
Reduced Drosophila suzukii Infestation in Berries Using Deterrent Compounds and Laminate Polymer Flakes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Colony
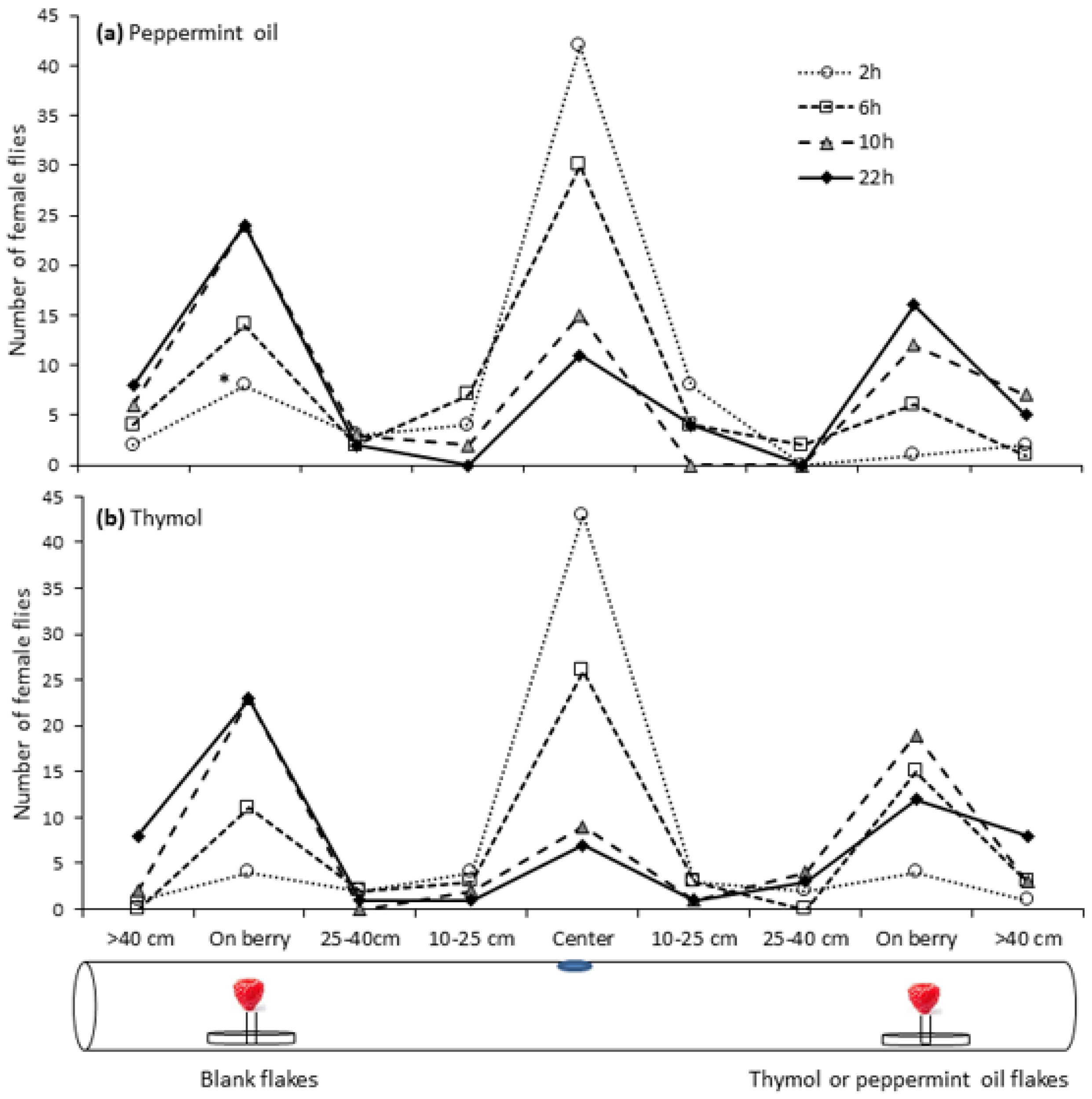
2.2. Repellency of Essential Oil Compounds Using Choice Bioassays
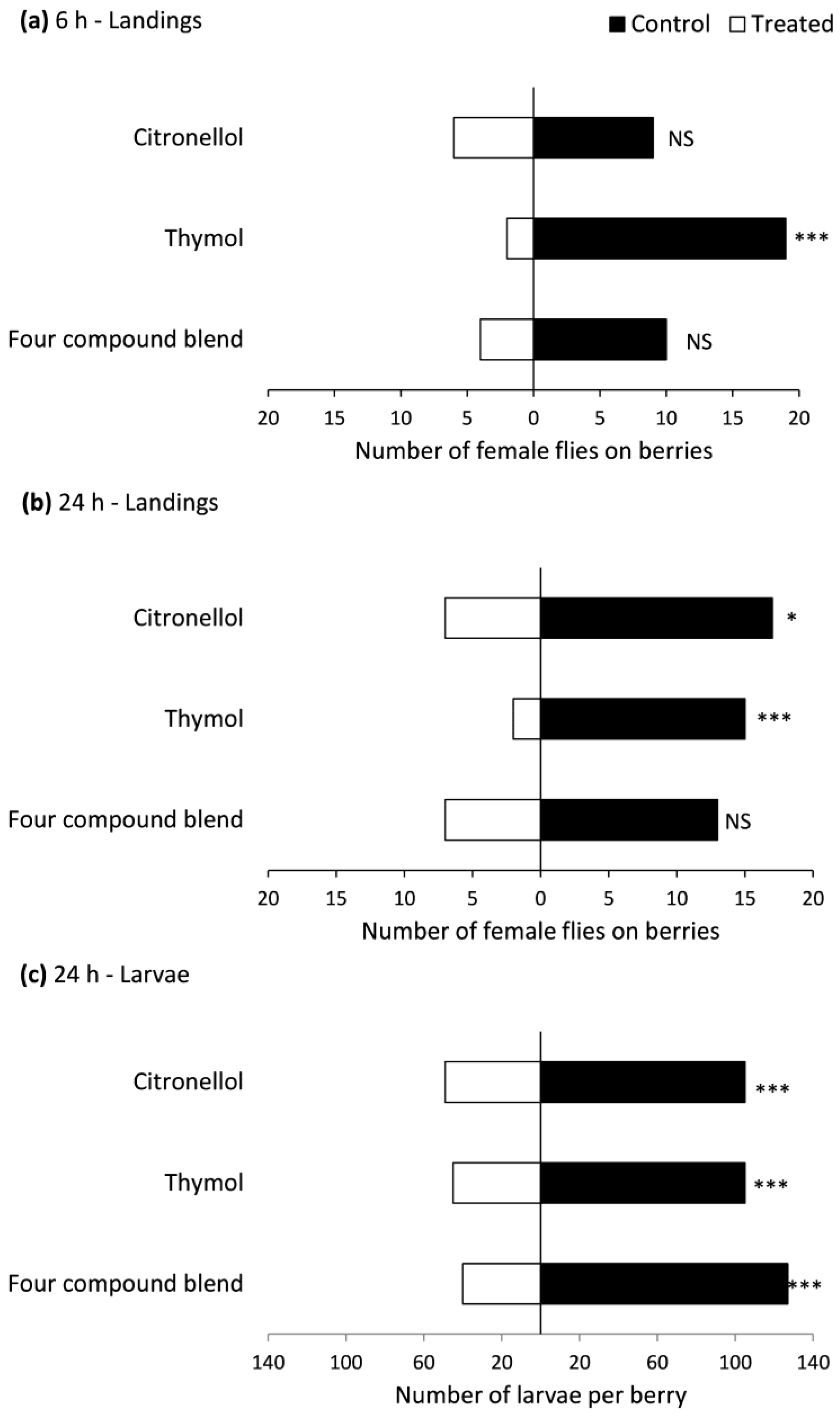
2.3. Oviposition Deterrence of Repellent Compounds Using Choice and No-Choice Bioassays
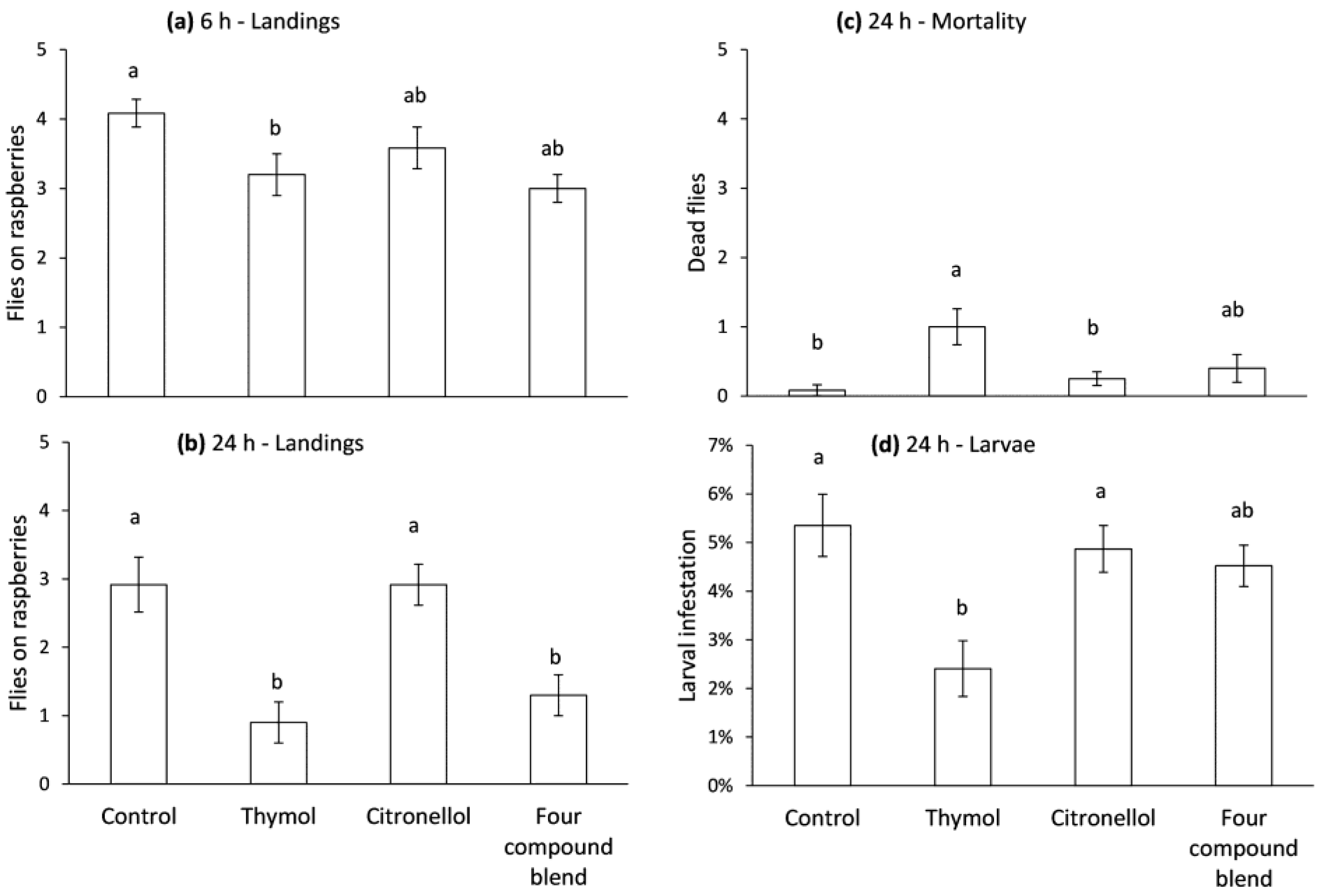
2.4. Repellency and Oviposition Deterrence of Compounds in Biopolymer Flakes Using a Choice Bioassay
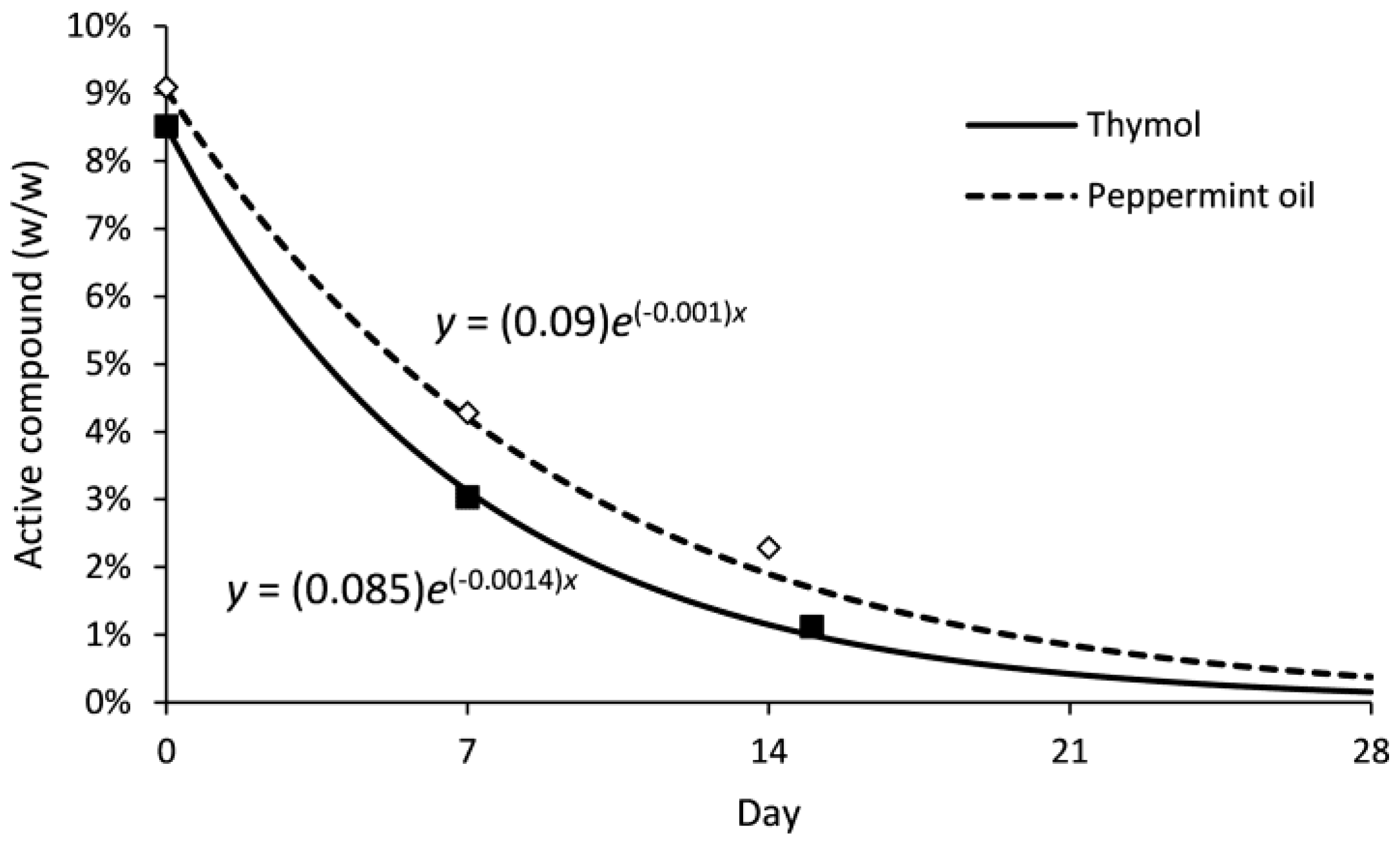
2.5. Oviposition Deterrence of Compounds in Biopolymer Flakes in the Field
2.6. Data Analysis
3. Results
3.1. Repellency of Essential Oil Compounds Using a Choice Bioassay
3.2. Oviposition Deterrence of Repellent Compounds Using Choice and No-Choice Bioassays
3.3. Repellency and Oviposition Deterrence of Compounds in Biopolymer Flakes Using a Choice Bioassay
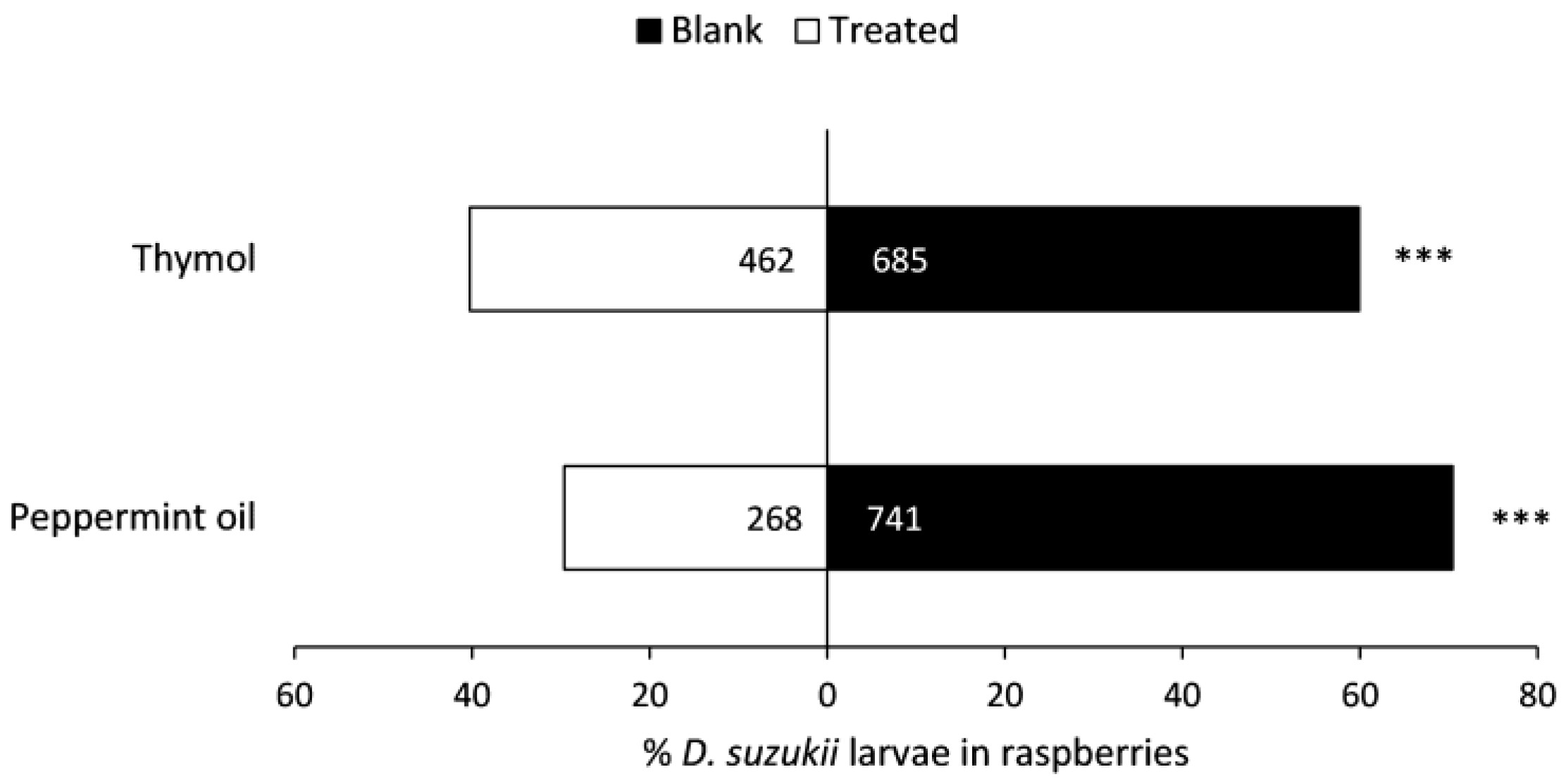
