
The Native Hawaiian Insect Microbiome Initiative: A Critical Perspective for Hawaiian Insect Evolution

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Processing and Sequencing
2.2. Bacterial Community Analyses
2.3. Data Availability
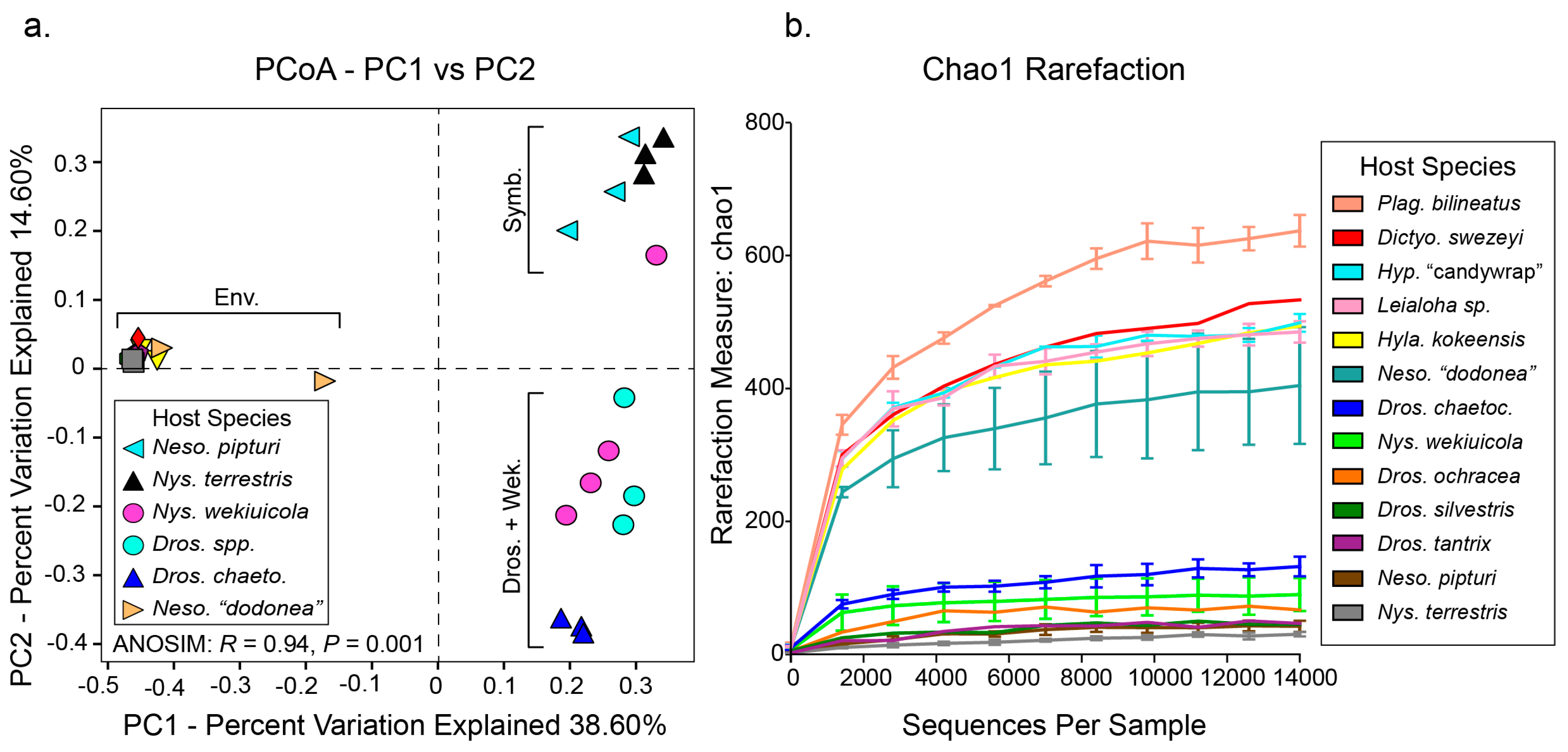
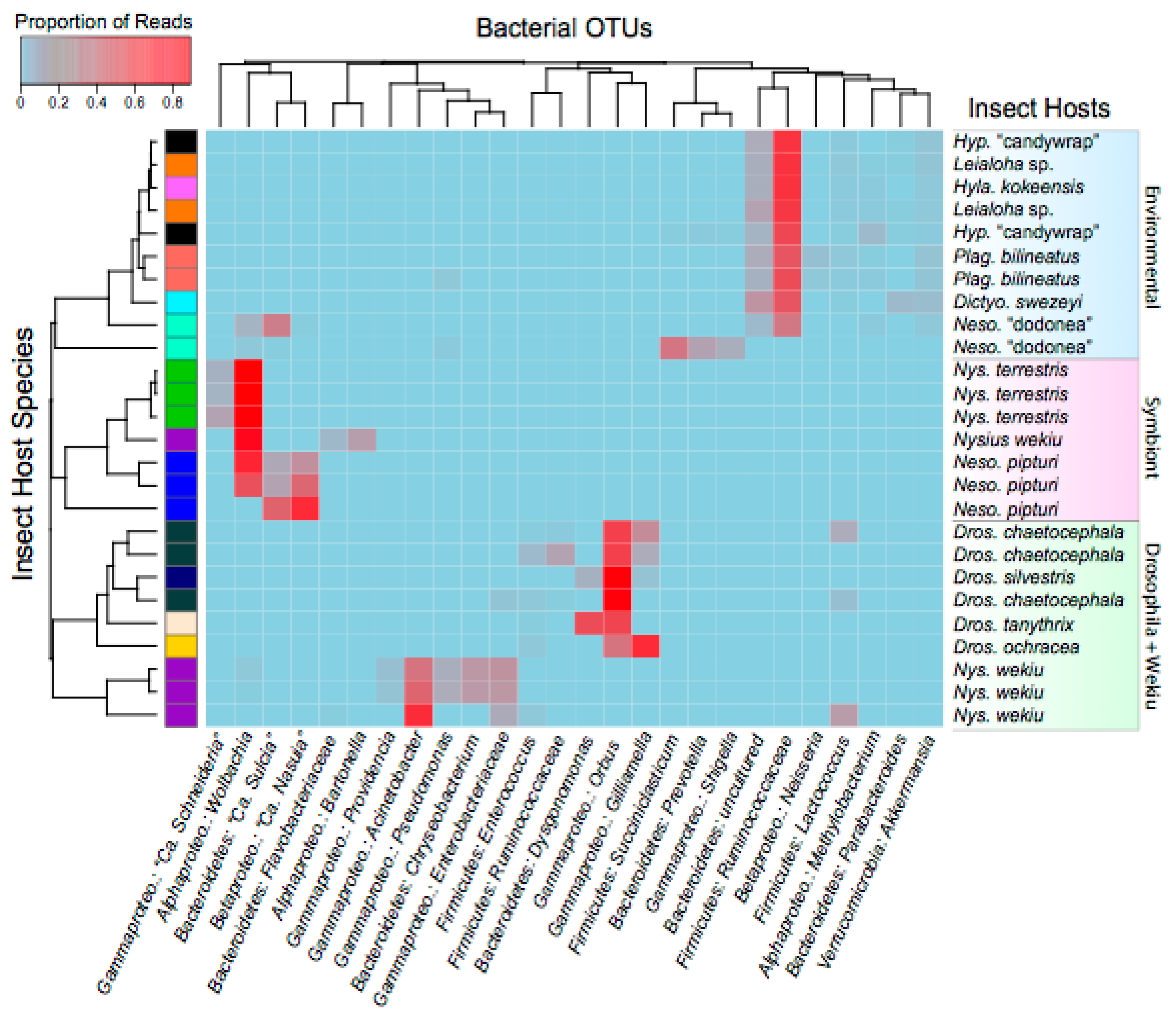
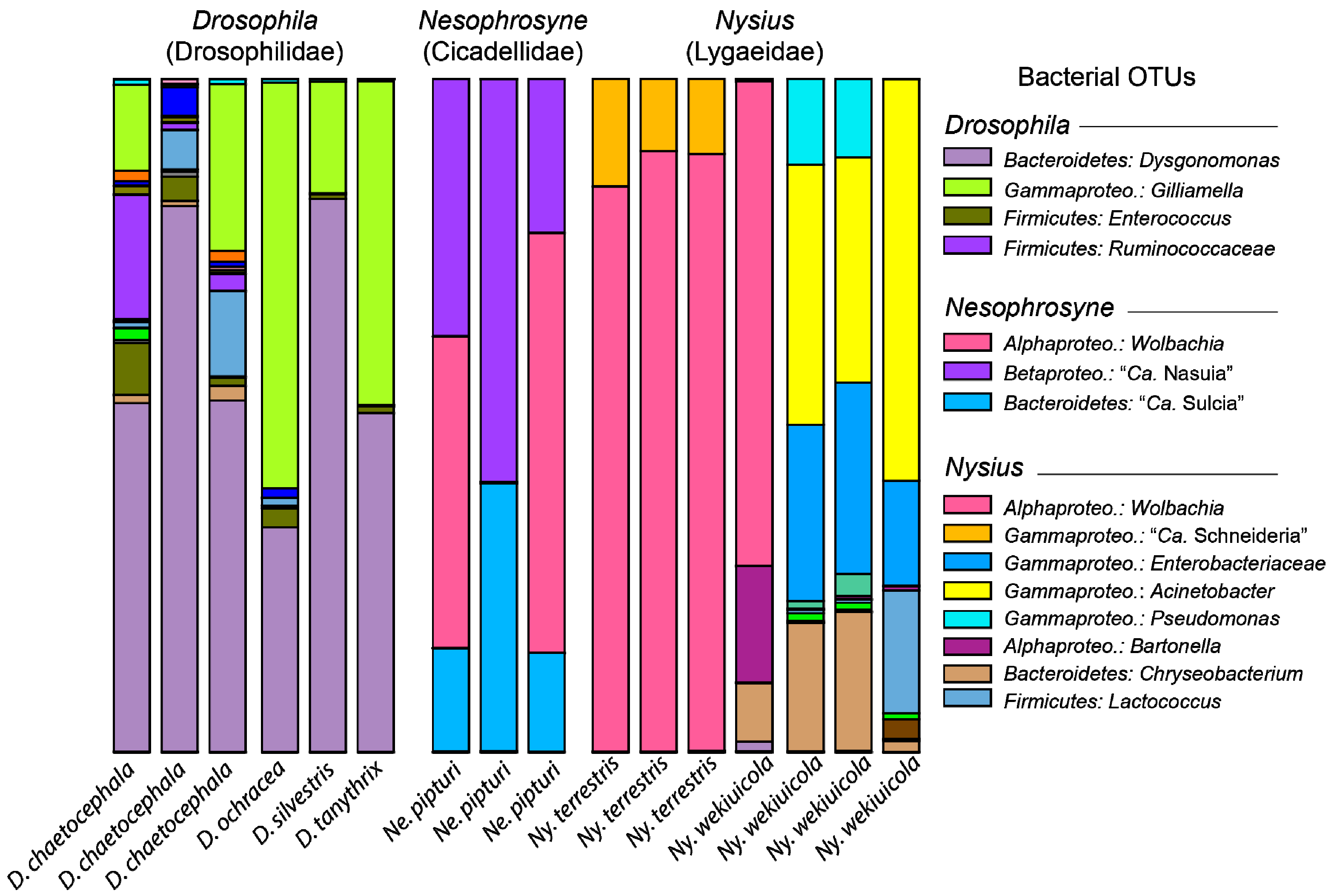
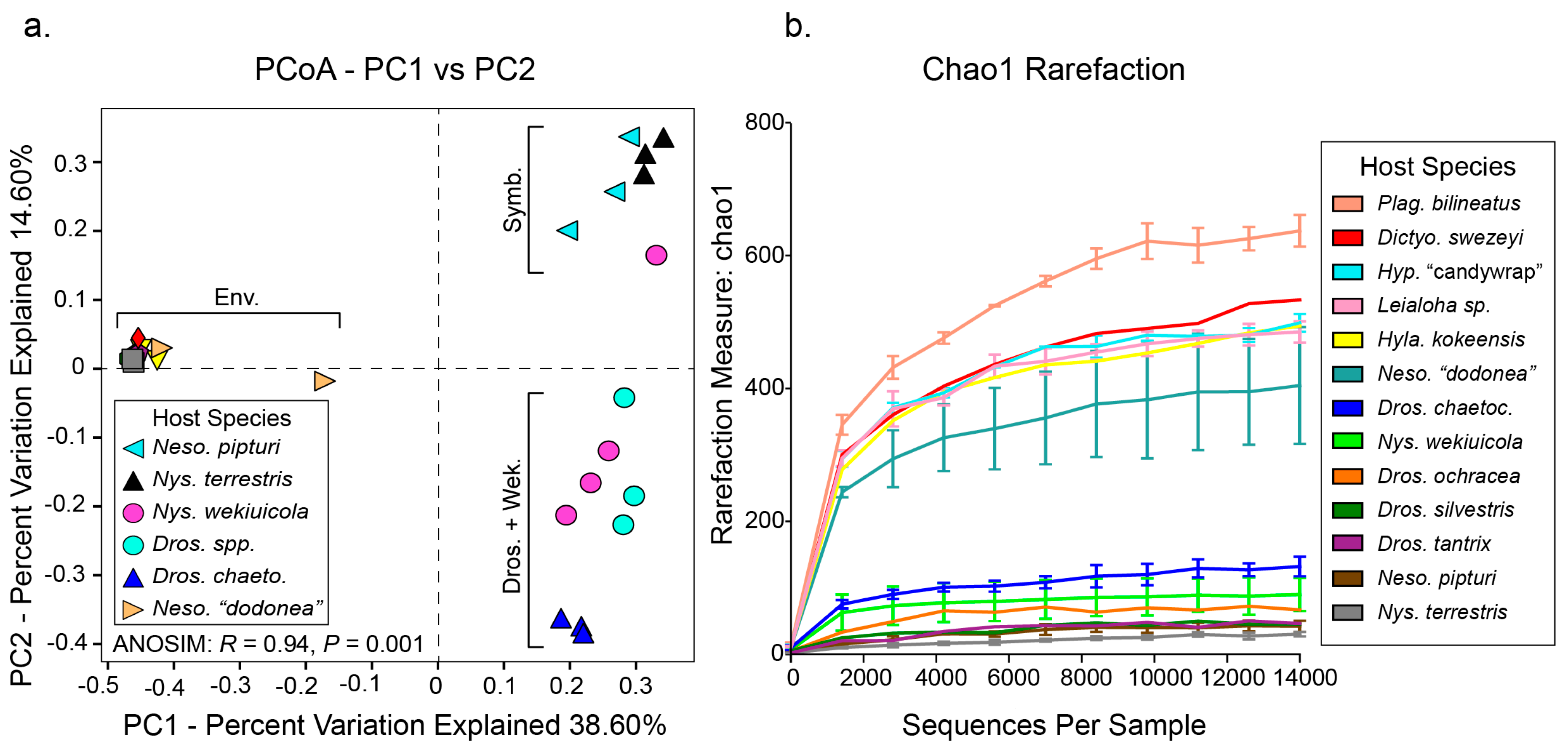
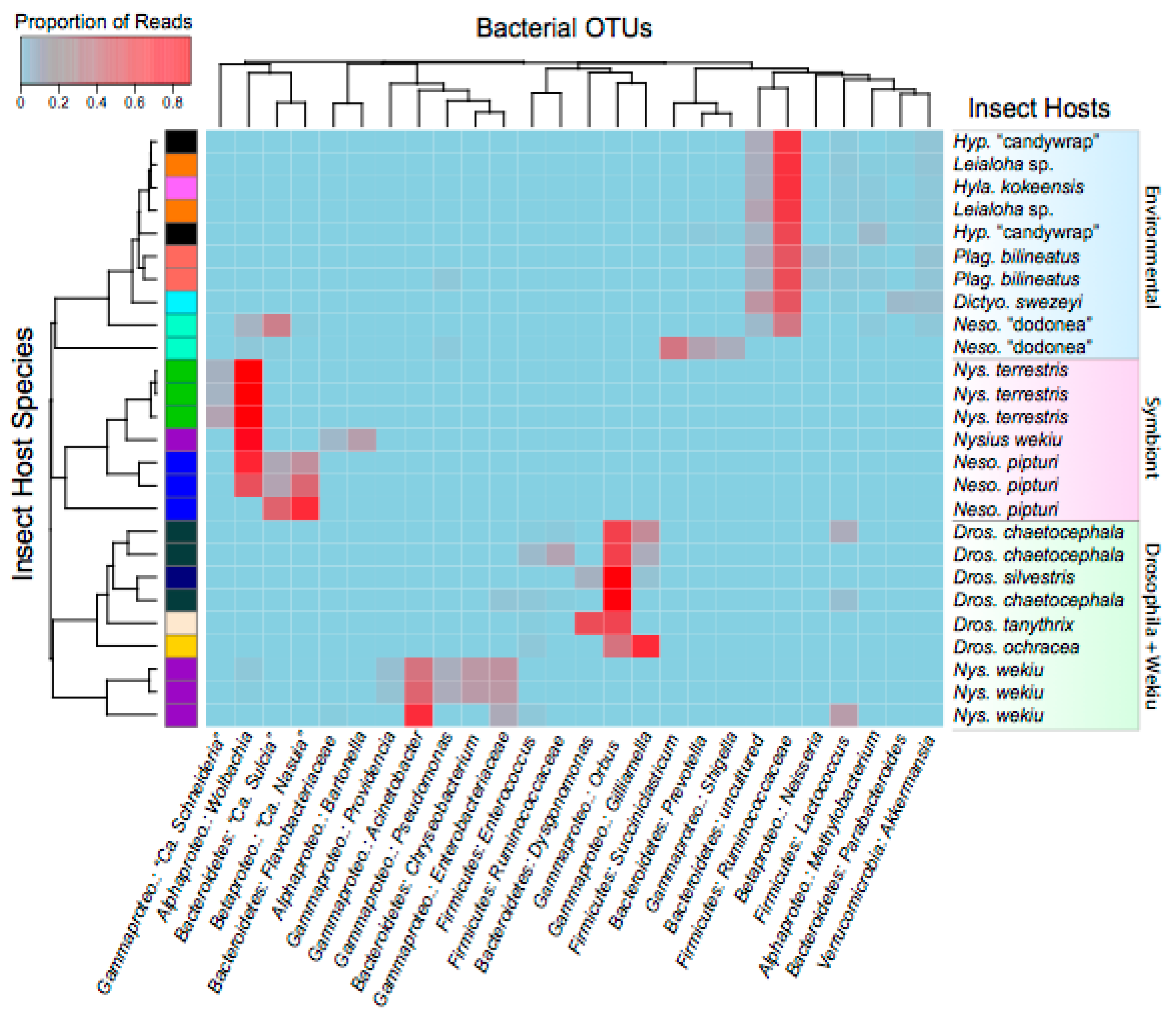
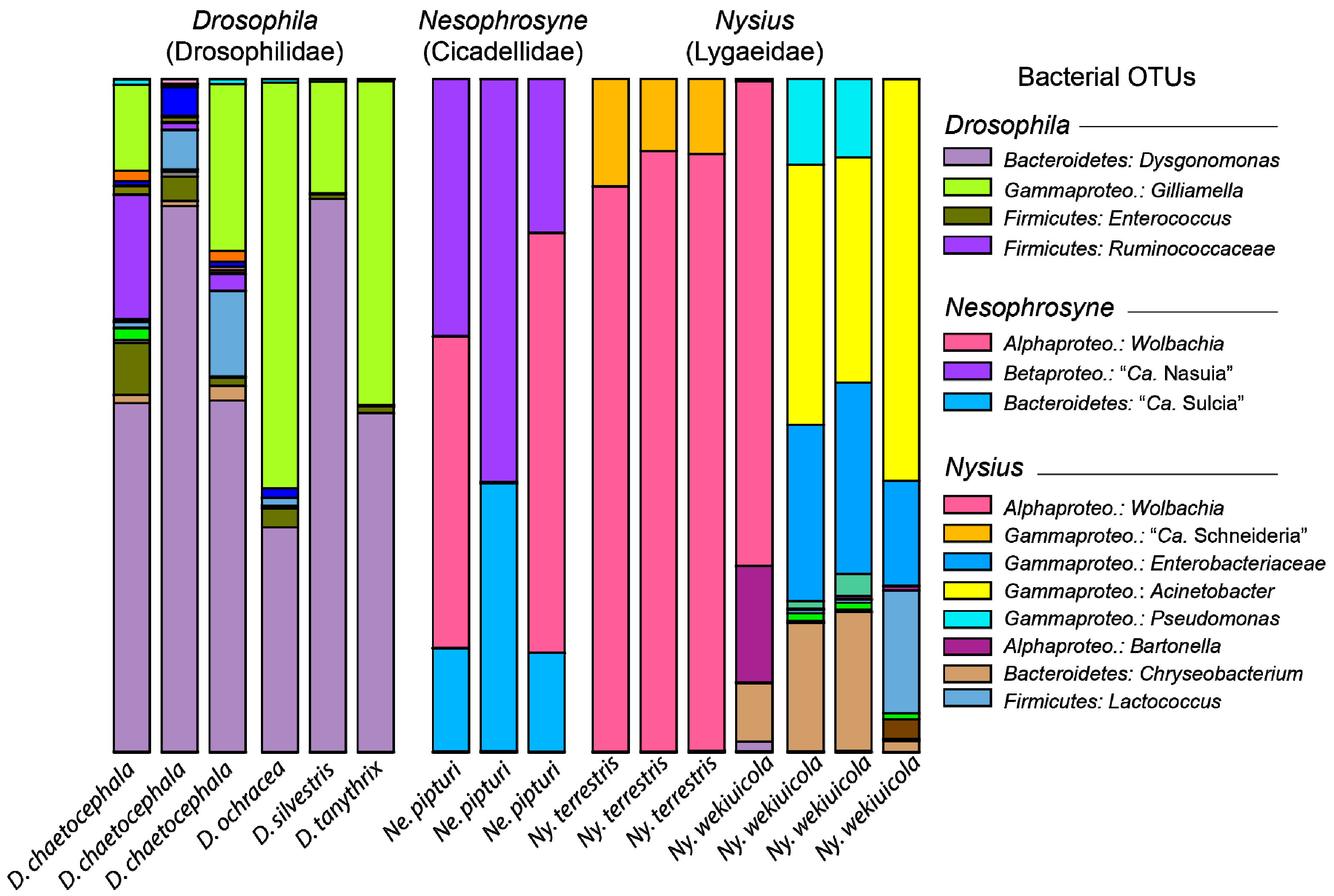
3. Results and Discussion
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, J.; Vavre, F. Bacterial symbionts in insects or the story of communities affecting communities. Philos. Trans. R. Soc. B 2011, 366, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H. Insect Symbiosis: An Introduction in Insect Symbiosis, 1st ed.; Bourtzis, K., Miller, T.A., Eds.; CRC Press LLC: Boca Raton, FL, USA, 2003; pp. 1–21. ISBN 1420005936. [Google Scholar]
- Hurst, C.J. The Mechanistic Benefits of Microbial Symbionts; Springer International Publishing: Cham, Switzerland, 2016; ISBN 9783319280684. [Google Scholar]
- Martinson, V.G.; Douglas, A.E.; Jaenike, J. Community structure of the gut microbiota in sympatric species of wild Drosophila. Ecol. Lett. 2017, 20, 629–639. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.P.; Moran, N.A. Extreme genome reduction in symbiotic bacteria. Nat. Rev. Microbiol. 2012, 10, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Muir, P.; Li, S.; Lou, S.; Wang, D.; Spakowicz, D.J.; Salichos, L.; Zhang, J.; Weinstock, G.M.; Iaacs, F.; Rozowsky, J.; et al. The real cost of sequencing: Scaling computation to keep pace with data generation. Genome Biol. 2016, 17, 1–9. [Google Scholar] [CrossRef]
- Fleischer, R.C.; McIntosh, C.E.; Tarr, C.L. Evolution on a volcanic conveyor belt: Using phylogeographic reconstructions and K–Ar-based ages of the Hawaiian Islands to estimate molecular evolutionary rates. Mol. Ecol. 1998, 7, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Price, J.P.; Clague, D.A. How old is the Hawaiian biota? Geology and phylogeny suggest recent divergence. Proc. R. Soc. Lond. B Biol. 2002, 269, 2429–2435. [Google Scholar] [CrossRef] [PubMed]
- Clague, D.A.; Sherrod, D.R.; Poland, M.; Takahashi, T.J.; Landowski, C.M. Growth and degradation of Hawaiian volcanoes in Characteristics of Hawaiian Volcanoes. Geol. Surv. Prof. Pap. 2014, 1801, 97–146. [Google Scholar] [CrossRef]
