
Sublethal Effects in Pest Management: A Surrogate Species Perspective on Fruit Fly Control



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Model
2.2. Model Parameterization: Tephritid Life History Data
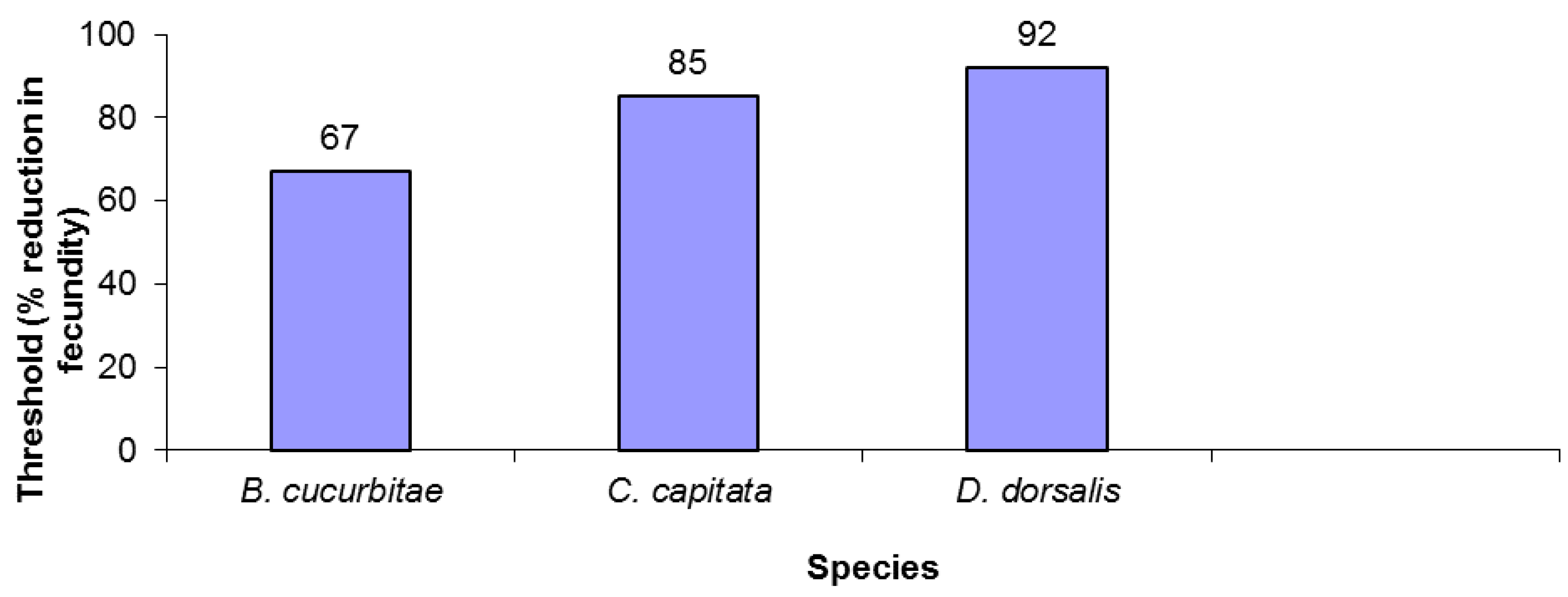
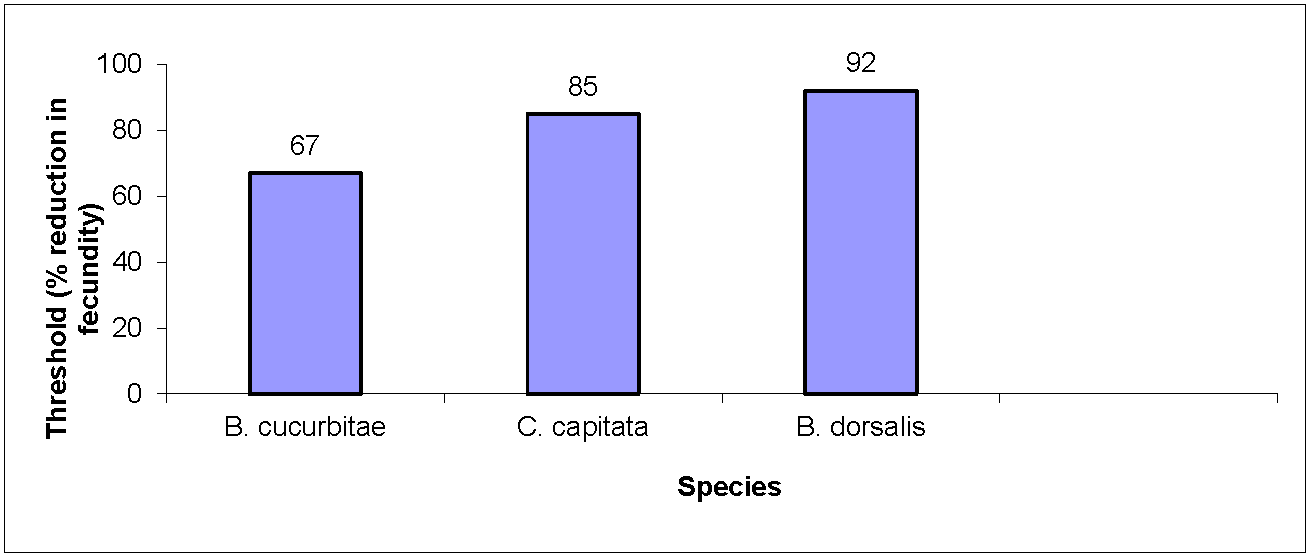
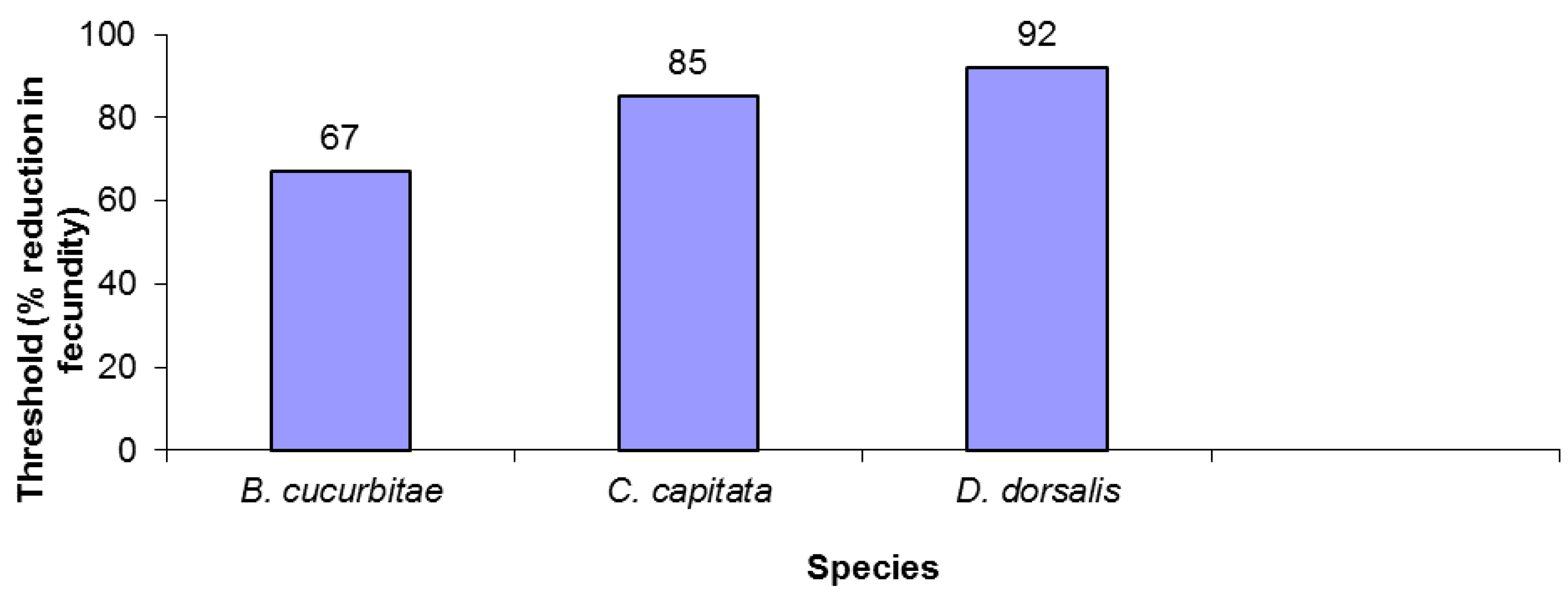
2.3. Calculation of Critical Extinction Thresholds
2.4. Pest Management: Tephritid Species’ Responses to Pesticides
3. Results
4. Discussion and Conclusions
Author Contributions
Conflicts of Interest
References
- Food Quality Protection Act. Available online: https://www.epa.gov/laws-regulations/summary-food-quality-protection-act (accessed on 26 July 2017).