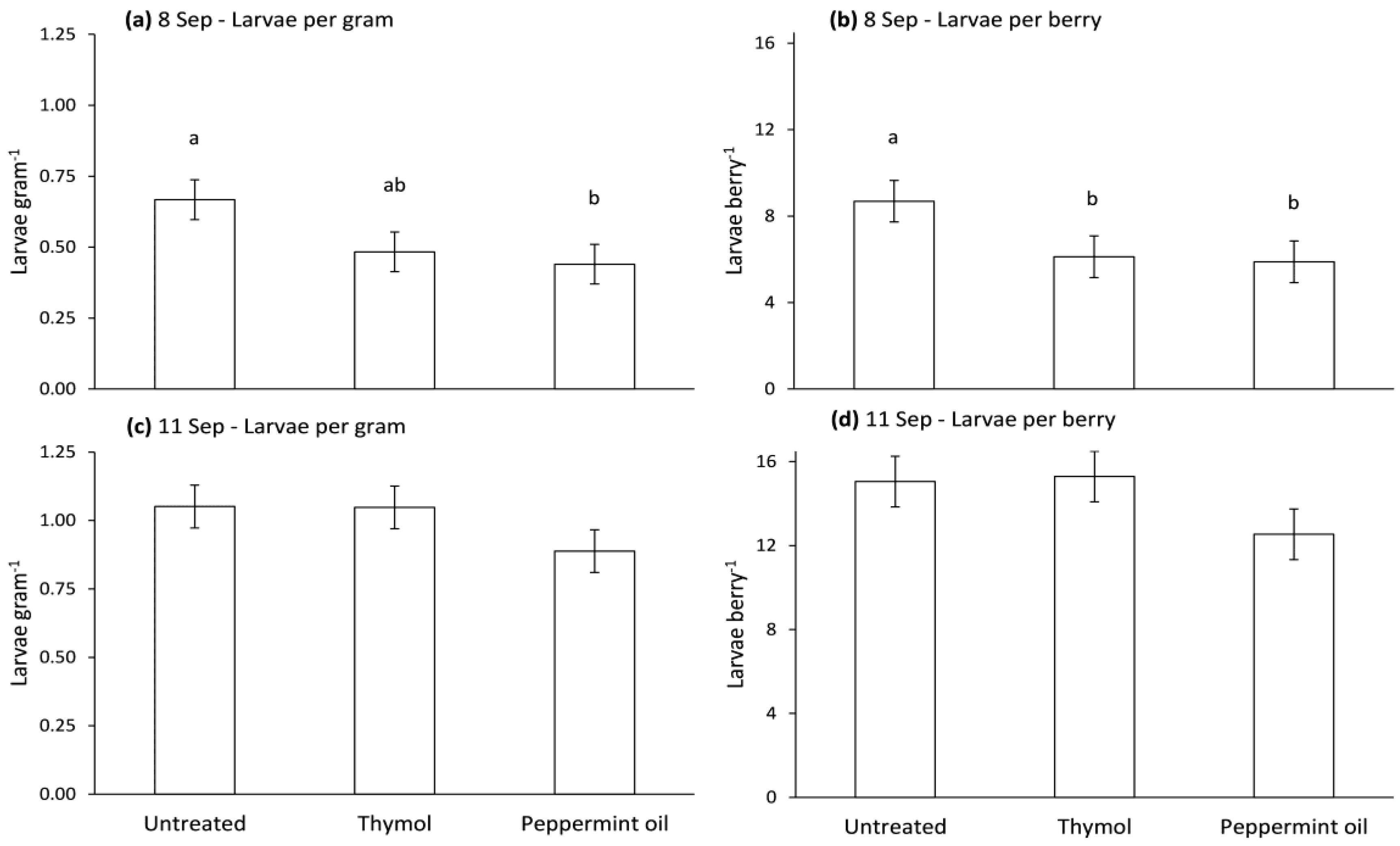
3.4. Oviposition Deterrence of Compounds in Biopolymer Flakes in the Field
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion of biology of spotted wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2014, 88, 469–494. [Google Scholar] [CrossRef]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Cini, A.; Loriatti, C.; Anfora, G. A review of the invasion of Drosphila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.C.; Valente, V.L.S. The first records of the invasive pest Drosophila suzukii in the South American continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wiman, N.G.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Burrack, H.J.; Chiu, J.; Daane, K.M.; Grassi, A.; Miller, B.; Tochen, S.; et al. Integrating temperature-dependent life table data into a matrix projection model for Drosophila suzukii population estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation in California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufield, P. In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Hamby, K.A.; Kwok, R.S.; Zalom, F.G.; Chiu, J.C. Integrating circadian activity and gene expression profiles to predict chronotoxicity of Drosophila suzukii response to insecticides. PLoS ONE 2013, 8, e68472. [Google Scholar] [CrossRef]
- Cowles, R.S.; Saona-Rodriguez, C.; Holdcraft, R.; Loeb, G.M.; Elsensohn, J.E.; Hesler, S.P. Sucrose improves insecticide activity against Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2015, 108, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2015, 140, 37–46. [Google Scholar] [CrossRef]
- Cormier, D.; Veilleux, J.; Firlej, A. Exclusion net to control spotted wing drosophila in blueberry fields. IOBC WPRS Bull. 2015, 109, 181–184. [Google Scholar]
- Leach, H.; Van Timmeren, S.; Isaacs, R. Exclusion netting delays and reduces Drosophila suzukii (Diptera: Drosophilidae) infestation in raspberries. J. Econ. Entomol. 2016, 109, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Schetelig, M.F.; Handler, A.M. Germline transformation of the spotted wing drosophilid, Drosophila suzukii, with a piggyBac transposon vector. Genetica 2013, 141, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Gabarra, R.; Riudavets, J.; Rodríguez, G.A.; Pujade-Villar, J.; Arnó, J. Prospects of the biological control of Drosophila suzukii. BioControl 2015, 60, 331–339. [Google Scholar] [CrossRef]
- Renkema, J.M.; Telfer, Z.; Gariepy, T.; Hallett, R.H. Dalotia coriaria as a predator of Drosophila suzukii: Functional responses, reduced fruit infestation and molecular diagnostics. Biol. Control 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Audsley, N. Further screening of entomopathogenic fungi and nematodes as control agents for Drosophila suzukii. Insects 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Taning, C.N.T.; Christiaens, O.; Berkvens, N.; Casteels, H.; Maes, M.; Smagghe, G. Oral RNAi to control Drosohila suzukii: Laboratory testing