- Alison, K.E. A Natural History of the Hawaiian Islands: Selected Readings II; University of Hawaii Press: Honolulu, HI, USA, 1994; ISBN 978-0-8248-1659-9. [Google Scholar]
- Carson, H.L.; Templeton, A.R. Genetic revolutions in relation to speciation phenomena: The founding of new populations. Annu. Rev. Ecol. Syst. 1984, 15, 97–132. [Google Scholar] [CrossRef]
- Shaw, K.L.; Gillespie, R.G. Comparative phylogeography of oceanic archipelagos: Hotspots for inferences of evolutionary process. Proc. Natl. Acad. Sci. USA 2016, 113, 7986–7993. [Google Scholar] [CrossRef] [PubMed]
- Howarth, F.G.; Mull, W.P. Hawaiian Insects and Their Kin; University of Hawaii Press: Honolulu, HI, USA, 1992. [Google Scholar]
- Rubinoff, D. Phylogeography and ecology of an endemic radiation of Hawaiian aquatic case-bearing moths (Hyposmocoma: Cosmopterigidae). Philos. Trans. R. Soc. B 2008, 363, 3459–3465. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T.J.; Millam, K.C.; Mast, A.R.; Paterson, T.B.; Theim, T.J.; Hipp, A.L.; Henss, J.M.; Smith, J.F.; Wood, K.R.; Kenneth, J.S. Origin, adaptive radiation and diversification of the Hawaiian lobeliads (Asterales: Campanulaceae). Proc. R. Soc. Lond. B Biol. 2009, 276, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Heed, W.B. Host plant specificity and speciation in Hawaiian Drosophila. Taxon 1971, 20, 115–121. [Google Scholar] [CrossRef]
- Boake, C.R.B. Sexual selection and speciation in Hawaiian Drosophila. Behav. Genet. 2005, 35, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Magnacca, K.N.; Foote, D.; O’Grady, P.M. A review of the endemic Hawaiian Drosophilidae and their host plants. Zootaxa 2008, 1728, 1–58. [Google Scholar] [CrossRef]
- Roderick, G.K.; Percy, D.M. Host plant use, diversification, and coevolution: Insights from remote oceanic islands. In Specialization, Speciation, and Radiation. Evolutionary Biology of Herbivorous Insects; Tilmon, K., Ed.; University of California Press: Berkeley, CA, USA, 2008; pp. 151–161. ISBN 9780520251328. [Google Scholar]
