.; Walters, K.F.A. Sub-lethal effects of thiamethoxam, a neonicotinoid pesticide, and propiconazole, a DMI fungicide, on colony initiation in bumblebee (Bombus terrestris) micro-colonies. Apidologie 2013, 44, 563–574. [Google Scholar] [CrossRef]
- Leong, J.M.; Thorp, R.W. Colour-coded sampling: The pan trap colour preferences of oligolectic and nonoligolectic bees associated with a vernal pool plant. Ecol. Entomol. 1999, 24, 329–335. [Google Scholar] [CrossRef]
- Stephen, W.P.; Rao, S. Unscented color traps for non-Apis bees (Hymenoptera: Apiformes). J. Kans. Entomol. Soc. 2005, 78, 373–380. [Google Scholar] [CrossRef]
- MSU Enviroweather. Available online: https://mawn.geo.msu.edu/ (accessed on 7 January 2017).
- USDA-NASS CropScape and Cropland Data Layer. Available online: https://www.nass.usda.gov/Research_and_Science/Cropland/SARS1a.php (accessed on 7 February 2017).
- Gibbs, J. Revision of the metallic Lasioglossum (Dialictus) of eastern North America (Hymenoptera: Halictidae: Halictini). Zootaxa 2011, 3073, 1–216. [Google Scholar]
- Gibbs, J.; Packer, L.; Dumesh, S.; Danforth, B.N. Revision and reclassification of Lasioglossum (Evylaeus), L. (Hemihalictus) and L. (Sphecodogastra) in eastern North America (Hymenoptera: Apoidea: Halictidae). Zootaxa 2013, 3672, 1–117. [Google Scholar] [CrossRef]
- LaBerge, W.E. A revision of the bees of the genus Melissodes in North and Central America. Part III (Hymenoptera, Apidae). Univ. Kans. Sci. Bull. 1961, 42, 283–663. [Google Scholar]
- Mitchell, T.B. Bees of the Eastern United States I. Tech. Bull. N. C. Agric. Exp. Stn. 1960, 141, 1–538. [Google Scholar]
- Rehan, S.M.; Sheffield, C.S. Morphological and molecular delineation of a new species in the Ceratina dupla species-group (Hymenoptera: Apidae: Xylocopinae) of eastern North America. Zootaxa 2011, 2873, 35–50. [Google Scholar]
- Mitchell, T.B. Bees of the Eastern United States II. Tech. Bull. N. C. Agric. Exp. Stn. 1962, 152, 1–557. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.E. lme4: Linear Mixed-Effects Models Using Eigen and S4. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 14 November 2016).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. Available online: http://CRAN.R-project.org/package=vegan (accessed on 14 November 2016).
- Roberts, D.A. labdsv: Ordination and Multivariate Analysis for Ecology. Available online: http://CRAN.R-project.org/package=labdsv (accessed on 14 November 2016).
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Anastassiades, M.; Lehotay, S.J.; Stajnbaher, D.; Schenck, F.J. Fast and easy multiresidue method employing acetonitrile extraction/partitioning and “dispersive solid-phase extraction” for the determination of pesticide residues in produce. J. AOAC Int. 2003, 86, 412–431. [Google Scholar] [PubMed]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models. Available online: http://CRAN.R-project.org/package=nlme (accessed on 14 November 2016).
- Champley, S. pwr: Basic Functions for Power Analysis. Available online: http://CRAN.R-project.org/package=pwr (accessed on 14 November 2016).
