
Oviposition, Life Cycle, and Longevity of the Leaf-Cutting Ant Acromyrmex rugosus rugosus


Abstract
:1. Introduction
2. Material and Methods
2.1. Colony Collection
2.2. Storage of the Colonies in the Laboratory
2.3. Queen Oviposition
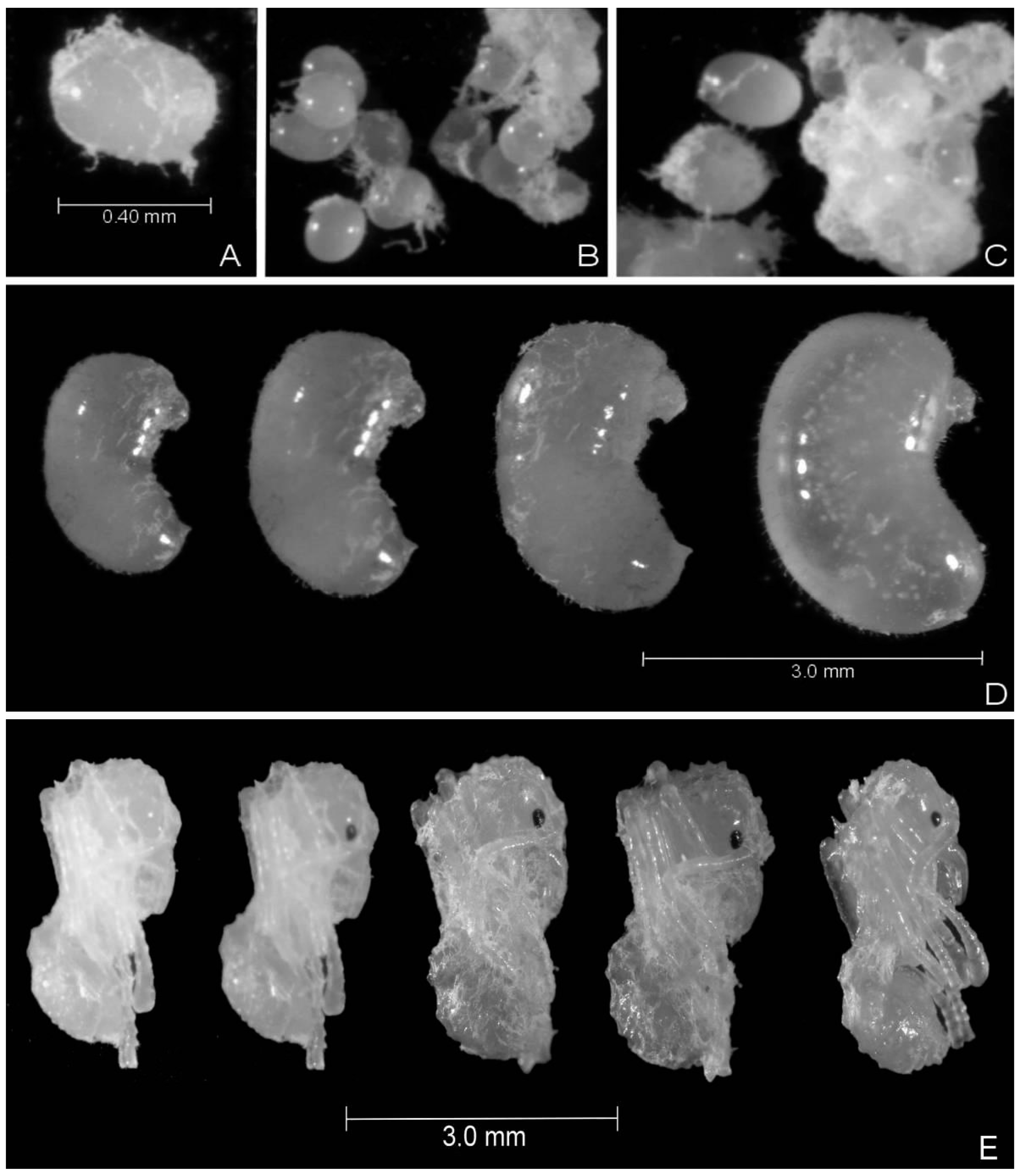
2.4. Workers’ Life Cycle (Egg Period until the Workers’ Emergence)
2.5. Workers’ Longevity
2.6. Data Analysis
3. Results
3.1. Queen Oviposition
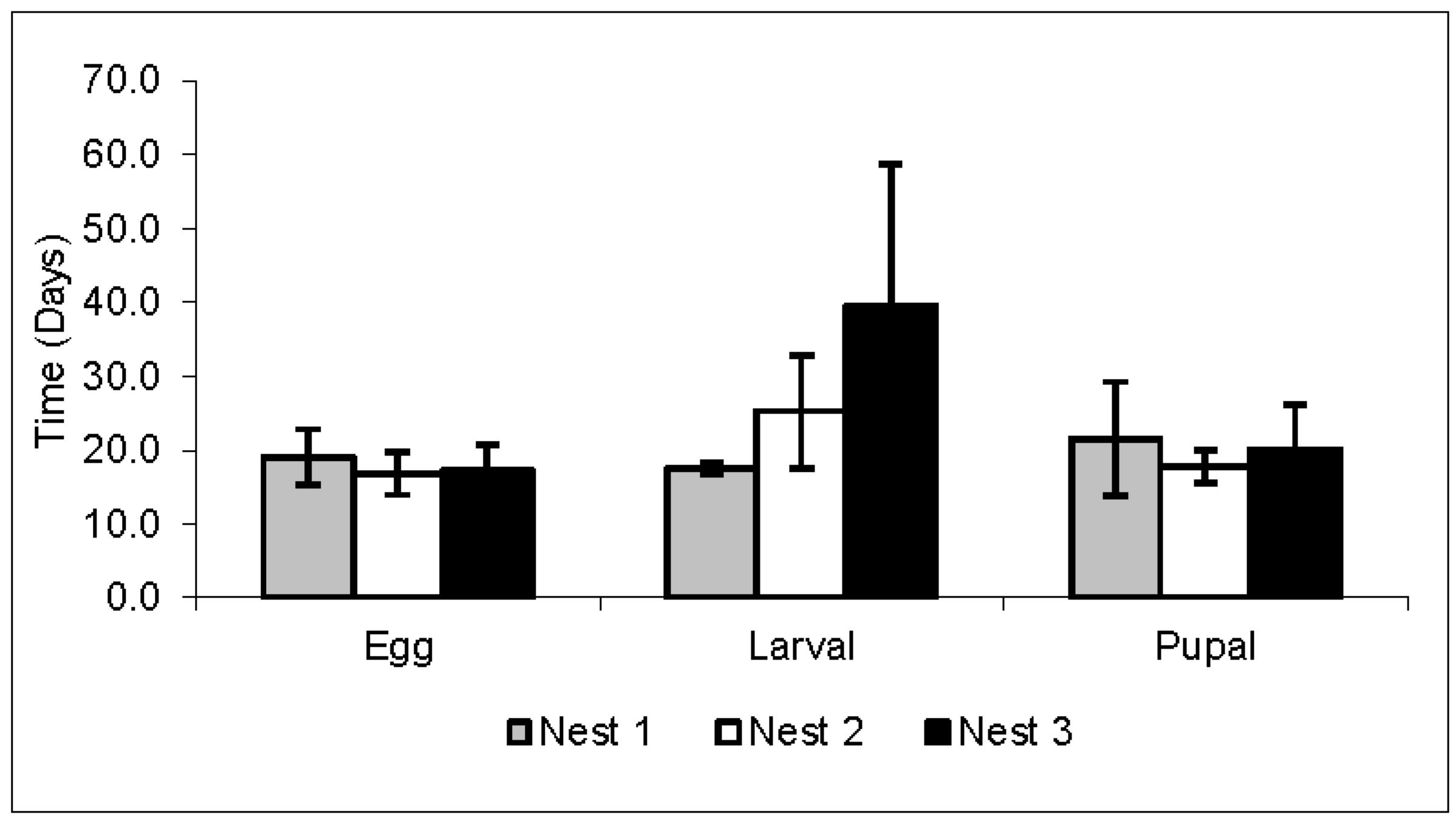
3.2. Workers’ Life Cycle
3.3. Longevity
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990; p. 732. [Google Scholar]
- Marinho, C.G.S.; Della Lucia, T.M.C. Egg-laying in Acromyrmex spp. (Hymenoptera: Formicidae) under laboratory conditions. Biociências 1998, 6, 71–79. [Google Scholar]
- Passera, L. Overview. In Applied Myrmecology: A World Perspective; Vander Meer, R.K., Jaffé, K., Cedano, A., Eds.; Westview Press: Boulder, CO, USA, 1990; pp. 133–137. [Google Scholar]
- Wilson, E.O. The Insect Societies; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1971; p. 548. [Google Scholar]
- Autuori, M. Algumas observações sobre formigas cultivadoras de fungo (Hym. Formicidae). Rev. Entomol. 1940, 11, 215–226. [Google Scholar]
- Autuori, M. Contribuição para o conhecimento da saúva (Atta spp.—Hymenoptera: Formicidae). II—O sauveiro inicial (Atta sexdens rubropilosa Forel, 1908). Arq. Inst. Biol. 1942, 13, 67–86. (In Portuguese) [Google Scholar]
- Bazire-Benazet, M. Sur la formation de l’oeuf alimentaire chez Atta sexdens rubropilosa, Forel, 1908 (Hym., Formicidae). Comptes Rendus de l’Académie des Sciences 1957, 244, 1277–1280. (In French) [Google Scholar]
- Weber, N.A. Gardening Ants: The Attines; Memoirs of the American Philosophical Society: Philadelphia, PA, USA, 1972; p. 146. [Google Scholar]
- Fowler, H.G. Variation sexual diferencial en Acromyrmex rugosus rugosus (Hymenoptera-Formicidae-Attini). Neotropica 1978, 24, 141–144. [Google Scholar]
- Febvay, G.; Ogier, C. Observations sur une ponte fertile d’ouvrieres d’Acromyrmex octospinosus (Reich) (Hymenoptera—Formicidae). Turrialba 1984, 34, 107–108. (In French) [Google Scholar]