- Banks, J.E.; Stark, J.D.; Vargas, R.I.; Ackleh, A. Parasitoids and ecological risk assessment: Can toxicity data developed for one species be used to protect an entire guild? Biol. Control 2011, 59, 336–339. [Google Scholar] [CrossRef]
- Stark, J.D.; Chen, X.D.; Johnson, C. Effects of herbicides on Behr’s Metalmark butterfly, a surrogate species for the endangered butterfly, Lange’s Metalmark. Environ. Pollut. 2012, 164, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Biondi, A; Zappala`, L.; Stark, J.D.; Desneux, N. Do biopesticides affect the demographic traits of a parasitoid wasp and its biocontrol services through sublethal effects? PLoS ONE 2013, 8, e76548. [Google Scholar] [CrossRef] [PubMed]
- Forbes, V.E.; Calow, P.; Grimm, V.; Hayashi, T.I.; Jager, T.; Katholm, A.; Palmqvist, A.; Pastorok, R.; Salvito, D.; Sibly, R.; Spromberg, J.; Stark, J.D.; Stillman, R.A. Adding value to ecological risk assessment with population modelling. Human Ecol. Risk Asses. 2011, 17, 287–299. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.D.; Banks, J.E.; Vargas, R.I. How risky is risk assessment? The role that life history strategies play in susceptibility of species to stress. Proc. Nat. Acad. Sci. USA 2004, 101, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Hanson, N.; Stark, J.D. A comparison of simple and complex population models to reduce uncertainty in ecological risk assessments of chemicals: Example with three species of Daphnia. Ecotox 2011, 20, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Hanson, N.; Stark, J.D. Utility of population models to reduce uncertainty and increase value relevance in ecological risk assessment: An example with Daphnia exposed to the pesticide spinosad. Int. Environ. Assess. Manag. 2012, 8, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Vargas, R. I.; Miyashita, D.H.; Nishida, T. Life history and demographic parameters of three laboratory reared tephritids (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 1984, 77, 651–656. [Google Scholar] [CrossRef]
- Piñero, J.C.; Mau, R.F.L.; Vargas, R.I. Managing oriental fruit fly (Diptera: Tephritidae) with spinosad-based protein bait sprays and sanitation in papaya orchards in Hawaii. J. Econ. Entomol. 2009, 102, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Prokopy, R.J.; Miller, N.W.; Piñero, J.C.; Barry, J.D.; Tran, L.C.; Oride, L.; Vargas, R.I. Effectiveness of GF-120 fruit fly bait spray applied to border area plants for control of Melon flies (Diptera: Tephritidae). J. Econ. Entomol. 2003, 96, 1485–1493. [Google Scholar] [CrossRef]
- Lefkovitch, L.P. The study of population growth in organisms grouped by stages. Biometrics 1965, 21, 1–18. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis, and Interpretation, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Cushing, J.M. An introduction to structured population dynamics. In CBMS-NSF Regional Conference Series in Applied Mathematics; Society for Industrial and Applied Mathematics (SIAM): Philadelphia, PA, USA, 1998; Volume 71. [Google Scholar]
- Banks, J.E.; Ackleh, A.S.; Stark, J.D. The use of surrogate species in risk assessment: using life history data to safeguard against false negatives. Risk Anal. 2010, 30, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Dyer, S.D.; Versteeg, D.J.; Belanger, S.E.; Chaney, J.G.; Raimondo, S.S.; Barron, M.G. Comparison of species sensitivity distributions derived from interspecies correlation models to distributions used to derive water quality criteria. Environ. Sci. Technol. 2008, 42, 3076–3083. [Google Scholar] [CrossRef]
- Liquido, N.J.; McQuate, G. T.; Suiter, K. CoFFHI: Compendium of Fruit Fly Host Information, version 1.0. United States Department of Agriculture, Center for Plant Health Science and Technology. Available online: https://www.cabdirect.org/cabdirect/abstract/20173115077 (accessed on 26 July 2017).
