Mating Behavior of Rosalia batesi (Coleoptera: Cerambycidae) Is Mediated by Male-Produced Sex Pheromones


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Field Observations on the Appearance of Adult Beetles
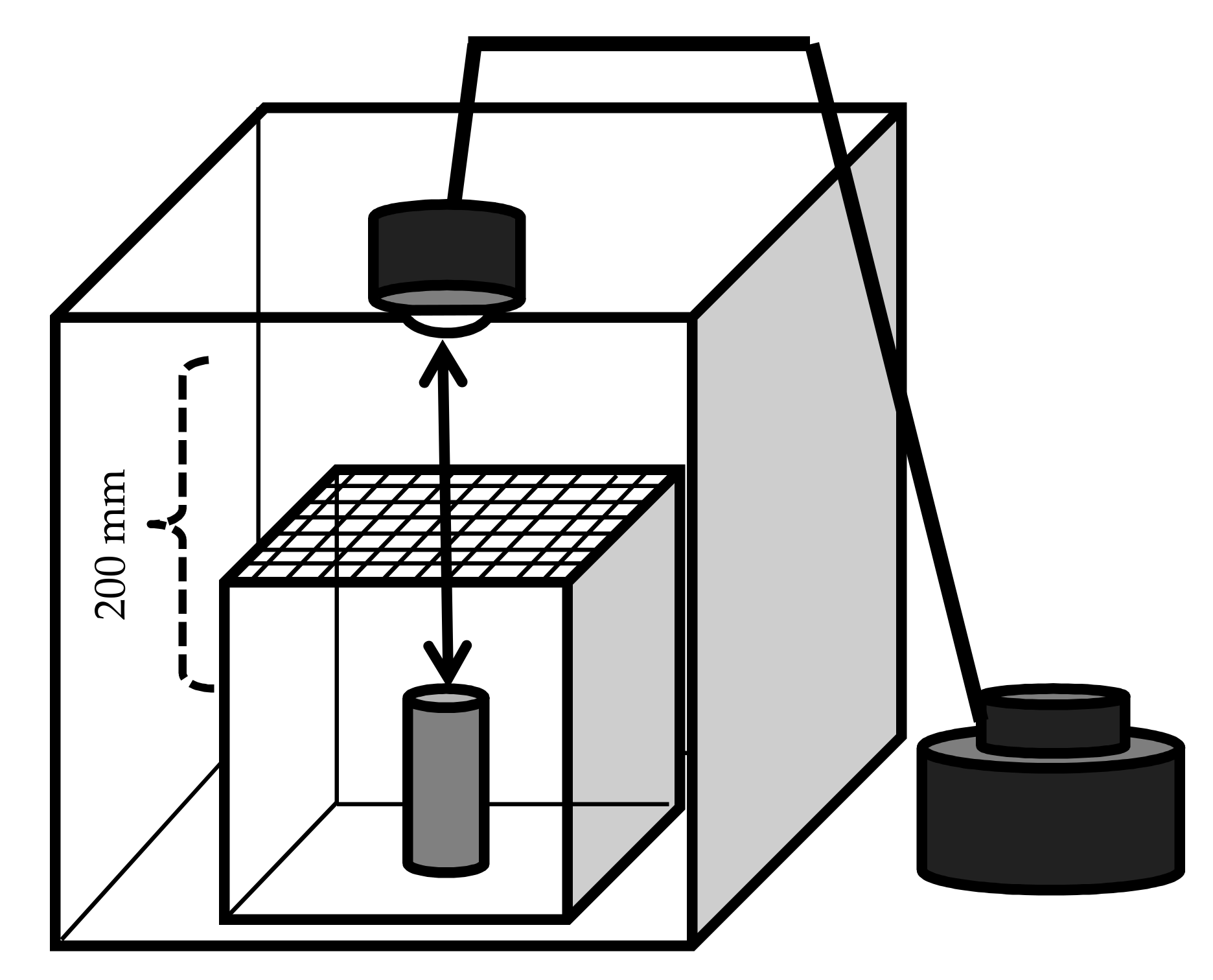
2.3. Laboratory Observations of Sexual Attraction and Subsequent Mating Behavior
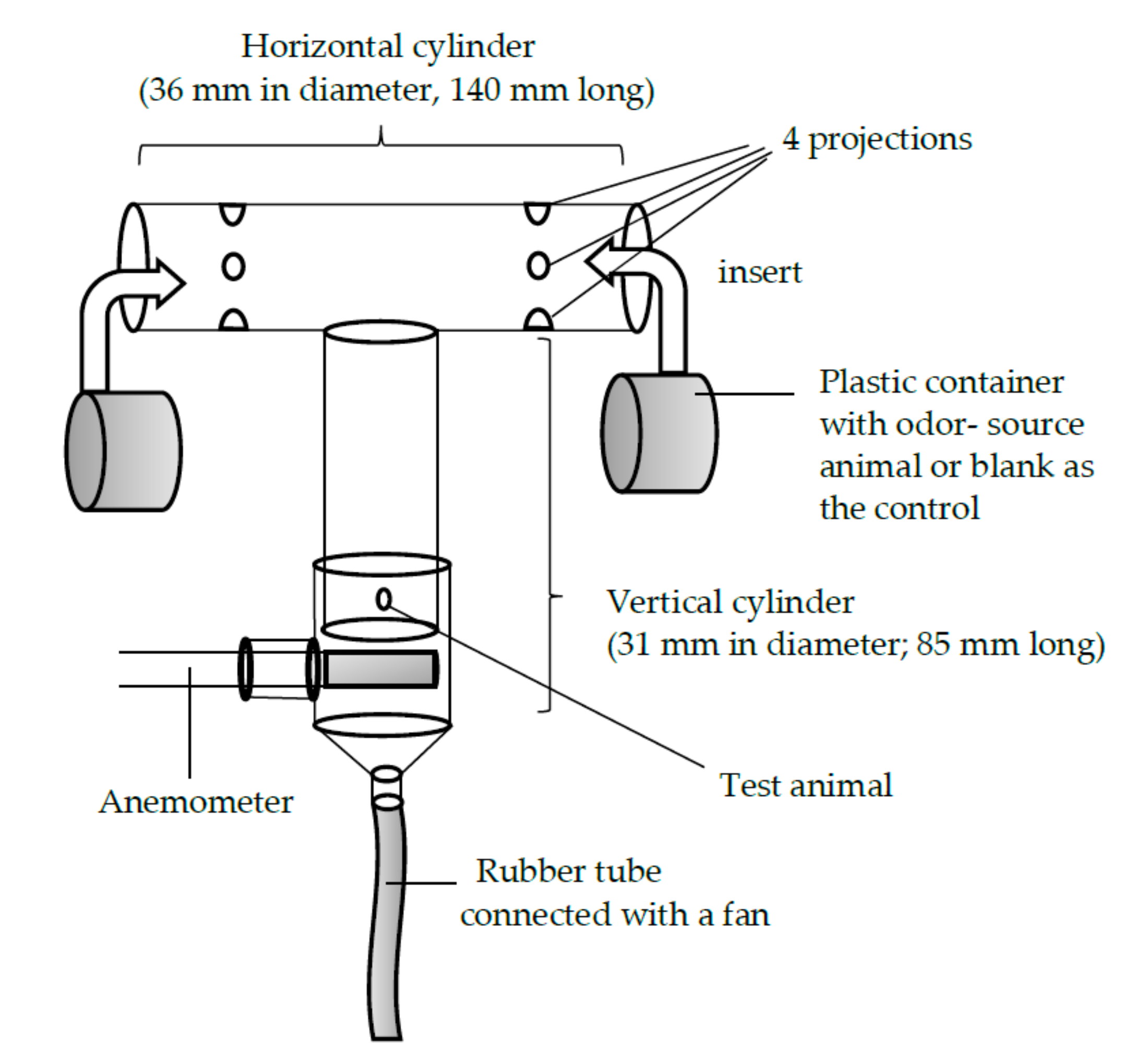
2.4. Laboratory Bioassay Using a T-Shaped Olfactometer
2.5. Laboratory Observation of Male Calling Behavior with Different Light Intensity and Temperature Regimes
2.6. Statistical Analyses
3. Results
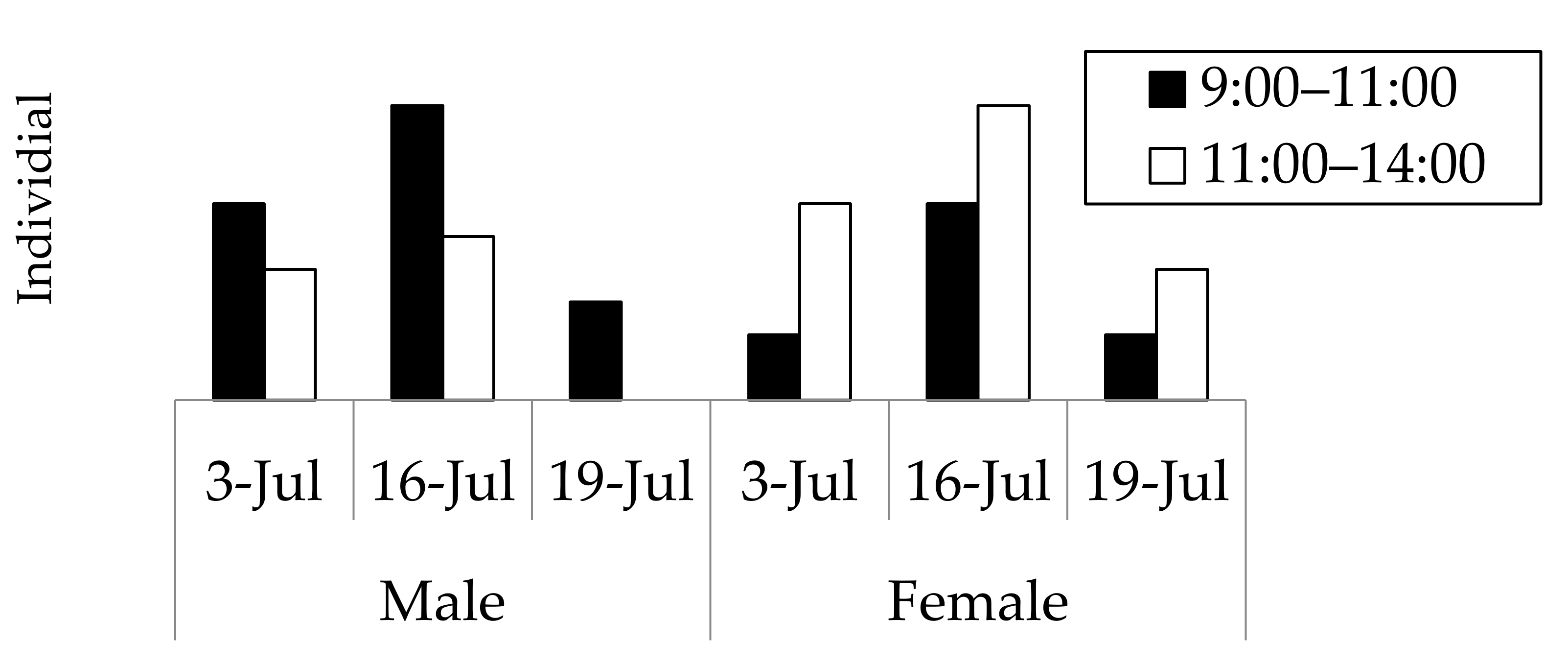
3.1. Field Observations of the Appearance of Adult Beetles
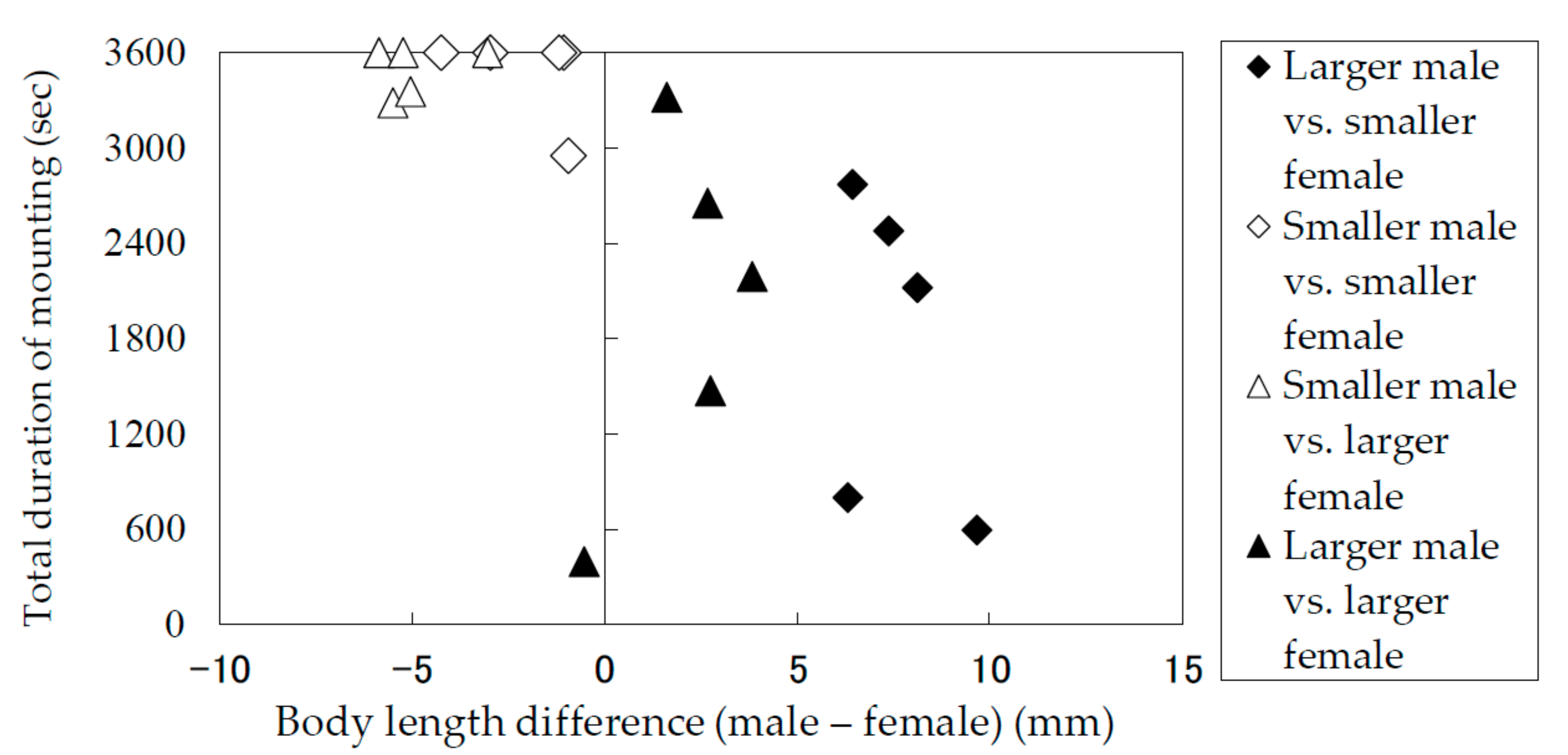
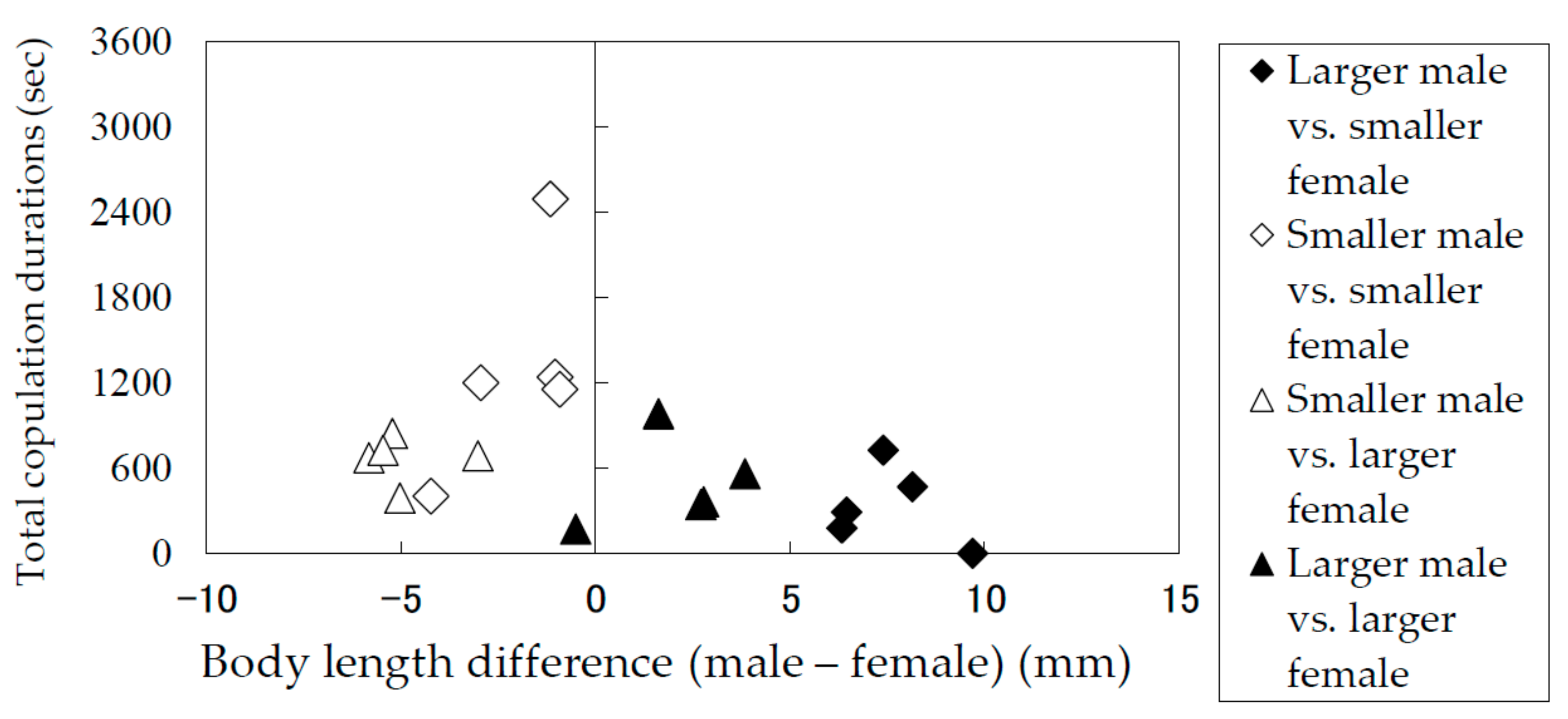
3.2. Durations of Mounting and Copulation in Relation to Body Lengths of Males and Females
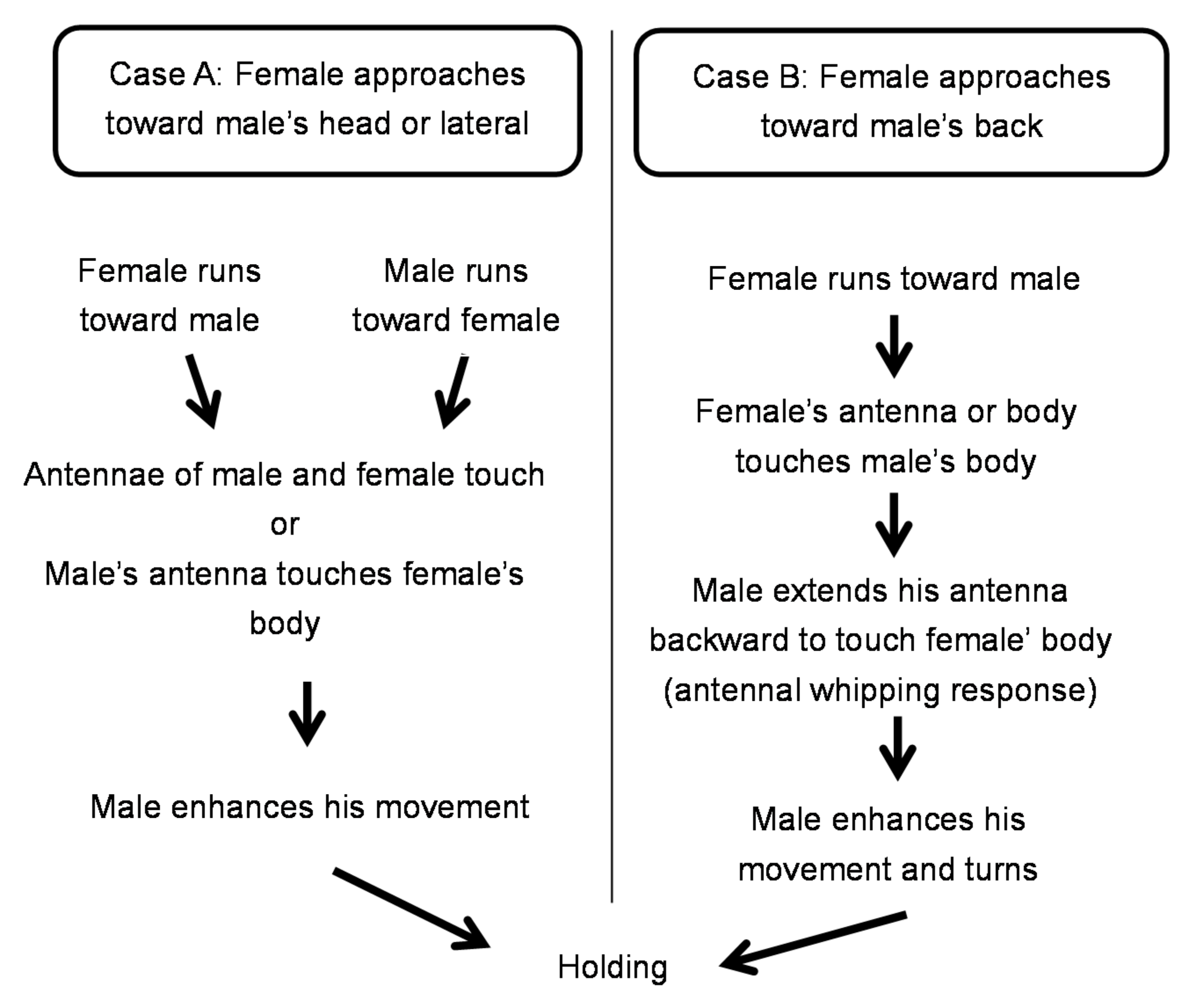
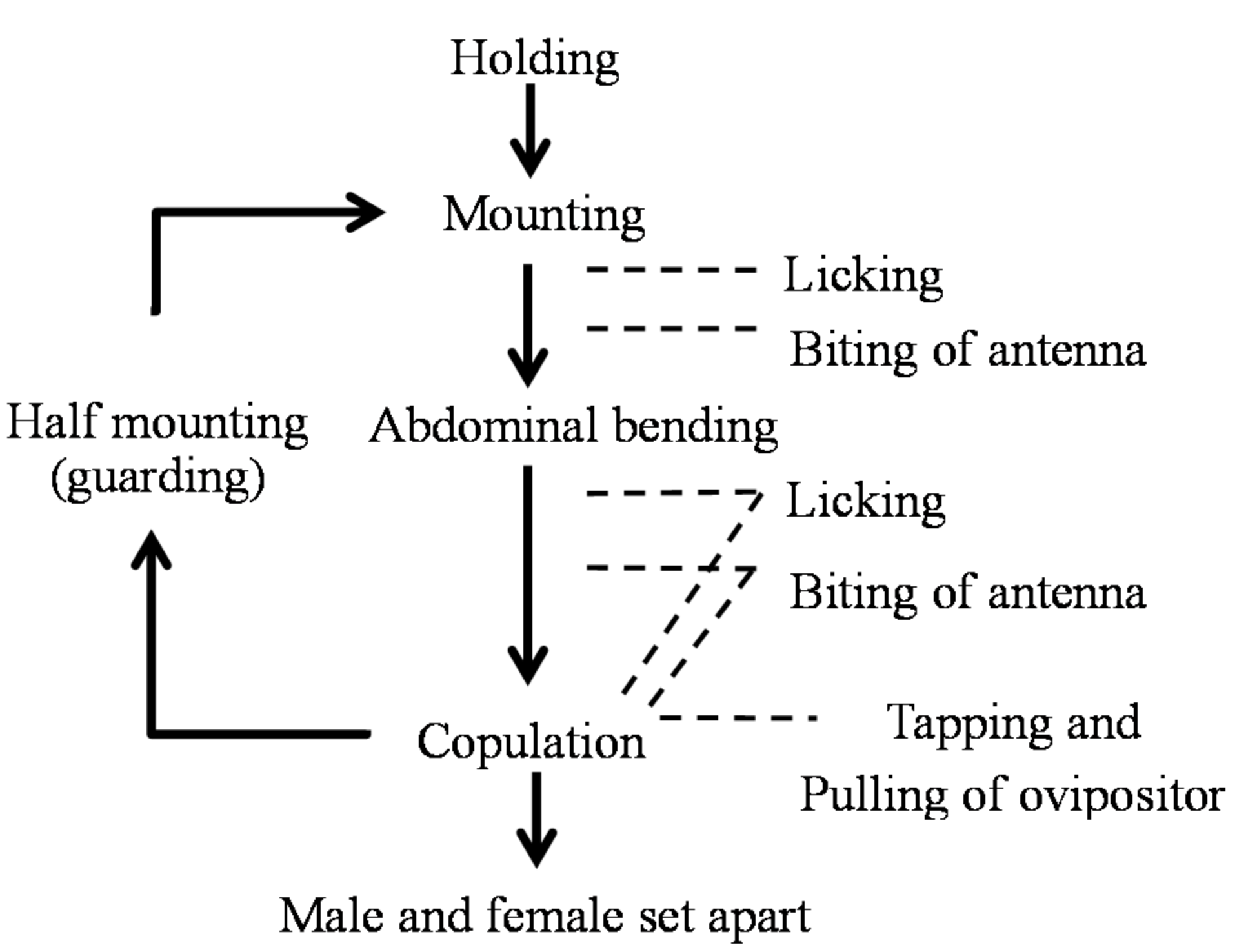
3.3. Ethogram of Mating Behavior
3.4. Male Behavior in the Push-Up Stance (Calling Behavior) and the Structure of the Male Abdominal Tip
3.5. Male and Female Choices in the T-Shaped Olfactometer
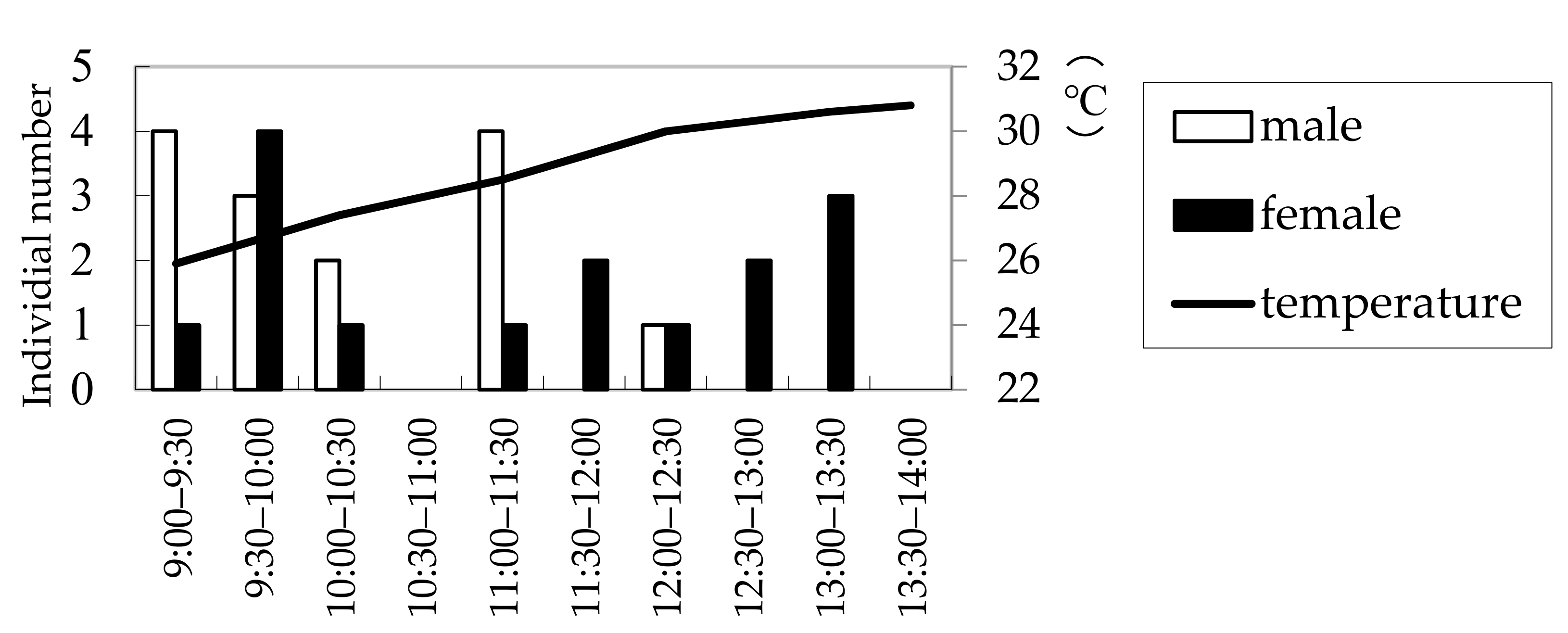
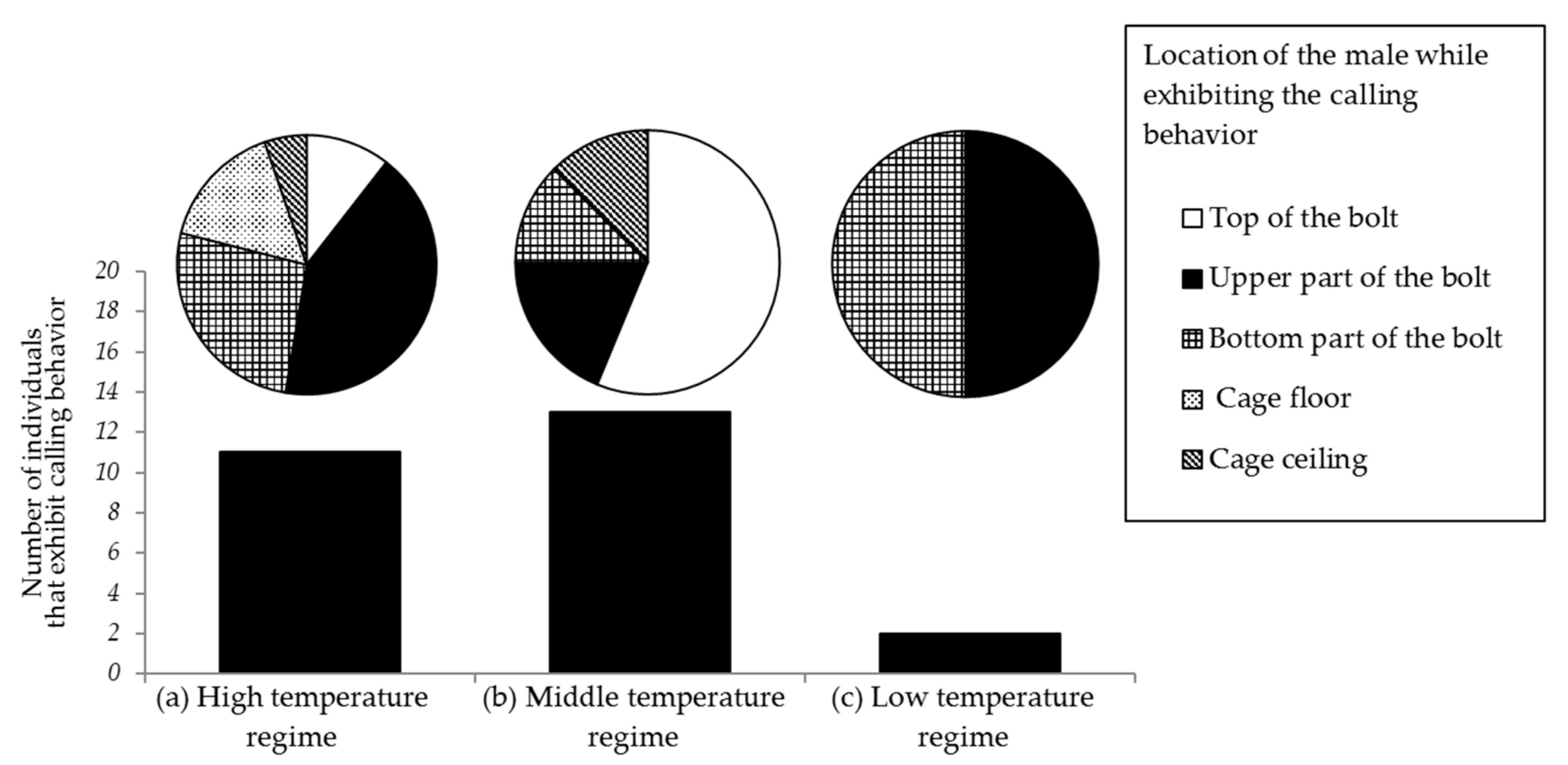
3.6. Effect of Temperature and Light Intensity on Male Calling Behavior
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Iwabuchi, K. Mating behavior of cerambycid beetles (In Japanese). Plant Prot. 1987, 41, 44–49. [Google Scholar]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae) V: Female mounting behavior. J. Ethol. 1987, 5, 131–136. [Google Scholar] [CrossRef]