- Camargo, R.S.; Forti, L.C.; Andrade, A.P.P.; Matos, C.A.O.; Lopes, J.F.S. Morphometry of the sexuals forms of Acromyrmex subterraneus brunneus Forel, 1911 (Hym., Formicidae) in queenright and queenless laboratory colonies. J. Appl. Entomol. 2005, 129, 347–351. [Google Scholar] [CrossRef]
- Dijkstra, M.B.; Boomsma, J.J. Are workers of Atta leafcutter ants capable of reproduction? Insectes Soc. 2006, 53, 136–140. [Google Scholar] [CrossRef]
- Bazire-Benazet, M. La ponte des ouvrières d’Atta laevigata Fred. Smith 1858 (Hym. Form.). Comptes Rendus de l’Académie des Sciences 1970, 270, 1614–1615. (In French) [Google Scholar]
- Dijkstra, M.B.; Nash, D.R.; Boomsma, J.J. Self-restraint and sterility in workers of Acromyrmex and Atta leafcutter ants. Insectes Soc. 2005, 52, 67–76. [Google Scholar] [CrossRef]
- Weber, N.A. Fungus-growing ants. Science 1966, 153, 587–604. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, J.; Markin, J.P. Brood nutrition and parental relationships of the imported fire ant Solenopsis invicta. J. Geogr. Entomol. Soc. 1973, 8, 294–303. [Google Scholar]
- Schreiber, J.R. Isolation of Attini brood from the social environment (Hymenoptera: Formicidae). Entomol. News 1974, 85, 303–314. [Google Scholar]
- Sudd, J.H. Ants: foraging, nesting, brood behavior, and polyethism. In Social Insect, 4th ed.; Hermann, H.R., Ed.; Academic Press: New York, NY, USA, 1982; pp. 107–155. [Google Scholar]
- Cassill, D.L.; Tschinkel, W.R. Effects of colony level attributes on larval feeding in the fire ant, Solenopsis invicta. Insectes Soc. 1999, 46, 261–266. [Google Scholar] [CrossRef]
- Schneider, M.O. Comportamento de Cuidado Com A Prole da Saúva-Limão Atta sexdEns Rubropilosa Forel, 1908 (Hymenoptera: Formicidae). [Dissertação (Mestrado em Ciências Biológicas/Zoologia)]. Ph.D. Thesis, Instituto de Biociências, Universidade Estadual Paulista, Rio Claro, Brazil, 2003; p. 80. (In Portuguese). [Google Scholar]
- Lopes, J.F.S.; Hughes, W.H.O.; Camargo, R.S.; Forti, L.C. Larval isolation and brood care in Acromyrmex leaf-cutting ants. Insectes Soc. 2005, 52, 333–338. [Google Scholar] [CrossRef]
- Camargo, R.S.; Forti, L.C.; Lopes, J.F.S.; Andrade, A.P.P.; Ottati, A.L.T. Age polyethism in the leaf-cutting ant Acromyrmex subterraneus brunneus Forel, 1911 (Hymenoptera: Formicidae). J. Appl. Entomol. 2007, 131, 139–145. [Google Scholar] [CrossRef]