- Hurd, P.D.J.; Linsley, E.G.; Michelbacher, A.E. Ecology of the Squash and Gourd Bee, Peponapis Pruinosa, on Cultivated Cucurbits in California (Hymenoptera: Apoidea). Smithson. Contrib. Zool. 1974, 168, 1–17. [Google Scholar]
- Giles, V.; Ascher, J.S. A survey of the bees of the Black Rock Forest Preserve, New York (Hymenoptera: Apoidea). J. Hymenopt. Res. 2006, 15, 208–231. [Google Scholar]
- Hurd, P.D. Superfamily Apoidea. In Catalog of Hymenoptera in America North of Mexico; Smithsonian Institution Press: Washington, DC, USA, 1979. [Google Scholar]
- Ashmead, W.H. The habits of the aculeate Hymenoptera IV. Psyche J. Entomol. 1894, 7, 75–79. [Google Scholar] [CrossRef]
- Batra, S.W.T. Behavior of the social bee, Lasioglossum zephyrum, within the nest (Hymenoptera: Halictidæ). Insectes Sociaux 1964, 11, 159–185. [Google Scholar] [CrossRef]
- Breed, M.D. Life cycle and behavior of a primitively social bee, Lasioglossum rohweri (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1975, 48, 64–80. [Google Scholar]
- Coelho, B.W.T. A review of the bee genus Augochlorella (Hymenoptera: Halictidae: Augochlorini). Syst. Entomol. 2004, 29, 282–323. [Google Scholar] [CrossRef]
- Eickwort, G.C. Aspects of the nesting biology of five Nearctic species of Agapostemon (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1981, 54, 337–351. [Google Scholar]
- Eickwort, G.C. First steps into eusociality: The sweat bee Dialictus lineatulus. Fla. Entomol. 1986, 69, 742–754. [Google Scholar] [CrossRef]
- Ivanochko, M. Taxonomy, Biology and Alfalfa Pollinating Potential of Canadian Leaf-Cutter Bees--Genus Megachile Latreille (Hymenoptera: Megachilidae); McGill-Queen’s University Press: Montreal, QC, Canada, 1979. [Google Scholar]
- Krombein, K.V. Trap-Nesting Wasps and Bees: Life Histories, Nests, and Associates; Smithsonian Press: Washington, DC, USA, 1967. [Google Scholar]
- McGinley, R.J. Studies of Halictinae (Apoidea: Halictidae). Smithson. Contrib. Zool. 2003, 429, 1–294. [Google Scholar] [CrossRef]
- Medler, J.T. Anthophora (Clisodon) terminalis Cresson in trap-nests in Wisconsin (Hymenoptera: Anthophoridae). Can. Entomol. 1964, 96, 1332–1336. [Google Scholar] [CrossRef]
- Michener, C.D. The Social Behavior of the Bees: A Comparative Study; Harvard University Press: Cambridge, MA, 1974. [Google Scholar]
- Michener, C.D. The parasitic groups of Halictidae (Hymenoptera, Apoidea). Univ. Kans. Sci. Bull. 1978, 51, 291–339. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2000. [Google Scholar]
- Michener, C.D.; Wille, A. The bionomics of a primitively social bee, Lasioglossum inconspicuum. Univ. Kans. Sci. Bull. 1961, 42, 1123–1202. [Google Scholar]
- Miliczky, E.R. Observations on the nesting biology of Tetralonia hamata Bradley with a description of its mature larva (Hymenoptera: Anthophoridae). J. Kans. Entomol. Soc. 1985, 58, 686–700. [Google Scholar]
- Mueller, U.G. Life history and social evolution of the primitively eusocial bee Augochlorella striata (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1996, 69, 116–138. [Google Scholar]
- Nininger, H.H. Notes on the life-history of Anthophora stanjordiana. Psyche 1920, 27, 135–137. [Google Scholar] [CrossRef]
- Norden, B.B. Nesting biology of Anthophora abrupta (Hymenoptera: Anthophoridae). J. Kans. Entomol. Soc. 1984, 57, 243–262. [Google Scholar]
- Ordway, E. The bionomics of Augochlorella striata and A. persimilis in Eastern Kansas (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1966, 39, 270–313. [Google Scholar]