- Iwabuchi, K. Reproductive behavior of cerambycid beetles (In Japanese). Nat. Insects 1995, 30, 2–6. [Google Scholar]
- Fukaya, M. Recent advances in sex pheromone studies on the white-spotted longicorn beetle. Anoplophora malasiaca. JARQ 2003, 37, 83–88. [Google Scholar] [CrossRef]
- Fukaya, M. Effects of male body size on mating activity and female mate refusal in the yellow-spotted longicorn beetle, Psacothea hilaris (Pascoe) (Coleoptera: Cerambycidae): Are small males inferior in mating? Appl. Entomol. Zool. 2004, 39, 603–609. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Lacey, E.S.; Ray, A.M.; Hanks, L.M. Calling behavior of the cerambycid beetle Neoclytus acuminatus acuminatus (F). J. Insect Behav. 2007, 20, 117–128. [Google Scholar] [CrossRef]
- Takakuwa, M. World’s Rosalia (In Japanese). In The Rosalia World; Takakuwa, M., Ed.; Mushi-Sha: Tokyo, Japan, 1998; pp. 35–100. [Google Scholar]
- Duelli, P.; Wermelinger, B. Rosalia alpina L.: Un cerambicide raro ed emblematico. Sherwood Foreste Alberi Oggi 2005, 114, 19–25. [Google Scholar]
- Iwata, R.; Aoki, M.; Nozaki, T.; Yamaguchi, M. Some notes on the biology of a hardwood-log-boring beetle, R. batesi Harold (Coleoptera: Cerambycidae), with special reference to its occurrences in a building and a suburban lumberyard. Jpn. J. Environ. Entomol. Zool. 1998, 9, 83–97. [Google Scholar]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef] [PubMed]
- Hanks, L.M.; Millar, J.G. Field bioassays of cerambycid pheromones reveal widespread parsimony of pheromone structures, enhancement by host plant volatiles, and antagonism by components from heterospecifics. Chemoecology 2013, 23, 21–44. [Google Scholar] [CrossRef]