- Bourke, A.F.G.; Franks, N.R. Social Evolution in Ants; Princeton University Press: Princeton, NJ, USA, 1995; p. 550. [Google Scholar]
- Coutinho, L.M. Aspectos ecológicos da saúva no cerrado. Os murundus de terra, as características psamofiticas da espécie e sua vegetação, e sua invasão pelo capim gordura. Rev. Bras. Biol. 1982, 42, 147–153. (In Portuguese) [Google Scholar]
- Silva, A.; Bacci, M.; Siqueira, C.G.; Bueno, O.C.; Pagnocca, F.C.; Hebling, M.J.A. Survival of Atta sexdens workers on different food sources. J. Insect Physiol. 2003, 49, 307–313. [Google Scholar] [CrossRef]
- Mueller, U.G.; Schultz, T.R.; Currie, C.R.; Adams, R.M.; Malloch, D. The origem of the Attine ant-fungus mutualism. Q. Rev. Biol. 2001, 76, 169–197. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. Sociobiology: The New Synthesis; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1975; p. 697. [Google Scholar]
- Andrade, A.P.P. Biologia E Taxonomia Comparadas Das Subespécies de Acromyrmex subterraneus Forel, 1893 (Hymenoptera, Formicidae) E Contaminação Das Operárias Por Iscas Tóxicas. [Tese (Doutorado em Ciências Biológicas/Zoologia)]. Ph.D. Thesis, Instituto de Biociências, Universidade Estadual Paulista, Botucatu, Brazil, 2002; p. 168. (In Portuguese). [Google Scholar]
- SAS System, version 8.2; SAS Institute: Cary, NC, USA, 2001.
- Weber, N.A. A 10-year colony of Acromyrmex octospinosus (Hymenoptera: Formicidae). Proc. Entomol. Soc. Wash. 1977, 79, 284–292. [Google Scholar]
- Bruner, S.C.; Barry, F.V. Observaciones sobre la biologia de la bibijagua. Mem. Soc. Cub. Hist. Nat. 1949, 19, 135–154. [Google Scholar]
- Mintzer, A.; Vinson, S.B. Cooperative colony foundation by females of the leafcutting ant Atta texana in the laboratory. J. N. Y. Entomol. Soc. 1985, 93, 1047–1051. [Google Scholar]
- Della Lucia, T.M.C.; Vilela, E.F.; Moreira, D.D.O.; Bento, J.M.S.; Dos Anjos, N. Egg-laying in Atta sexdens rubropilosa, under laboratory conditions. In Applied Myrmecology: A World Perspective; Vander Meer, R.K., Jaffé, K., Cedeño, A., Eds.; Westview Press: Boulder, CO, USA, 1990; pp. 173–179. [Google Scholar]
