Prediction of Sporulation and Germination by the Spider Mite Pathogenic Fungus Neozygites floridana (Neozygitomycetes: Neozygitales: Neozygitaceae) Based on Temperature, Humidity and Time

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Tetranychus urticae Stock Culture
2.2. Neozygites floridana Isolate
2.3. Cadaver Production
2.4. Experimental Setup
2.4.1. Detached Leaf Assay: Temperature and RH Combinations
2.4.2. Detached Leaf Assay: Minimum Period at Optimal Conditions for Sporulation
2.4.3. Semi-Field Screen House Experiment
2.5. Statistical Analysis
2.5.1. Detached Leaf Assays: Temperature, RH, and Duration
Number of Primary Conidia and Capilliconidia, Total Number of Conidia, and Proportion of Capilliconidia
2.5.2. Semi-Field Screen House Experiment
Analyses for Experiment Considering Duration
Goodness-of-Fit Assessment and Software
3. Results
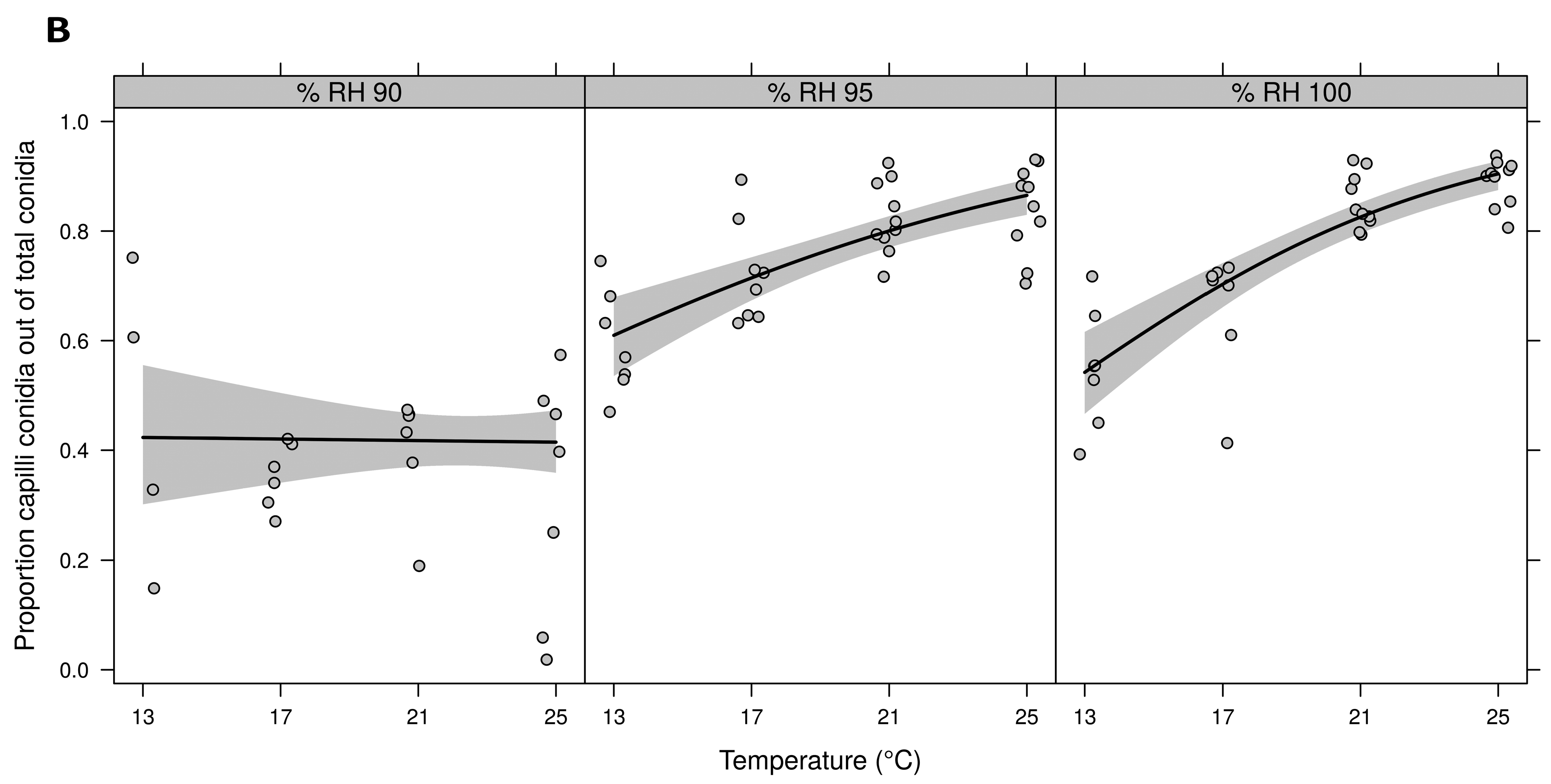
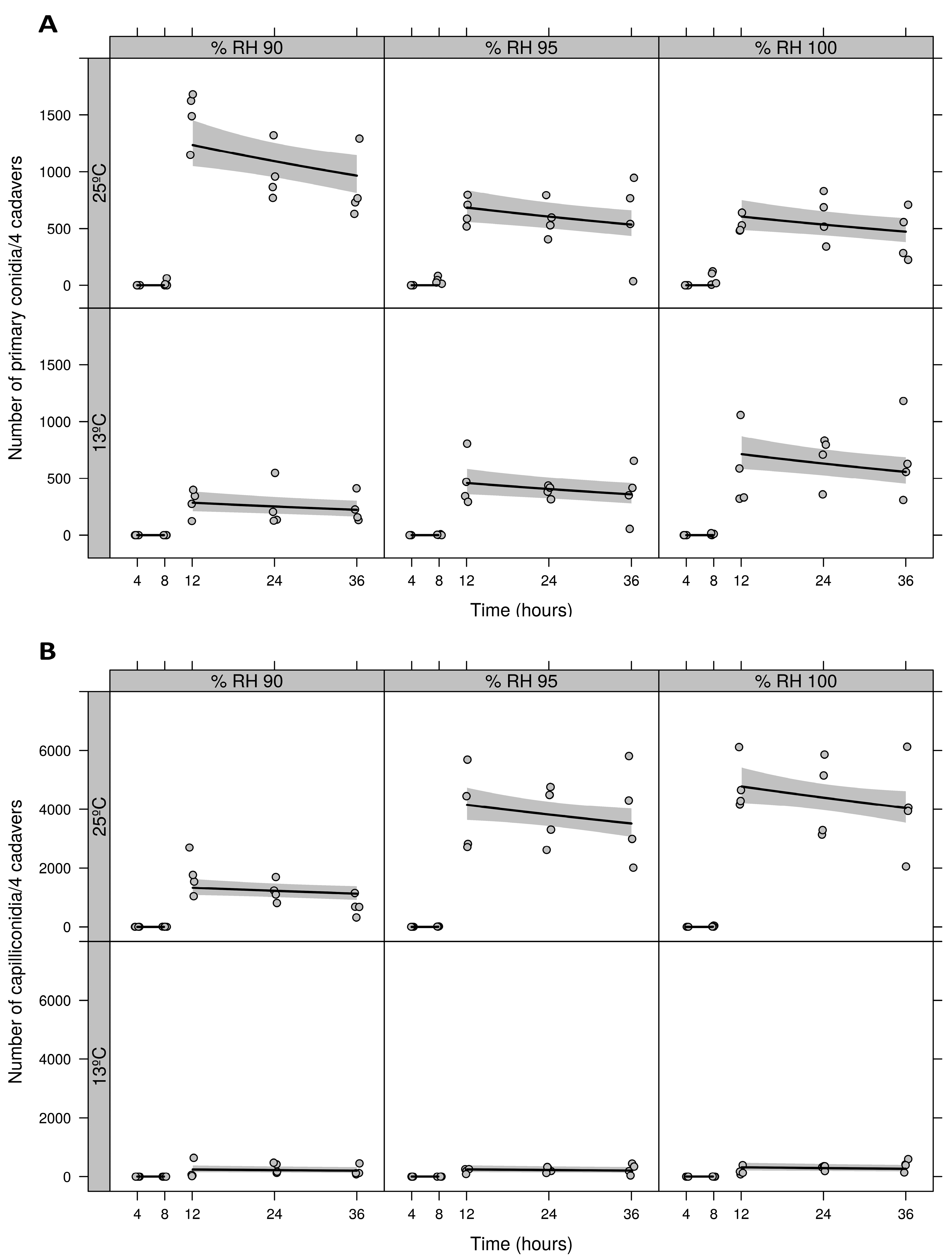
3.1. Detached Leaf Assay: Temperature and RH Combinations
3.2. Detached Leaf Assay: Minimum Period for Optimal RH and Temperature
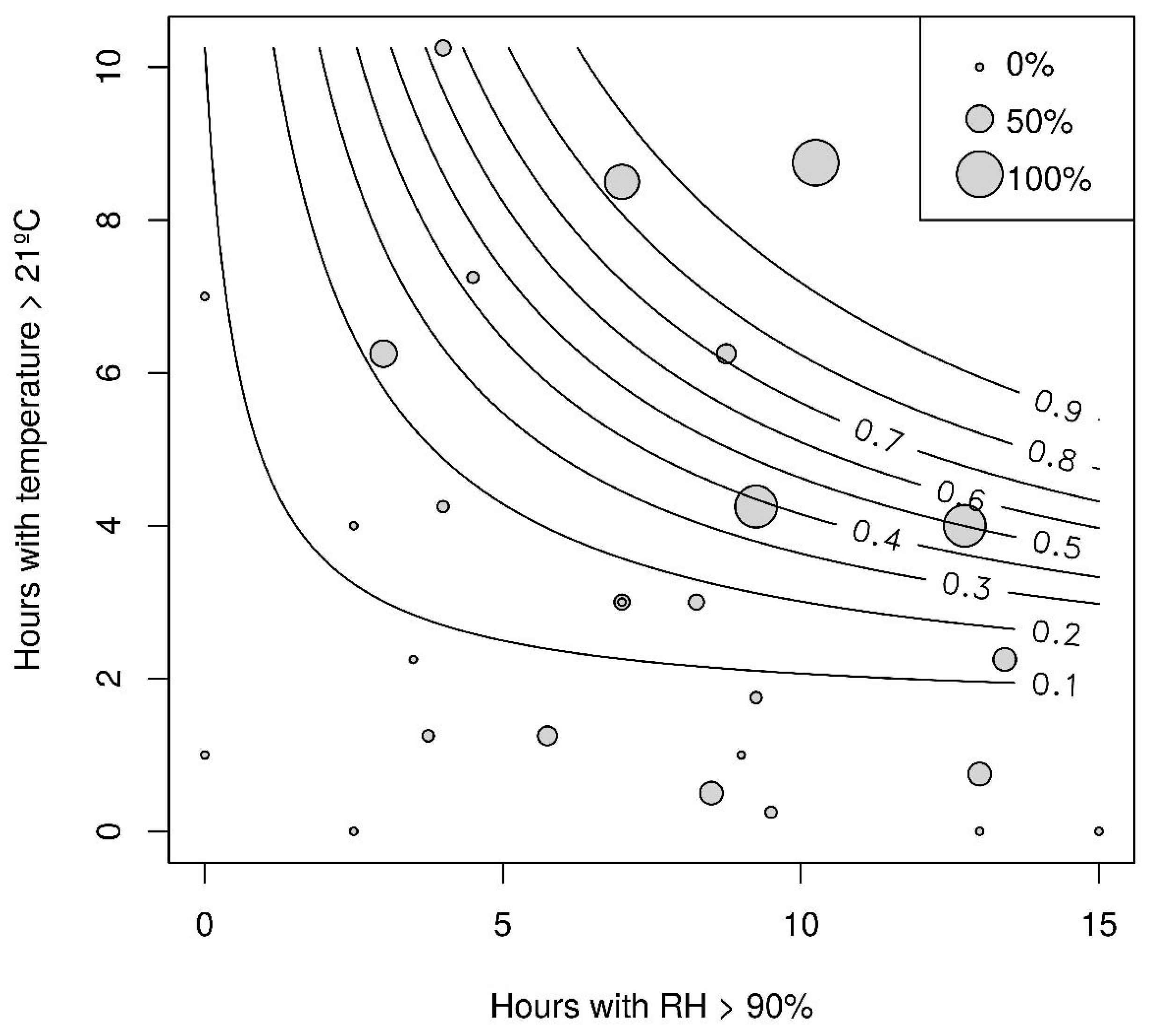
3.3. Semi-Field Screen House Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Humber, R.A. Entomophthoromycota: A new phylum and reclassification for entomophthoroid fungi. Mycotaxon 2012, 120, 477–492. [Google Scholar] [CrossRef]
- Keller, S. Entomophthorales of Switzerland. II. Erynia, Eryniopsis, Neozygites, Zoophthora and Tarichium. Sydowia 1991, 43, 39–112. [Google Scholar]
- Keller, S. The genus Neozygites (Zygomycetes, Entomophthorales) with special reference to species found in tropical regions. Sydowia 1997, 49, 118–146. [Google Scholar]