- Bennett, G.M.; O’Grady, P.M. Host-plants shape insect diversity: Phylogeny, origin, and species diversity of native Hawaiian leafhoppers (Cicadellidae: Nesophrosyne). Mol. Phylogenet. Evol. 2012, 65, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, E.C. Insects of Hawaii Introduction, 1st ed.; University of Hawaii press: Honolulu, HI, USA, 1948; Volume 1, ISBN 978-0-8248-2427-3. [Google Scholar]
- Ashlock, P.D.; Gagne, W.C. A remarkable new micropterous-nysius species from the Aeolian Zone of Mauna-Kea, Hawaii Island (Hemiptera, Heteroptera, Lygaeidae). Int. J. Entomol. 1983, 25, 47–55. [Google Scholar]
- Eiben, J.A.; Rubinoff, D. Life history and captive rearing of the Wekiu bug (Nysius wekiuicola, Lygaeidae), an alpine carnivore endemic to the Mauna Kea volcano of Hawaii. J. Insect Conserv. 2010, 14, 701–709. [Google Scholar] [CrossRef]
- Hardy, D.E. Insects of Hawaii: Diptera Cyclorrapha II; University of Hawaii Press: Honolulu, HI, USA, 1965; Volume 12, ISBN 0-8248-0341-8. [Google Scholar]
- Montgomery, S.L. Carnivorous caterpillars: The behavior, biogeography and conservation of Eupithecia (Lepidoptera: Geometridae) in the Hawaiian Islands. GeoJournal 1983, 7, 549–556. [Google Scholar] [CrossRef]
- O’Connor, T.K.; Humphrey, P.T.; Lapoint, R.T.; Whiteman, N.K.; O’Grady, P.M. Microbial interactions and the ecology and evolution of Hawaiian Drosophilidae. Front. Microbial. 2014, 5, 616. [Google Scholar] [CrossRef]
- Ort, B.S.; Bantay, R.M.; Pantoja, N.A.; O’Grady, P.M. Fungal diversity associated with Hawaiian Drosophila host plants. PLoS ONE 2012, 7, e40550. [Google Scholar] [CrossRef] [PubMed]
- Newell, P.D.; Douglas, A.E. Interspecies interactions determine the impact of the gut microbiota on nutrient allocation in Drosophila melanogaster. Appl. Environ. Microbiol. 2014, 80, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Buchner, P. Endosymbiosis of animals with plant microorganisms. Interscience 1965, 7, 168. [Google Scholar] [CrossRef]
- Houk, E.J.; Griffiths, G.W. Intracellular symbiotes of the Homoptera. Annu. Rev. Entomol. 1980, 25, 161–187. [Google Scholar] [CrossRef]
- Shigenobu, S.; Wilson, A.C. Genomic revelations of a mutualism: The pea aphid and its obligate bacterial symbiont. Cell. Mol. Life Sci. 2011, 68, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.; Hannon, G.J. Fastx-Toolkit. FASTQ/A Short-Reads Preprocessing Tools. Available online: http:// hannonlab.cshl.edu/fastx_toolkit/ (accessed on 15 March 2017).