- Takakuwa, M. Distributional expansion of a cerambycine longicon beetle, R. batesi, in southern Fossa Magna area, central Japan, with special references to its surroundings (In Japanese). Gekkan-Mushi 2000, 353, 2–17. [Google Scholar]
- Wickham, J.D.; Harrison, R.D.; Lu, W.; Guo, Z.; Millar, J.G.; Hanks, L.M.; Chen, Y. Generic lures attract cerambycid beetles in a tropical montane rain forest in southern China. J. Econ. Entomol. 2014, 107, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, D.E.; Hanks, L.M.; Lacey, E.S.; Barbour, J.D. First documentation of a volatile sex pheromone in a longhorned beetle (Coleoptera: Cerambycidae) of the primitive subfamily Prioninae. Ann. Entomol. Soc. Am. 2006, 99, 718–722. [Google Scholar] [CrossRef]
- Millar, J.G.; Hanks, L.M.; Moreira, J.A.; Barbour, J.D.; Lacey, E.S. Pheromone chemistry of cerambycid beetles. In Chemical Ecology of Wood-Boring Insects; Nakamuta, K., Millar, J.G., Eds.; Forestry and Forest Products Research Institute: Tsukuba, Japan, 2009; pp. 52–79. [Google Scholar]
- Fierke, M.K.; Skabeikis, D.D.; Millar, J.G.; Teale, S.A.; McElfresh, J.S.; Hanks, L.M. Identification of a male-produced aggregation pheromone for Monochamus scutellatus scutellatus and an attractant for the congener Monochamus notatus (Coleoptera: Cerambycidae). J. Econ. Entomol. 2012, 105, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Arakawa, M.; Kiyota, R.; Hoshino, K.; Ando, T. Carbonyl reduction in the biosynthesis of a male sex pheromone secreted by the grape borer Xylotrechus pyrrhoderus. J. Chem. Ecol. 2014, 40, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Essig, E.O. The California-laurel borer Rosalia funebris Mots. (Coleoptera: Cerambycidae). Pan-Pac. Entomol. 1943, 19, 91–92. [Google Scholar]