- Duarte, V.S.; Silva, R.A.; Wekesa, V.W.; Rizzato, F.B.; Dias, C.T.S.; Delalibera, I., Jr. Impact of natural epizootics of the fungal pathogen Neozygites floridana (zygomycetes: Entomophthorales) on population dynamics of Tetranychus evansi (acari: Tetranychidae) in tomato and nightshade. Biol. Control 2009, 51, 81–90. [Google Scholar] [CrossRef]
- Humber, R.A.; Moraes, G.J.; Dossantos, J.M. Natural infection of Tetranychus evansi [acarina, tetranychidae] by a triplosporium sp. [zygomycetes, entomophthorales] in northeastern brazil. Entomophaga 1981, 26, 421–425. [Google Scholar] [CrossRef]
- Klubertanz, T.H.; Pedigo, L.P.; Carlson, R.E. Impact of fungal epizootics on the biology and management of the twospotted spider mite (acari: Tetranychidae) in soybean. Environ. Entomol. 1991, 20, 731–735. [Google Scholar] [CrossRef]
- Boykin, L.S.; Campbell, W.V.; Beute, M.K. Effect of pesticides on Neozygites floridana (entomophthorales: Entomophthoraceae) and arthropod predators attacking the twospotted spider mite (acari: Tetranychidae) in north-carolina peanut fields. J. Econ. Entomol. 1984, 77, 969–975. [Google Scholar] [CrossRef]
- Carner, G.R.; Canerday, T.D. Entomophthora sp. As a factor in regulation of twospotted spider mite in cotton. J. Econ. Entomol. 1970, 63, 638–640. [Google Scholar] [CrossRef]
- Smitley, D.R.; Kennedy, G.G.; Brooks, W.M. Role of the entomogenous fungus, Neozygites floridana, in population declines of the twospotted spider mite, Tetranychus urticae, on field corn. Entomol. Exp. Appl. 1986, 41, 255–264. [Google Scholar] [CrossRef]
- Mietkiewski, R.; Balazy, S.; Van der Geest, L.P.S. Observations on a mycosis of spider-mites (acari, tetranychidae) caused by Neozygites floridana in poland. J. Invertebr. Pathol. 1993, 61, 317–319. [Google Scholar] [CrossRef]