- Araújo, M.S.; Della Lucia, T.M.C. Periodicidade de oviposição em rainhas de Atta laevigata F. Smith, 1858 (Hymenoptera: Formicidae) em condições de laboratório. Rev. Ceres 1993, 40, 104–112. (In Portuguese) [Google Scholar]
- Ribeiro, M.M.R.; Marinho, C.G.S.; Della Lucia, T.M.C. Oviposição em Atta bisphaerica (Hymenoptera, Formicidae) sob condições de laboratório. In Mirmecologia-Biodiversidade E Bioindicação; Fernandes, W.D., Silvestre, R., Aranda, R., Eds.; Universidade Federal de Mato Grosso do Sul: Campo Grande, MS, Brazil, 2005; pp. 417–418. [Google Scholar]
- LaPolla, J.S.; Mueller, U.G.; Seid, M.; Cover, S.P. Predation by the army ant Neivamyrmex rugulosus on the fungus-growing ant Trachymyrmex arizonensis. Insectes Soc. 2002, 49, 251–260. [Google Scholar] [CrossRef]
- Camargo, R.S.; Forti, L.C.; Lopes, J.F.S.; Noronha, N.C.J.; Ottati, A.L.T. Laying by Acromyrmex subterraneus brunneus Forel, 1911 (Hymenoptera: Formicidae) workers in queenless colonies. Insectes Sci. 2007, 14, 65–75. [Google Scholar]
- Passera, L.; Aron, S. Early sex discrimination and male brood elimination by workers of the Argentine ant. Proc. R. Soc. Lond. B 1996, 263, 1041–1046. [Google Scholar] [CrossRef]
- Bollazzi, M.; Roces, F. Thermal preference for fungus culturing and brood location by workers of the thatching grass-cutting ant Acromyrmex heyeri. Insectes Soc. 2002, 49, 153–157. [Google Scholar] [CrossRef]
- Wheeler, D.E. Nourishment in ants: Patterns in individuals and societies. Nourishment and Evolution in Insect Societies; Hunt, J.H., Napela, C.A., Eds.; Westview Press: Boulder, CO, USA, 1994; pp. 245–278. [Google Scholar]
- Torre-Grossa, J.P.; Febvay, G.; Kermarrec, A. Larval instars of the worker caste in the Attine ant, Acromyrmex octospinosus (Hymenoptera; Formicidae). Colemania 1982, 1, 141–147. [Google Scholar]
- Solis, D.R.; Fox, E.G.P.; Ceccato, M.; Reiss, I.C.; Décio, P.; Lorenzon, N.; Silva, N.G.; Bueno, O.C. On the morphology of the worker immatures of the leafcutter ant Atta sexdens Linnaeus (Hymenoptera: Formicidae). Microsc. Res. Tech. 2012, 75, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.D.; Tschinkel, W.R. Fire and polymorphism: The ergonomics of brood production. Behav. Ecol. Sociobiol. 1985, 16, 323–336. [Google Scholar] [CrossRef]