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Pena, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 15 March 2017).
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/index.html/ (accessed on 18 December 2017).
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Bennett, G.M.; Moran, N.A. Small, smaller, smallest: The origins and evolution of ancient dual symbioses in a phloem-feeding insect. Genome Biol. Evol. 2013, 5, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, Y.; Kikuchi, Y.; Hosokawa, T.; Koga, R.; Meng, X.Y.; Kamagata, Y.; Nikoh, N.; Fukatsu, T. Evolution of symbiotic organs and endosymbionts in lygaeid stinkbugs. ISME J. 2012, 6, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Shapira, M. Gut microbiotas and host evolution: Scaling up symbiosis. Trends Ecol. Evol. 2016, 31, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Hammer, T.J.; Janzen, D.H.; Hallwachs, W.; Jaffe, S.L.; Fierer, N. Caterpillars lack a resident gut microbiome. Proc. Natl. Acad. Sci. USA 2017, 114, 9641–9646. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct Ecol 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Liu, N.; Yan, X.; Zhang, M.; Xie, L.; Wang, Q.; Huang, Y.; Zhou, X.; Wang, S.; Zhou, Z. Microbiome of fungus-growing termites: A new reservoir for lignocellulase genes. Appl. Environ. Microbiol. 2011, 77, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.; Aksoy, S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011, 27, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [PubMed]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Matthews, K.A.; Kaufman, T.C.; Gelbart, W.M. Research resources for Drosophila: The expanding universe. Nat. Rev. Genet. 2005, 6, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Drysdale, R. FlyBase: A database for the Drosophila research community. Methods Mol. Biol. 2008, 420, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Bellen, H.J.; Tong, C.; Tsuda, H. 100 years of Drosophila research and its impact on vertebrate neuroscience: A history lesson for the future. Nat. Rev. Neurosci. 2010, 11, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Carson, H.L.; Hardy, D.E.; Spieth, H.T.; Stone, W.S. The evolutionary biology of the Hawaiian Drosophilidae. In Essays in Evolution and Genetics in Honor of Theodosius Dobzhansky; Hecht, M.K., Steere, W.C., Eds.; Springer: Boston, MA, USA, 1970; pp. 437–543. ISBN 978-1-4615-9587-8. [Google Scholar]