- Vreysen, M.J.B.; Robinson, A.S.; Hendrichs, J.; Kenmore, P. Areawide integrated pest management (AW-IPM): principles, practice and prospects. In Area-wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 3–33. [Google Scholar]
- Mau, R.F.L.; Jang, E.B.; Vargas, R.I. The Hawaii area-wide fruit fly pest management programme: Influence of partnerships and a good education programme. In Area-wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands; pp. 671–683.
- Stark, J.D.; Vargas, R.; Miller, N. Toxicity of spinosad in protein bait to three economically important tephritid fruit fly species (Diptera: Tephritidae) and their parasitoids (Hymenoptera: Braconidae). J. Econ. Entomol. 2004, 97, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, C. P.; Sherman, M. Lethal and sublethal effects of avermectin B1 on three fruit fly species (Diptera: Tephritidae). J. Econ. Entomol. 1987, 80, 344–347. [Google Scholar] [CrossRef]
- Stark, J.D.; Vargas, R.I.; Messing, R.H.; Purcell, M. Effects of cyromazine and diazinon on three economically important Hawaiian tephritid fruit flies (Diptera: Tephritidae) and their endoparasitoids (Hymenoptera: Braconidae). J. Econ. Entomol. 1992, 85, 1687–1694. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species | C. capitata | B. cucurbitae | B. dorsalis | |||
|---|---|---|---|---|---|---|
| Stage | ||||||
| Egg | 0.96 | 0 | 0.86 | 0 | 0.92 | 0 |
| Larva | 0.916667 | 0 | 0.918605 | 0 | 0.771739 | 0 |
| Pupa | 0.943182 | 0 | 0.911392 | 0 | 0.859155 | 0 |
| Adult | 0.012048 | 8.434882 | 0.013889 | 4.232877 | 0.016393 | 22.13273 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banks, J.E.; Vargas, R.I.; Ackleh, A.S.; Stark, J.D. Sublethal Effects in Pest Management: A Surrogate Species Perspective on Fruit Fly Control. Insects 2017, 8, 78. https://doi.org/10.3390/insects8030078
Banks JE, Vargas RI, Ackleh AS, Stark JD. Sublethal Effects in Pest Management: A Surrogate Species Perspective on Fruit Fly Control. Insects. 2017; 8(3):78. https://doi.org/10.3390/insects8030078
Chicago/Turabian StyleBanks, John E., Roger I. Vargas, Azmy S. Ackleh, and John D. Stark. 2017. "Sublethal Effects in Pest Management: A Surrogate Species Perspective on Fruit Fly Control" Insects 8, no. 3: 78. https://doi.org/10.3390/insects8030078
APA StyleBanks, J. E., Vargas, R. I., Ackleh, A. S., & Stark, J. D. (2017). Sublethal Effects in Pest Management: A Surrogate Species Perspective on Fruit Fly Control. Insects, 8(3), 78. https://doi.org/10.3390/insects8030078