- Chapuisat, M.; Keller, L. Division of labour influences the rate of ageing in weaver ant workers. Proc. R. Soc. B 2002, 269, 909–913. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nests | Total | Average Rate/Day | SD | Eggs/h |
|---|---|---|---|---|
| 1 | 338 | 84.5 | 53.6 | 3.5 |
| 2 | 231 | 57.8 | 45.7 | 2.4 |
| 3 | 151 | 37.8 | 22.7 | 1.6 |
| Species | Development Stages/Days | Emergence/Days | References | ||
|---|---|---|---|---|---|
| Egg | Larval | Pupal | Worker | ||
| Acromyrmex rugosus rugosus | 17.7 | 27.4 | 19.8 | 62.9 | |
| Acromyrmex octospinosus | 24.0 | 18.0 | 16.0 | 63.0 | [30] |
| Acromyrmex lundi | - | - | - | 60.0 | [8] |
| Acromyrmex subterraneus subterraneus | 21.8 | 21.8 | 15.8 | 62;0 | [28] |
| Acromyrmex subterraneus brunneus | 21.2 | 25.2 | 21.2 | 69.4 | [28] |
| Acromyrmex subterraneus molestans | 20.8 | 17.2 | 14.2 | 53.0 | [28] |
| Atta sexdens rubropilosa | 25.0 | 22.0 | 10.0 | 62.0 | [6] |
| Atta insularis | 15.5 | 17.0 | 14.0 | 50.0 | [31] |
| Atta texana | 15.0 | - | 14.0 | 50.0 | [32] |
| Nest | Number of Workers | Longevity | |
|---|---|---|---|
| Maximum (Weeks/Months) | Median (Weeks) | ||
| 1 | 18 | 12/3 | 5 |
| 2 | 27 | 25/6 | 5 |
| 3 | 23 | 29/7 | 7 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verza, S.S.; Mussury, R.M.; Camargo, R.S.; Andrade, A.P.P.; Forti, L.C. Oviposition, Life Cycle, and Longevity of the Leaf-Cutting Ant Acromyrmex rugosus rugosus. Insects 2017, 8, 80. https://doi.org/10.3390/insects8030080
Verza SS, Mussury RM, Camargo RS, Andrade APP, Forti LC. Oviposition, Life Cycle, and Longevity of the Leaf-Cutting Ant Acromyrmex rugosus rugosus. Insects. 2017; 8(3):80. https://doi.org/10.3390/insects8030080
Chicago/Turabian StyleVerza, Sandra S., Rosilda M. Mussury, Roberto S. Camargo, Ana Paula P. Andrade, and Luiz C. Forti. 2017. "Oviposition, Life Cycle, and Longevity of the Leaf-Cutting Ant Acromyrmex rugosus rugosus" Insects 8, no. 3: 80. https://doi.org/10.3390/insects8030080
APA StyleVerza, S. S., Mussury, R. M., Camargo, R. S., Andrade, A. P. P., & Forti, L. C. (2017). Oviposition, Life Cycle, and Longevity of the Leaf-Cutting Ant Acromyrmex rugosus rugosus. Insects, 8(3), 80. https://doi.org/10.3390/insects8030080