- Carson, H.L.; Kaneshiro, K.Y. Drosophila of Hawaii: Systematics and ecological genetics. Annu. Rev. Ecol. Syst. 1976, 7, 311–345. [Google Scholar] [CrossRef]
- Kambysellis, M.P.; Craddock, E.M. Ecological and reproductive shifts in the diversification of the endemic Hawaiian Drosophila. In Molecular Evolution and Adaptive Radiation; Givnish, T.J., Sytsma, K.J., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 475–509. ISBN 0521779294. [Google Scholar]
- Martinson, V.G.; Carpinteyro-Ponce, J.; Moran, N.A.; Markow, T.A. A distinctive and host-restricted gut microbiota in populations of a cactophilic Drosophila species. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host–microbe model system. PLoS genet. 2011. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of acquisition of the gut microbiota of the honey bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [PubMed]
- Pramono, A.K.; Sakamoto, M.; Iino, T.; Hongoh, Y.; Ohkuma, M. Dysgonomonas termitidis sp. nov., isolated from the gut of the subterranean termite Reticulitermes speratus. Int. J. Syst. Evol. Microbiol. 2015, 65, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Brune, A.; Dietrich, C. The gut microbiota of termites: Digesting the diversity in the light of ecology and evolution. Annu. Rev. Microbiol. 2015, 69, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Tran, P.; Gerardo, N.M. Symbiosis and Insect Diversification: An Ancient Symbiont of Sap-Feeding Insects from the Bacterial Phylum Bacteroidetes. Appl. Environ. Microbiol. 2005, 71, 8802–8810. [Google Scholar] [CrossRef] [PubMed]
- Bressan, A.; Mulligan, K.L. Localization and morphological variation of three bacteriome-inhabiting symbionts within a planthopper of the genus Oliarus (Hemiptera: Cixiidae). Environ. Microbiol. Rep. 2013, 5, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Sandström, J.; Moran, N. How nutritionally imbalanced is phloem sap for aphids? Entomol. Exp. Appl. 1999, 91, 203–210. [Google Scholar] [CrossRef]
- McCutcheon, J.P.; Moran, N.A. Functional convergence in reduced genomes of bacterial symbionts spanning 200 My of evolution. Genome Biol. Evolut. 2010, 2, 708–718. [Google Scholar] [CrossRef]