- Ray, A.M.; Lacey, E.S.; Hanks, L.M. Predicted taxonomic patterns in pheromone production by longhorned beetles. Naturwissenschaften 2006, 93, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.M.; Millar, J.G.; McElfresh, J.S.; Swift, I.P.; Barbour, J.D.; Hanks, L.M. Male-produced aggregation pheromone of the cerambycid beetle Rosalia funebris. J. Chem. Ecol. 2009, 35, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Herter, W. Der Alpenbock, Rosalia alpina (Coleoptera, Cerambycidae). Mitt. Entomol. Ges. Basel 2002, 52, 157–161. [Google Scholar]
- Cardé, R.T. Defining attraction and aggregation pheromones: Teleological versus functional perspective. J. Chem. Ecol. 2014, 40, 519–520. [Google Scholar] [CrossRef] [PubMed]
- Kosi, A.Ž.; Zou, Y.; Hoskovec, M.; Vrezec, A.; Stritih, N.; Millar, J.G. Novel, male-produced aggregation pheromone of the cerambycid beetle Rosalia alpina, a priority species of European conservation concern. PLoS ONE 2017, 12, e0183279. [Google Scholar]
- Hirano, M.; Higashijima, K.; Koga, Y.; Ochi, T. Distributional expansion of Rosalia batesi in central Kinki district, Japan. Gekkan-Mushi 2017, 556, 44–47. [Google Scholar]
- Fukaya, M.; Yasuda, T.; Akino, T.; Yasui, H.; Wakamura, S.; Fukada, T.; Ogawa, Y. Effects of male body size on mating behavior and female mate refusal in the white–spotted longicorn beetle, Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2004, 39, 731–737. [Google Scholar] [CrossRef]