against larval and adult stages. J. Pest Sci. 2016, 89, 803–814. [Google Scholar] [CrossRef]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Hofstetter, R.W.; Biddick, L.L. Plant essential oils as arrestants and repellents for neonate larvae of the codling moth (Lepidoptera: Tortricidae). Environ. Entomol. 1999, 28, 954–960. [Google Scholar] [CrossRef]
- Van Tol, R.W.H.M.; Swarts, H.J.; van der Linden, A.; Visser, J.H. Repellence of the red bud borer Resseliella oculiperda from grafted apple trees by impregnation of rubber budding strips with essential oils. Pest Manag. Sci. 2007, 63, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Picard, I.; Hollingsworth, R.G.; Salmieri, S.; Lacroix, M. Repellency of essential oils to Frankliniella occidentalis (Thysanoptera: Thripidae) as affected by type of oil and polymer release. J. Econ. Entomol. 2012, 105, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Wallingford, A.K.; Hesler, S.P.; Cha, D.H.; Loeb, G.M. Behavioral response of spotted-wing drosophila, Drosophila suzukii Matsumura, to aversive odors and a potential oviposition deterrent in the field. Pest Manag. Sci. 2015, 72, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Krause-Pham, C.; Ray, A. Conservation of olfactory avoidance in Drosophila species and identification of repellents of Drosophila suzukii. Sci. Rep. 2015, 5, 11527. [Google Scholar] [CrossRef] [PubMed]
- Renkema, J.M.; Wright, D.; Buitenhuis, R.; Hallett, R.H. Plant essential oils and potassium metabisulfite as repellents for Drosophila suzukii (Diptera: Drosophilidae). Sci. Rep. 2016, 6, 21432. [Google Scholar] [CrossRef] [PubMed]
- Erland, L.A.E.; Rheault, M.R.; Mahmoud, S.S. Insecticidal and oviposition deterrent effects of essential oils and their constituents against the invasive pest Drosophila suzukii (Matsumura) (Diptera: Drosophilidae). Crop Prot. 2015, 78, 20–26. [Google Scholar] [CrossRef]
- Renkema, J.; University of Florida, Balm, FL, USA. Unpublished work. 2017.