- Bennett, G.M.; Moran, N.A. Heritable symbiosis: The advantages and perils of an evolutionary rabbit hole. Proc. Natl. Acad. Sci. USA 2015, 112, 10169–10176. [Google Scholar] [CrossRef] [PubMed]
- Sailendharan, S.; Kost, C.; Kaltenpoth, M. Symbiont Acquisition and Replacement as a Source of Ecological Innovation. Trends Microbiol. 2017, 5, 375–390. [Google Scholar] [CrossRef]
- Toenshoff, E.R.; Gruber, D.; Horn, M. Co-evolution and symbiont replacement shaped the symbiosis between adelgids (Hemiptera: Adelgidae) and their bacterial symbionts. Environ. Microbiol. 2012, 14, 1284–1295. [Google Scholar] [CrossRef] [PubMed]
- Koga, R.; Bennett, G.M.; Cryan, J.R.; Moran, N.A. Evolutionary replacement of obligate symbionts in an ancient and diverse insect lineage. Environ. Microbiol. 2013, 15, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia? A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.; Miller, W.J.; Riegler, M. Arthropods shopping for Wolbachia. In Manipulative Tenants: Bacteria Associated with Arthropods; Zchori-Fein, E., Bourtzis, K., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 149–174. ISBN 9781439827499. [Google Scholar]
- Bennett, G.M.; Pantoja, N.A.; O’Grady, P.M. Diversity and phylogenetic relationships of Wolbachia in Drosophila and other native Hawaiian insects. Fly 2012, 6, 273–283. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, L.; Plichart, C.; Sang, A.C.; Brelsfoard, C.L.; Bossin, H.C.; Dobson, S.L. Open release of male mosquitoes infected with a Wolbachia biopesticide: Field performance and infection containment. PLoS Negl. Trop. Dis. 2012, 6, e1797. [Google Scholar] [CrossRef] [PubMed]
- Blaxter, M.L. The promise of a DNA taxonomy. Philos. Trans. R. Soc. B 2004, 359, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Family | Species | Collection Location | Feeding Behavior | Replicates |
|---|---|---|---|---|
| Cerambycidae | Plagithmysus bilineatus | Hawaii Island, HI, Upper Wailuku | Herbivorous | 3 2 |
| Cicadellidae | Nesophrosyne pipturi | Oahu, HI, Manoa Cliff Tr. | Sap-feeding | 3 |
| Cicadellidae | Nesophrosyne “dodonea” 1 | Oahu, HI, Diamond Head | Sap-feeding | 2 |
| Lygaeidae | Nysius terrestris | Hawaii Island, HI, Mauna Kea | Sap/seed-feeding | 3 |
| Lygaeidae | Nysius wekiuicola | Hawaii Island, HI, Mauna Kea | Carnivorous | 5 2 |