- Nordengen, I.; Klingen, I. Comparison of methods for estimating the prevalence of Neozygites floridana in Tetranychus urticae populations infesting strawberries. J. Invertebr. Pathol. 2006, 92, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Van der Geest, L.P.; Elliot, S.L.; Breeuwer, J.A.; Beerling, E.A. Diseases of mites. Exp. Appl. Acarol. 2000, 24, 497–560. [Google Scholar] [CrossRef] [PubMed]
- Delalibera, I., Jr.; Hajek, A.E.; Humber, R.A. Use of cell culture media for cultivation of the mite pathogenic fungi Neozygites tanajoae and Neozygites floridana. J. Invertebr. Pathol. 2003, 84, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, G.G.; Smitley, D.R. Method of Controlling Plant Feeding Mites with the Fungus Neozygites floridana. U.S. Patent No 4,752,468, 21 June 1988. [Google Scholar]
- Carner, G.R. A description of the life cycle of Entomophthora sp. in the two-spotted spider mite. J. Invertebr. Pathol. 1976, 28, 245–254. [Google Scholar] [CrossRef]
- Delalibera, I., Jr.; Demétrio, C.G.B.; Manly, B.F.J.; Hajek, A.E. Effect of relative humidity and origin of isolates of Neozygites tanajoae (zygomycetes: Entomophthorales) on production of conidia from cassava green mite, Mononychellus tanajoa (acari: Tetranychidae), cadavers. Biol. Control 2006, 39, 489–496. [Google Scholar] [CrossRef]
- Elliot, S.L. Ecology and Epizootiology of Neozygite floridana, a Pathogen of the Cassava Green Mite; Imperial College: London, UK, 1998. [Google Scholar]
- Pell, J.K.; Eilenberg, J.; Hajek, A.E.; Steinkraus, D.C. Biology, ecology and pest management potential of Entomophthorales. In Fungi as Biocontrol Agents. Progress, Problems and Potential; Butt, T.M., Jackson, C.W., Magan, N., Eds.; CABII Publishing: Oxon, UK, 2001; pp. 71–153. ISBN 0-85199-356-7. [Google Scholar]