- Packer, L. Multiple-foundress associations in a temperate population of Halictus ligatus (Hymenoptera; Halictidae). Can. J. Zool. 1986, 64, 2325–2332. [Google Scholar] [CrossRef]
- Packer, L. The social organisation of Lasioglossum (Dialictus) laevissimum (Smith) in southern Alberta. Can. J. Zool. 1992, 70, 1767–1774. [Google Scholar] [CrossRef]
- Packer, L.; Jessome, V.; Lockerbie, C.; Sampson, B. The phenology and social biology of four sweat bees in a marginal environment: Cape Breton Island. Can. J. Zool. 1989, 67, 2871–2877. [Google Scholar] [CrossRef]
- Packer, L.; Sampson, B.; Lockerbie, C.; Jessome, V. Nest architecture and brood mortality in four species of sweat bee (Hymenoptera; Halictidae) from Cape Breton Island. Can. J. Zool. 1989, 67, 2864–2870. [Google Scholar] [CrossRef]
- Ribble, D.W. The monotypic North American subgenus Larandrena of Andrena (Hymenoptera: Apoidea). Bull. Univ. Neb. State Mus. 1967, 6, 27–42. [Google Scholar]
- Richards, M.H.; Packer, L. Annual variation in survival and reproduction of the primitively eusocial sweat bee Halictus ligatus (Hymenoptera: Halictidae). Can. J. Zool. 1995, 73, 933–941. [Google Scholar] [CrossRef]
- Richards, M.H.; Richards, M.H.; Vickruck, J.L.; Rehan, S.M. Colony social organisation of Halictus confusus in southern Ontario, with comments on sociality in the subgenus H. (Seladonia). J. Hymenopt. Res. 2010, 19, 144–158. [Google Scholar]
- Roberts, R.B. Biology of the bee genus Agapostemon (Hymenoptera: Halictidae). Kans. Univ. Sci. Bull. 1969, 48, 101–124. [Google Scholar]
- Roberts, R.B. Revision of the bee genus Agapostemon (Hymenoptera: Halictidae). Kans. Univ. Sci. Bull. 1972, 49, 437–590. [Google Scholar]
- Soucy, S.L. Nesting biology and socially polymorphic behavior of the sweat bee Halictus rubicundus (Hymenoptera: Halictidae). Ann. Entomol. Soc. Am. 2002, 95, 57–65. [Google Scholar] [CrossRef]
- Stockhammer, K.A. Nesting habits and life cycle of a sweat bee, Augochlora pura (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1966, 39, 157–192. [Google Scholar]
- Stockhammer, K.A. Some notes on the biology of the blue sweat bee, Lasioglossum coeruleum (Apoidea: Halictidae). J. Kans. Entomol. Soc. 1967, 40, 177–189. [Google Scholar]
- Vickruck, J.l.; Rehan, S.M.; Sheffield, C.S.; Richards, M.H. Nesting biology and DNA barcode analysis of Ceratina dupla and C. mikmaqi, and comparisons with C. calcarata (Hymenoptera: Apidae: Xylocopinae). Can. Entomol. 2011, 143, 254–262. [Google Scholar] [CrossRef]
- Williams, P.H.; Thorp, R.W.; Richardson, L.L.; Colla, S.R. Bumble Bees of North America: An Identification Guide; Princeton University Press: Princeton, NJ, USA, 2014. [Google Scholar]
- Zayed, A.; Constantin, Ş.A.; Packer, L. Successful biological invasion despite a severe genetic load. PLoS ONE 2007, 2, e868. [Google Scholar] [CrossRef] [PubMed]
- Abrams, J.; Eickwort, G.C. Nest switching and guarding by the communal sweat bee Agapostemon virescens (Hymenoptera, Halictidae). Insectes Sociaux 1981, 28, 105–116. [Google Scholar] [CrossRef]
- Abrams, J.; Eickwort, G.C. Biology of the communal sweat bee Agapostemon virescens (Hymenoptera: Halictidae) in New York State. Agriculture 1980, 1, 1–20. [Google Scholar]
- Graenicher, S. Some observations on the life history and habits of parasitic bees. Bull. Wis. Nat. Hist. Soc. 1905, 3, 153–167. [Google Scholar]
- Pesenko, Y.A.; Banaszak, J.; Radchenko, V.G.; Cierzniak, T. Bees of the Family Halictidae (excluding Sphecodes) of Poland: Taxonomy, Ecology, Bionomics; Wyzszej Szkoly Pedagogicznej: Bydgszsz, Poland, 2000. [Google Scholar]
- Rau, P. Ecological and behavior notes on Missouri insects. Trans. Acad. Sci. St. Louis 1922, 24, 1–71. [Google Scholar]