| Delphacidae | Dictyophorodelphax swezeyi | Kauai, HI, Kokee State Park | Sap/seed-feeding | 1 |
| Delphacidae | Leialoha sp. 1 | Oahu, HI, Kaala Road | Sap/seed-feeding | 3 2 |
| Drosophilidae | Drosophila chaetocephala | Oahu, HI, Mt Kaala | Detritivorous | 3 |
| Drosophilidae | Drosophila ochracea | Hawaii Island, HI | Detritivorous | 1 3 |
| Drosophilidae | Drosophila silvestris | Hawaii Island, HI | Detritivorous | 1 3 |
| Drosophilidae | Drosophila tanythrix | Hawaii Island, HI | Detritivorous | 1 3 |
| Cosmopterigidae | Hyposmocoma “sp. candywrap-case” 1 | Oahu, HI, Palikea | Lichenivorous | 3 2 |
| Colletidae | Hylaeus kokeensis | Kauai, HI | Flower Feeding | 1 |
| Colletidae | Hylaeus kauaiensis | Kauai, HI | Flower Feeding | 1 2 |
| Family | Seq ID | Species | Raw Reads | Merged Fragments | Shannon Diversity Index | Pielou’s Evenness Index |
|---|---|---|---|---|---|---|
| Cerambycidae | IM_001 | Plagithmysus bilineatus | 62,589 | 4067 | N/A | N/A |
| IM_002 | Plagithmysus bilineatus | 210,792 | 32,601 | 2.78 | 0.519 | |
| IM_003 | Plagithmysus bilineatus | 279,336 | 48,751 | 2.92 | 0.528 | |
| Cicadellidae | IM_006 | Nesophrosyne pipturi | 281,608 | 28,096 | 1.19 | 0.498 |
| IM_007 | Nesophrosyne pipturi | 285,705 | 54,199 | 1.03 | 0.402 | |
| IM_008 | Nesophrosyne pipturi | 408,688 | 37,665 | 1.08 | 0.437 | |
| IM_064 | Nesophrosyne “dodonea” 1 | 140,222 | 20,007 | 3.54 | 0.713 | |
| IM_065 | Nesophrosyne “dodonea” 1 | 275,866 | 40,540 | 2.32 | 0.462 | |
| Colletidae | IM_054 | Hylaeus kokeensis | 135,535 | 23,067 | 2.23 | 0.453 |
| IM_057 | Hylaeus kauaiensis | 80,603 | 10,664 | N/A | N/A | |
| Cosmopterigidae | IM_047 | Hyposmocoma sp. 1 (candywrap-case) | 68,231 | 7180 | N/A | N/A |
| IM_048 | Hyposmocoma sp. 1 (candywrap-case) | 332,269 | 52,954 | 2.40 | 0.448 | |
| IM_049 | Hyposmocoma sp. 1 (candywrap-case) | 141,714 | 20,496 | 2.58 | 0.509 | |
| Delphacidae | IM_029 | Dictyophorodelphax swezeyi | 157,229 | 30,539 | 2.64 | 0.523 |
| IM_041 | Leialoha sp. 1 | 180,872 | 28,045 | 2.39 | 0.465 | |
| IM_042 | Leialoha sp. 1 | 92,971 | 6127 | N/A | N/A | |
| IM_043 | Leialoha sp. 1 | 214,609 | 32,972 | 2.28 | 0.442 | |
| Drosophilidae | IM_044 | Drosophila chaetocephala | 205,429 | 30,495 | 2.66 | 0.652 |
| IM_045 | Drosophila chaetocephala | 222,912 | 37,809 | 1.45 | 0.378 | |