- De Castro, T.R.; Wekesa, V.W.; Moral, R.D.; Demetrio, C.G.B.; Delalibera, I.; Klingen, I. The effects of photoperiod and light intensity on the sporulation of brazilian and norwegian isolates of Neozygites floridana. J. Invertebr. Pathol. 2013, 114, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Klingen, I.; Holthe, M.P.; Westrum, K.; Suthaparan, A.; Torp, T. Effect of light quality and light-dark cycle on sporulation patterns of the mite pathogenic fungus Neozygites floridana (Neozygitales: Entomophthoromycota), a natural enemy of Tetranychus urticae. J. Invertebr. Pathol. 2016, 137, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Klingen, I.; Wærsted, G.; Westrum, K. Overwintering and prevalence of Neozygites floridana (Zygomycetes: Entomophthorales) in hibernating females of Tetranychus urticae (Acari: Tetranychidae) under cold climatic conditions in strawberries. Exp. Appl. Acarol. 2008, 46, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Duarte, V.S.; Westrum, K.; Ribeiro, A.E.L.; Gondim, M.G.C., Jr.; Klingen, I.; Delalibera, I., Jr. Abiotic and biotic factors affecting resting spore formation in the mite pathogen Neozygites floridana. Int. J. Microbiol. 2013, 2013, 276168. [Google Scholar] [CrossRef]
- Wekesa, V.W.; Moraes, G.J.; Ortega, E.M.M.; Delalibera, I., Jr. Effect of temperature on sporulation of Neozygites floridana isolates from different climates and their virulence against the tomato red spider mite, Tetranychus evansi. J. Invertebr. Pathol. 2010, 103, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Bruck, D.J.; Lacey, L.A. Chapter 3—Principles of epizootiology and microbial control. In Insect Pathology, 2nd ed.; Kaya, H.K., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 29–72. ISBN 978-0-12-384984-7. [Google Scholar]