- Batra, S.W.T. Ethology of the vernal eusocial bee, Dialictus laevissimus (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1987, 60, 100–108. [Google Scholar]
- Atwood, C.E. Studies on the Apoidea of western Nova Scotia with special reference to visitors to apple bloom. Can. J. Res. 1933, 9, 443–457. [Google Scholar] [CrossRef]
- Gibbs, J.; Brady, S.G.; Kanda, K.; Danforth, B.N. Phylogeny of halictine bees supports a shared origin of eusociality for Halictus and Lasioglossum (Apoidea: Anthophila: Halictidae). Mol. Phylogenet. Evol. 2012, 65, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Stout, J.C. Homing ability of the bumblebee Bombus terrestris (Hymenoptera: Apidae). Apidologie 2001, 32, 105–111. [Google Scholar] [CrossRef]
- Sanford, J.C.; Hanneman, R.E. The use of bees for the purpose of inter-mating in potato. Am. Potato J. 1981, 58, 481–485. [Google Scholar] [CrossRef]
- Buchmann, S.L. Buzz pollination in angiosperms. In Handbook of Experimental Pollination Biology; Van Nostrand Reinhold: New York, NY, USA, 1983; pp. 73–113. [Google Scholar]
- Teppner, H. Pollinators of tomato, Solanum lycopersicum (Solanaceae), in Central Europe. Phyton 2005, 45, 217–235. [Google Scholar]
- Bowers, K.A.W. The pollination ecology of Solanum rostratum (Solanaceae). Am. J. Bot. 1975, 62, 633–638. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Mallinger, R.E.; Werts, P.; Gratton, C. Pesticide use within a pollinator-dependent crop has negative effects on the abundance and species richness of sweat bees, Lasioglossum spp., and on bumble bee colony growth. J. Insect Conserv. 2015, 19, 999–1010. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, M.A.; Tuell, J.K.; Isaacs, R.; Gibbs, J.; Ascher, J.S.; Landis, D.A. Implications of three biofuel crops for beneficial arthropods in agricultural landscapes. BioEnergy Res. 2010, 3, 6–19. [Google Scholar] [CrossRef]
- Bennett, A.B.; Isaacs, R. Landscape composition influences pollinators and pollination services in perennial biofuel plantings. Agric. Ecosyst. Environ. 2014, 193, 1–8. [Google Scholar] [CrossRef]
- Adams, D.E.; Perkins, W.E.; Estes, J.R. Pollination systems in Paspalum dilatatum Poir. (Poaceae): An example of insect pollination in a temperate grass. Am. J. Bot. 1981, 68, 389–394. [Google Scholar] [CrossRef]
- Terrell, E.E.; Batra, S.W.T. Insects collect pollen of eastern wildrice, Zizania aquatica (Poaceae). Castanea 1984, 49, 31–34. [Google Scholar]
- Geroff, R.K.; Gibbs, J.; McCravy, K.W. Assessing bee (Hymenoptera: Apoidea) diversity of an Illinois restored tallgrass prairie: Methodology and conservation considerations. J. Insect Conserv. 2014, 18, 951–964. [Google Scholar] [CrossRef]
- Droege, S.; Tepedino, V.J.; LeBuhn, G.; Link, W.; Minckley, R.L.; Chen, Q.; Conrad, C. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Kimoto, C.; DeBano, S.J.; Thorp, R.W.; Rao, S.; Stephen, W.P. Investigating temporal patterns of a native bee community in a remnant North American bunchgrass prairie using blue vane traps. J. Insect Sci. Online 2012. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.K.; Leslie, T.; Rajotte, E.G.; Kammerer, M.A.; Otieno, M.; Biddinger, D.J. Comparative trapping efficiency to characterize bee abundance, diversity, and community composition in apple orchards. Ann. Entomol. Soc. Am. 2015, 108, 785–799. [Google Scholar] [CrossRef]