| IM_046 | Drosophila chaetocephala | 208,202 | 29,385 | 2.33 | 0.620 | |
| IM_050 | Drosophila ochracea | 193,518 | 32,766 | 1.57 | 0.524 | |
| IM_051 | Drosophila silvestris | 251,611 | 58,065 | 1.18 | 0.416 | |
| IM_052 | Drosophila tanythrix | 317,357 | 58,839 | 0.87 | 0.315 | |
| Lygaeidae | IM_015 | Nysius terrestris | 362,988 | 44,577 | 0.56 | 0.226 |
| IM_016 | Nysius terrestris | 421,244 | 33,881 | 0.45 | 0.230 | |
| IM_017 | Nysius terrestris | 370,619 | 34,477 | 0.45 | 0.230 | |
| IM_025 | Nysius wekiuicola | 365,402 | 36,829 | 0.92 | 0.331 | |
| IM_059 | Nysius wekiuicola | 108,745 | 14,561 | 2.23 | 0.597 | |
| IM_060 | Nysius wekiuicola | 121,857 | 19,236 | 2.26 | 0.604 | |
| IM_062 | Nysius wekiuicola | 127,518 | 15,219 | 2.11 | 0.580 | |
| IM_063 | Nysius wekiuicola | 108,665 | 10,776 | N/A | N/A | |
| Control | C-O | N/A | 1297 | 263 | N/A | N/A |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poff, K.E.; Stever, H.; Reil, J.B.; Seabourn, P.; Ching, A.J.; Aoki, S.; Logan, M.; Michalski, J.R.; Santamaria, J.; Adams, J.W.; et al. The Native Hawaiian Insect Microbiome Initiative: A Critical Perspective for Hawaiian Insect Evolution. Insects 2017, 8, 130. https://doi.org/10.3390/insects8040130
Poff KE, Stever H, Reil JB, Seabourn P, Ching AJ, Aoki S, Logan M, Michalski JR, Santamaria J, Adams JW, et al. The Native Hawaiian Insect Microbiome Initiative: A Critical Perspective for Hawaiian Insect Evolution. Insects. 2017; 8(4):130. https://doi.org/10.3390/insects8040130
Chicago/Turabian StylePoff, Kirsten E., Heather Stever, Jonathan B. Reil, Priscilla Seabourn, Alexander J. Ching, Sayaka Aoki, Mitchel Logan, Jennifer R. Michalski, Jessika Santamaria, Jesse W. Adams, and et al. 2017. "The Native Hawaiian Insect Microbiome Initiative: A Critical Perspective for Hawaiian Insect Evolution" Insects 8, no. 4: 130. https://doi.org/10.3390/insects8040130
APA StylePoff, K. E., Stever, H., Reil, J. B., Seabourn, P., Ching, A. J., Aoki, S., Logan, M., Michalski, J. R., Santamaria, J., Adams, J. W., Eiben, J. A., Yew, J. Y., Ewing, C. P., Magnacca, K. N., & Bennett, G. M. (2017). The Native Hawaiian Insect Microbiome Initiative: A Critical Perspective for Hawaiian Insect Evolution. Insects, 8(4), 130. https://doi.org/10.3390/insects8040130