- Michelsen, A. Observations on the sexual behavior of some longicorn beetles, subfamily Lepturinae (Coleoptera, Cerambycidae). Behaviour 1963, 22, 152–166. [Google Scholar] [CrossRef]
- Michelsen, A. On the evolution of tactile stimulatory actions in longhorned beetles (Coleoptera, Cerambycidae). Z. Tierpsychol. 1966, 23, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, A. The sexual behaviour of some longhorned beetles (Col., Cerambycidae). Entomol. Medd. 1966, 34, 329–355. [Google Scholar]
- Goldsmith, S.K.; Stewart, Z.; Adams, S.; Trimble, A. Body size, male aggression, and male mating success in the cottonwood borer, Plectrodera scalator (Coleoptera: Cerambycidae). J. Insect Behav. 1996, 9, 719–727. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Santos, J.C. Feeding and mating behavior of Dorcacerus barbatus (Olivier, 1790) (Coleoptera: Cerambycidae) on Lantana camara L. (Verbenaceae). Lundiana 2009, 9, 81–84. [Google Scholar]
- Lacey, E.S.; Millar, J.G.; Moreira, J.A.; Hanks, L.M. Male-produced aggregation pheromones of the cerambycid beetles Xylotrechus colonus and Sarosesthes fulminans. J. Chem. Ecol. 2009, 35, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Nishio, Y. On the fan-like cutinized organ on the 8th abdominal sternite of the male of Cermabycidae (In Japanese). Akitu 1959, 8, 67–71. [Google Scholar]
- Nishio, Y. On the 8th abdominal tergite of the male of some cerambycid-beetles (In Japanese). Kontyû 1959, 27, 276–283. [Google Scholar]
- Barbour, J.D.; Cervantes, D.E.; Lacey, E.S.; Hanks, L.M. Calling behavior in the primitive longhorned beetle Prionus californicus Mots. J. Insect Behav. 2006, 19, 623–629. [Google Scholar] [CrossRef]