- Mogren, C.L.; Rand, T.A.; Fausti, S.W.; Lundgren, J.G. The effects of crop intensification on the diversity of native pollinator communities. Environ. Entomol. 2016, 45, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Feltham, H.; Park, K.; Goulson, D. Field realistic doses of pesticide imidacloprid reduce bumblebee pollen foraging efficiency. Ecotoxicology 2014, 23, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Tasei, J.N.; Ripault, G.; Rivault, E. Hazards of imidacloprid seed coating to Bombus terrestris (Hymenoptera: Apidae) when applied to sunflower. J. Econ. Entomol. 2001, 94, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Harder, L.D. Behavioral responses by bumble bees to variation in pollen availability. Oecologia 1990, 85, 41–47. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site ID | Flower Color | Potato Variety | Latitude | Longitude |
|---|---|---|---|---|
| 1 | white | ‘Pike’ | 43.4480 | −84.8882 |
| 2 | white | ‘Pike’ | 43.4444 | −84.8974 |
| 3 | purple | ‘Lamoka’ | 43.4444 | −84.8974 |
| 4 | purple | ‘FL1922’ | 43.3209 | −85.0123 |
| 5 | white | ‘Pike’ | 43.4480 | −84.8882 |
| 6 | purple | ‘FL2137’ | 43.4106 | −84.9183 |
| 7 | pink | ‘Lamoka’ | 43.3867 | −84.9707 |
| 8 | pink | ‘Lamoka’ | 43.3369 | −85.0547 |
| 9 | purple | ‘FL2137’ | 43.3956 | −84.9047 |
| 10 | white | ‘Snowden’ | 43.3828 | −84.9027 |
| 11 | pink | ‘Lamoka’ | 43.3722 | −85.0747 |
| 12 | purple | ‘FL2137’ | 43.3564 | −85.0493 |
| Taxa | Nesting | Behavior | Native/Exotic | References | No. of Individuals | % of Total |
|---|---|---|---|---|---|---|
| ANDRENIDAE | ||||||
| Andrena miserabilis Cresson 1872 | ground | solitary | native | [74] | 1 | 0.1 |
| APIDAE | ||||||
| Anthophora abrupta Say 1837 | ground | solitary | native | [68] | 1 | 0.1 |
| An. bomboides Kirby 1837 | ground | solitary | native | [67] | 2 | 0.3 |
| An. terminalis Cresson 1869 | wood/cavity | solitary | native | [60] | 6 | 0.8 |
| Apis mellifera Linnaeus 1758 | hive | adv. eusocial | exotic | [61] | 24 | 3.2 |
| Bombus auricomus (Robertson 1903) | hive | eusocial | native | [61] | 1 | 0.1 |
| B. bimaculatus Cresson 1863 | hive | eusocial | native | [61] | 7 | 0.9 |
| B. fervidus (Fabricius 1798) | hive | eusocial | native | [61] | 5 | 0.7 |
| B. griseocollis (DeGeer 1773) | hive | eusocial | native | [61] | 2 | 0.3 |
| B. impatiens Cresson 1863 | hive | eusocial | native | [61] | 5 | 0.7 |
| B. perplexus Cresson 1863 | hive | eusocial | native | [61] | 1 | 0.1 |
| B. ternarius Say 1837 | hive | eusocial | native | [61] | 1 | 0.1 |
| Ceratina mikmaqi Rehan and Sheffield 2011 | stem | solitary | native | [82] | 1 | 0.1 |
| Eucera hamata (Bradley 1942) | ground | solitary | native | [65] | 5 | 0.7 |
| Melissodes agilis Cresson 1878 1 | ground | solitary | native | [36,89] | 13 | 1.7 |
| M. bimaculatus (Lepeletier 1825) | ground | solitary | native | [36,51] | 13 | 1.7 |
| M. communis Cresson 1878 2 | ground | solitary | native | [36] | 3 | 0.4 |
| M. desponsus Smith 1854 2,3 | ground | solitary | native | [36] | 11 | 1.5 |
| M. subillatus LaBerge 1961 2,4 | ground | solitary | native | [36] | 2 | 0.3 |
| Peponapis pruinosa (Say 1837) 5 | ground | solitary | native | [48] | 64 | 8.5 |
| COLLETIDAE | ||||||
| Hylaeus affinis (Smith 1853) 2 | stem | solitary | native | 3 | 0.4 | |
| H. mesillae (Cockerell 1896) | stem | solitary | native | [89] | 1 | 0.1 |
| HALICTIDAE | ||||||
| Augochlora pura (Say 1837) | rotten wood | solitary | native | [80] | 4 | 0.5 |
| Augochlorella aurata (Smith 1853) | ground | solitary/eusocial | native | [66,69,72,73] | 2 | 0.3 |
| Agapostemon texanus Cresson 1872 | ground | solitary | native | [55,77] | 2 | 0.3 |
| Ag. virescens (Fabricius 1775) | ground | communal | native | [85,86] | 9 | 1.2 |