- Wallingford, A.K.; Connelly, H.L.; Dore Brind’Amour, G.; Boucher, M.T.; Mafra-Neto, A.; Loeb, G.M. Field evaluation of an oviposition deterrent for management of spotted-wing drosophila, Drosophila suzukii and potential nontarget effects. J. Econ. Entomol. 2016, 109, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Mori, B.A.; Evenden, M.L. Mating disruption of Coleophora deauratella (Lepidoptera: Coleophoridae) using laminate flakes in a red clover seed production fields. Pest Manag. Sci. 2014, 71, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Leite, B.L.S.; Souza, T.T.; Antoniolli, A.R.; Guimarãee, A.G.; Siqueira, R.S.; Quintans, J.S.S.; Bonjardim, L.R.; Alves, P.B.; Blank, A.F.; Botelho, M.A.; et al. Volatile constituents and behavioral changes induced by Cymbopogon winterianus leaf essential oil in rodents. Afr. J. Biotechnol. 2011, 10, 8312–8319. [Google Scholar] [CrossRef]
- Rodrigues, K.A.F.; Dias, C.N.; Amaral, F.M.M.; Moraes, D.F.C.; Mouchrek Filho, V.E.; Andrade, E.H.A.; Maia, J.G.S. Molluscicidal and larvicidal activities and essential oil composition of Cymbopogon winterianus. Pharm. Biol. 2013, 51, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Juan, L.W.; Lucia, A.; Zerba, E.N.; Harrand, L.; Marco, M.; Masuh, H.M. Chemical composition and fumigant toxicity of the essential oils from 16 species of Eucalyptus against Haematobia irritans (Diptera: Muscidae) adults. J. Econ. Entomol. 2011, 104, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Chattopadhyay, A.; Yadav, A.; Kumari, R. Variation in the chemical composition and yield of essential oil of rose-scented geranium (Pelargonium sp.) by the foliar application of metallic salts. Flavour Fragr. J. 2008, 23, 133–136. [Google Scholar] [CrossRef]
- Gallardo, A.; Picollo, M.I.; González-Audino, P.; Mougabure-Cueto, G. Insecticidal activity of individual and mixed monoterpenoids of geranium essential oil against Pediculus humans capitis (Phthiraptera: Pediculidae). J. Med. Entomol. 2012, 49, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.N.; Mallavarapu, G.R.; Baskaran, K.; Ramesh, S.; Kumar, S. Composition of the essential oils of two isomenthone-rich variants of geranium (Pelargonium sp.). Flavour Fragr. J. 1998, 13, 389–392. [Google Scholar] [CrossRef]
- Umezu, T.; Sakata, A.; Ito, H. Ambulation-promoting effect of peppermint oil and identification of its active constituents. Pharmacol. Biochem. Behav. 2001, 69, 383–390. [Google Scholar] [CrossRef]
- Rohloff, J.; Skagen, E.B.; Steen, A.H.; Beisvåg, T.; Iversen, T.-H. Essential oil composition of Norwegian peppermint (Mentha × piperita L.) and Sachalinmint [Mentha sachalinensis (Briq.) Kudô]. Acta Agric. Scand. Sect. B Plant Soil Sci. 2000, 50, 161–168. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Craker, L.E.; Xing, B. Effects of Cd, Pb, and Cu on growth and essential oil contents of dill, peppermint, and basil. Environ. Exp. Bot. 2006, 58, 9–16. [Google Scholar] [CrossRef]
- Hudaib, M.; Aburjai, T. Volatile components of Thymus vulgaris L. from wild growing and cultivated plants in Jordan. Flavour Fragr. J. 2007, 22, 322–327. [Google Scholar] [CrossRef]
- Omidbaigi, R.; Kazemi, S.; Daneshfar, E. Harvest time effecting on the essential oil content and compositions of Thymus vulgaris. J. Essent. Oil Bear. Plants 2008, 11, 162–167. [Google Scholar] [CrossRef]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal tolerances of the spotted-wing drosophila Drosopohila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, reproductive output and population growth of the fruit fly pest Drosophila suzukii (Diptera: Drosophilidae) on artificial diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Dreves, A.; Cave, A.; Lee, J. A Detailed Guide for Testing Fruit for the Presence of Spotted Wing Drosophila (SWD) Larvae; Extension Service; Oregon State University: Corvallis, OR, USA, 2014. [Google Scholar]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014. [Google Scholar]
- JMP® Pro. Statistical Analysis Software, Version 12.0.1; SAS Institute: Cary, NC, USA, 2015.