- Iwabuchi, K. Sex pheromones in Cerambycidae (In Japanese). In Environmental Entomology–Behavior, Physiology and Chemical Ecology; Hidaka, T., Matsumoto, T., Eds.; University of Tokyo Press: Tokyo, Japan, 1999; pp. 436–451. [Google Scholar]
- Noldt, U.; Fettköther, R.; Dettner, K. Structure of the sex pheromone-producing prothoracic glands of the male old house borer, Hylotrupes bajulus (L.) (Coleoptera: Cerambycidae). Int. J. Insect Morphol. Embryol. 1995, 24, 223–234. [Google Scholar] [CrossRef]
- Fettköther, R.; Dettner, K.; Schröder, F.; Meyer, H.; Francke, W.; Noldt, U. The male pheromone of the old house borer Hylotrupes bajulus (L.) (Coleoptera: Cerambycidae): Identification and female response. Experientia 1995, 51, 270–277. [Google Scholar] [CrossRef]
- Benham, G.S., Jr.; Farrar, R.J. Notes on the biology of Prionus laticollis (Coleoptera: Cerambycidae). Can. Entomol. 1976, 108, 569–576. [Google Scholar] [CrossRef]
- Lemay, M.A.; Silk, P.J.; Sweeney, J. Calling behavior of Tetropium fuscum (Coleoptera: Cerambycidae: Spondylidinae). Can. Entomol. 2010, 142, 256–260. [Google Scholar] [CrossRef]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae). I. Behavioral sequences and existence of the male sex pheromone. Appl. Entomol. Zool. 1982, 17, 494–500. [Google Scholar] [CrossRef]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae) II. Female recognition by male and the existence of a female sex pheromone. Appl. Entomol. Zool. 1985, 20, 416–423. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination of the Male and the Female a) | Individual No. | Body Length (mm) | Body Length Difference (Male—Female) (mm) | Time required until the First Mounting Took Place (sec) | Copulation Duration (sec) b) | 6th Copulation and so Forth | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | 1st | 2nd | 3rd | 4th | 5th | ||||
| Larger male vs. smaller female | M1 | F1 | 29.5 | 22.1 | 7.4 | 1129 | 155 | 2 | 189 | 208 | 174 | |
| M2 | F2 | 29.3 | 22.8 | 6.4 | 3600 | 96 | 195 | – | ||||
| M3 | F3 | 33.0 | 23.3 | 9.7 | 15 | – | ||||||
| M4 | F4 | 30.2 | 23.9 | 6.3 | 3019 | 180 | – | |||||
| M4 | F5 | 30.2 | 22.0 | 8.1 | 274 | 156 | 162 | 152 | – | |||
| Smaller male vs. smaller female | M5 | F6 | 19.0 | 23.3 | −4.2 | 1287 | 249 | 155 | – | |||
| M6 | F7 | 18.0 | 20.9 | −3.0 | 578 | 214 | 91 | 285 | 33 | 97 | yes | |
| M7 | F8 | 21.8 | 22.8 | −1.1 | 2112 | 110 | 242 | 174 | 290 | 220 | yes | |
| M8 | F9 | 20.9 | 22.0 | −1.2 | 733 | 96 | 23 | 172 | 114 | – | ||
| M9 | F10 | 21.1 | 22.0 | −0.9 | 134 | 221 | 74 | 46 | 98 | 148 | yes | |
| Smaller male vs. larger female | M7 | F11 | 21.8 | 24.8 | −3.0 | 671 | 212 | 15 | 91 | 140 | 55 | yes |
| M10 | F12 | 21.3 | 27.1 | −5.8 | 425 | 47 | 119 | 111 | 107 | 112 | yes | |
| M11 | F13 | 21.1 | 26.3 | −5.2 | 1002 | 91 | 183 | 42 | 114 | 25 | yes | |
| M8 | F13 | 20.9 | 26.3 | −5.5 | 45 | 211 | 37 | 147 | 125 | 208 | ||
| M10 | F13 | 21.3 | 26.3 | −5.0 | 1449 | 121 | 136 | 132 | – | |||
| Larger male vs. larger female | M4 | F13 | 30.2 | 26.3 | 3.8 | 1015 | 57 | 26 | 117 | 213 | 147 | |
| M12 | F14 | 29.6 | 26.8 | 2.8 | 3242 | 166 | 26 | 170 | – | |||
| M13 | F12 | 26.6 | 27.1 | −0.5 | 840 | 174 | – | |||||