| Halictus confusus Smith 1853 | ground | solitary/eusocial | native | [76] | 3 | 0.4 |
| H. ligatus Say 1837 | ground | eusocial | native | [70,75] | 7 | 0.9 |
| H. rubicundus (Christ 1791) | ground | solitary/eusocial | native | [79] | 1 | 0.1 |
| Lasioglossum albipenne (Robertson 1890) 2 | ground | eusocial | native | 2 | 0.3 | |
| L. anomalum (Robertson 1892) 2 | ground | eusocial | native | 1 | 0.1 | |
| L. bruneri (Crawford 1902) 2 | ground | eusocial | native | 13 | 1.7 | |
| L. cinctipes (Provancher 1888) | ground | eusocial | native | [72,73] | 1 | 0.1 |
| L. coeruleum (Robertson 1893) | rotten wood | eusocial | native | [81] | 10 | 1.3 |
| L. coriaceum (Smith 1853) 2 | ground | solitary | native | 12 | 1.6 | |
| L. cressonii (Robertson 1890) 2 | rotten wood | eusocial | native | [39] | 2 | 0.3 |
| L. ellisiae (Sandhouse 1924) 2 | ground | eusocial | native | 3 | 0.4 | |
| L. heterognathum (Mitchell 1960) 2 | ground | eusocial | native | 4 | 0.5 | |
| L. illinoense (Robertson 1892) 2 | ground | eusocial | native | 13 | 1.7 | |
| L. imitatum (Smith 1853) | ground | eusocial | native | [64] | 26 | 3.4 |
| L. laevissimum (Smith 1853) | ground | eusocial | native | [71,72,73] | 2 | 0.3 |
| L. leucocomum (Lovell 1908) 2 | ground | eusocial | native | 48 | 6.4 | |
| L. leucozonium (Schrank 1781) | ground | solitary | exotic | [88,91] | 67 | 8.9 |
| L. lineatulum (Crawford 1906) | ground | eusocial | native | [56] | 36 | 4.8 |
| L. macoupinense (Robertson 1895) 2 | ground | solitary | native | 1 | 0.1 | |
| L. oceanicum (Cockerell 1916) 2 | ground | eusocial | native | 20 | 2.6 | |
| L. paraforbesii McGinley 1986 2 | ground | solitary | native | 14 | 1.9 | |
| L. pectorale (Smith 1853) 2 | ground | solitary | native | 31 | 4.1 | |
| L. perpunctatum (Ellis 1913) 2 | ground | eusocial | native | 48 | 6.4 | |
| L. pilosum (Smith 1853) 2 | ground | eusocial | native | 157 | 20.8 | |
| L. smilacinae (Robertson 1897) 2 | ground | eusocial | native | 4 | 0.5 | |
| L. versatum (Robertson 1902) | ground | eusocial | native | [53] | 11 | 1.5 |
| L. zephyrum (Smith 1853) | ground | eusocial | native | [52] | 1 | 0.1 |
| L. zonulum (Smith 1848) | ground | solitary | exotic | [88,91] | 24 | 3.2 |
| Sphecodes coronus Mitchell 1956 | ground | cleptoparasite | native | [62] | 1 | 0.1 |
| S. cressonii (Robertson 1903) | ground | cleptoparasite | native | [62] | 1 | 0.1 |
| S. mandibularis Cresson 1872 | ground | cleptoparasite | native | [62] | 1 | 0.1 |
| MEGACHILIDAE | ||||||
| Megachile latimanus Say 1823 | ground | solitary | native | [87] | 2 | 0.3 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buchanan, A.L.; Gibbs, J.; Komondy, L.; Szendrei, Z. Bee Community of Commercial Potato Fields in Michigan and Bombus impatiens Visitation to Neonicotinoid-Treated Potato Plants. Insects 2017, 8, 30. https://doi.org/10.3390/insects8010030
Buchanan AL, Gibbs J, Komondy L, Szendrei Z. Bee Community of Commercial Potato Fields in Michigan and Bombus impatiens Visitation to Neonicotinoid-Treated Potato Plants. Insects. 2017; 8(1):30. https://doi.org/10.3390/insects8010030
Chicago/Turabian StyleBuchanan, Amanda L., Jason Gibbs, Lidia Komondy, and Zsofia Szendrei. 2017. "Bee Community of Commercial Potato Fields in Michigan and Bombus impatiens Visitation to Neonicotinoid-Treated Potato Plants" Insects 8, no. 1: 30. https://doi.org/10.3390/insects8010030
APA StyleBuchanan, A. L., Gibbs, J., Komondy, L., & Szendrei, Z. (2017). Bee Community of Commercial Potato Fields in Michigan and Bombus impatiens Visitation to Neonicotinoid-Treated Potato Plants. Insects, 8(1), 30. https://doi.org/10.3390/insects8010030