- Diepenbrock, L.M.; Rosensteel, D.O.; Hardin, J.A.; Sial, A.A.; Burrack, H.J. Season-long programs for control of Drosophila suzukii in southeastern blueberries. Crop Prot. 2016, 81, 76–84. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtyre, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Hummelbrunner, L.A.; Isman, M.B. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef] [PubMed]
- González, J.O.W.; Gutiérrez, M.M.; Murray, A.P.; Ferrero, A.A. Composition and biological activity of essential oils from Labiatae against Nezara viridula (Hemiptera: Pentatomidae) soybean pest. Pest Manag. Sci. 2011, 67, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-R.; Haribalan, P.; Son, B.-K.; Ahn, Y.-J. Fumigant toxicity of plant essential oils against Camptomyia corticalis (Diptera: Cecidomyiidae). J. Econ. Entomol. 2012, 105, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a constituent of thyme essential oil, is a positive allosteric modulator of human GABA(A) receptors and a homo-oligomeric GABA receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, D.V.; Angelov, I.; Vicente, G.; Stateva, R.P.; García-Risco, M.R.; Reglero, G.; Ibañez, E.; Fornari, T. Extraction of thymol from different varieties of thyme plants using green solvents. J. Sci. Food Agric. 2015, 95, 2901–2907. [Google Scholar] [CrossRef] [PubMed]
- Paster, N.; Menasherov, M.; Ravid, U.; Juven, B. Antifungal activity of oregano and thyme essential oils applied as fumigants against fungi attacking stored grain. J. Food Prot. 1995, 58, 81–85. [Google Scholar] [CrossRef]
- Calvet, C.; Pinochet, J.; Camprubi, A.; Estaun, V.; Rodriguez-Kabana, R. Evaluation of natural chemical compounds against root-lesion and root-knot nematodes and side-effects on the infectivity of arbuscular mycorrhizal fungi. Eur. J. Plant Pathol. 2001, 107, 601–605. [Google Scholar] [CrossRef]
- Ji, P.; Momol, M.T.; Olson, S.M.; Pradhanang, P.M.; Jones, J.B. Evaluation of thymol as a biofumigant for control for bacterial wilt of tomato under field conditions. Plant Dis. 2005, 89, 497–500. [Google Scholar] [CrossRef]
- Konstantopoulou, I.; Vassilopooulou, L.; Mavragani-Tsipidou, P.; Scouras, Z.G. Insecticidal effects of essential oils. A study of the effects of essential oils extracted from eleven Greek aromatic plants on Drosophila auraria. Experientia 1992, 48, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Vasudevan, P.; Tandon, M.; Razdan, R.K. Larvicidal and mosquito repellent action of peppermint (Mentha piperita) oil. Bioresour. Technol. 2000, 71, 267–271. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Repellent, larvicidal and pupicidal activity of essential oils and their formulations against house fly, Musca domestica. Med. Vet. Entomol. 2011, 25, 302–310. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil | Major Compounds (%) | Total % | References | ||
|---|---|---|---|---|---|
| Citronella | citronellal (34) | geraniol (22) | citronellol (12) | 68 | [32,33] |
| Eucalyptus | 1,8 cineole (68) | α—terpineol (12) | 80 | [34] | |
| Geranium | citronellol (34) | geraniol (16) 1 | citronellyl formate (9) | 71 | [35,36,37] |
| isomenthone (7) | linalool (5) | ||||
| Peppermint | menthol (45) | menthone (14) | 59 | [38,39,40] | |
| Thyme | thymol (35) | γ—terpinene (20) | ρ—cymene (20) | 75 | [41,42] |
| Compounds | Treatments | ||
|---|---|---|---|
| Thymol | Citronellol | Four-Compound Blend | |
| Thymol | 11.56 | 0.68 | 3.5 |
| Citronellol | 0.68 | 11.56 | 3.4 |
| Menthol | 0.68 | 0.68 | 4.5 |
| Geraniol | 0.68 | 0.68 | 2.2 |
| Total | 13.6 | 13.6 | 13.6 |
| Replicates | |||
| Choice | 10 | 10 | 10 |
| No-choice 23–24 July 1 | 4 | 5 | 5 |
| No-choice 30–31 July | 6 | 7 | 5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renkema, J.M.; Buitenhuis, R.; Hallett, R.H. Reduced Drosophila suzukii Infestation in Berries Using Deterrent Compounds and Laminate Polymer Flakes. Insects 2017, 8, 117. https://doi.org/10.3390/insects8040117
Renkema JM, Buitenhuis R, Hallett RH. Reduced Drosophila suzukii Infestation in Berries Using Deterrent Compounds and Laminate Polymer Flakes. Insects. 2017; 8(4):117. https://doi.org/10.3390/insects8040117
Chicago/Turabian StyleRenkema, Justin M., Rosemarije Buitenhuis, and Rebecca H. Hallett. 2017. "Reduced Drosophila suzukii Infestation in Berries Using Deterrent Compounds and Laminate Polymer Flakes" Insects 8, no. 4: 117. https://doi.org/10.3390/insects8040117
APA StyleRenkema, J. M., Buitenhuis, R., & Hallett, R. H. (2017). Reduced Drosophila suzukii Infestation in Berries Using Deterrent Compounds and Laminate Polymer Flakes. Insects, 8(4), 117. https://doi.org/10.3390/insects8040117