| M14 | F15 | 30.2 | 27.5 | 2.7 | 2867 | 122 | 100 | 18 | 105 | – | ||
| M11 | F13 | 28.0 | 26.3 | 1.6 | 1460 | 277 | 112 | 160 | 94 | 91 | yes | |
 |
| Odor-Source Animal | Test Animal | Responses (%) | Total (N) | χ2 Statistic | ||
|---|---|---|---|---|---|---|
| (a) Odor-Source Animal Side | (b) Control Side (Blank) | (c) Inactive | χ2 (d.f. = 1) between (a) and (b) | |||
| male | male | 38 | 52 | 10 | 50 | 1.0889 < χ20.05 = 3.841 |
| female | male | 26 | 40 | 34 | 50 | 1.485 < χ20.05 = 3.841 |
| female | female | 46 | 32 | 22 | 50 | 0.3141 < χ20.05 = 3.841 |
| male | female | 84 | 8 | 8 | 50 | 31.3913 > χ20.001 = 10.828 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiriyama, S.; Iwata, R.; Fukaya, M.; Hoshino, Y.; Yamanaka, Y. Mating Behavior of Rosalia batesi (Coleoptera: Cerambycidae) Is Mediated by Male-Produced Sex Pheromones. Insects 2018, 9, 48. https://doi.org/10.3390/insects9020048
Kiriyama S, Iwata R, Fukaya M, Hoshino Y, Yamanaka Y. Mating Behavior of Rosalia batesi (Coleoptera: Cerambycidae) Is Mediated by Male-Produced Sex Pheromones. Insects. 2018; 9(2):48. https://doi.org/10.3390/insects9020048
Chicago/Turabian StyleKiriyama, Satoshi, Ryûtarô Iwata, Midori Fukaya, Youtaro Hoshino, and Yasuyuki Yamanaka. 2018. "Mating Behavior of Rosalia batesi (Coleoptera: Cerambycidae) Is Mediated by Male-Produced Sex Pheromones" Insects 9, no. 2: 48. https://doi.org/10.3390/insects9020048
APA StyleKiriyama, S., Iwata, R., Fukaya, M., Hoshino, Y., & Yamanaka, Y. (2018). Mating Behavior of Rosalia batesi (Coleoptera: Cerambycidae) Is Mediated by Male-Produced Sex Pheromones. Insects, 9(2), 48. https://doi.org/10.3390/insects9020048