- Brown, G.C.; Hasibuan, R. Conidial discharge and transmission efficiency of Neozygites floridana, an entomopathogenic fungus infecting 2-spotted spider-mites under laboratory conditions. J. Invertebr. Pathol. 1995, 65, 10–16. [Google Scholar] [CrossRef]
- Oduor, G.I.; Yaninek, J.S.; van der Geest, L.P.S.; de Moraes, G.J. Germination and viability of capilliconidia of Neozygites floridana (zygomycetes: Entomophthorales) under constant temperature, humidity, and light conditions. J. Invertebr. Pathol. 1996, 67, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Klingen, I.; Nilsen, S.S. Mechanisms important for the epidemic development of Neozygites floridana in Tetranychus urticae. IOBC WPRS Bull. 2009, 45, 287–289. [Google Scholar]
- Boulard, T.; Fatnassi, H.; Roy, J.C.; Lagier, J.; Fargues, J.; Smits, N.; Rougier, M.; Jeannequin, B. Effect of greenhouse ventilation on humidity of inside air and in leaf boundary-layer. Agric. For. Meteorol. 2004, 125, 225–239. [Google Scholar] [CrossRef]
- Fargues, J.; Vidal, C.; Smits, N.; Rougier, M.; Boulard, T.; Mermier, M.; Nicot, P.; Reich, P.; Jeannequin, B.; Ridray, G.; et al. Climatic factors on entomopathogenic hyphomycetes infection of Trialeurodes vaporariorum (homoptera: Aleyrodidae) in mediterranean glasshouse tomato. Biol. Control 2003, 28, 320–331. [Google Scholar] [CrossRef]
- Duarte, V.S. Studies for Implementation of Neozygites floridana as Control Agent of Two-Spotted Spider Mite in Strawberry; Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo: São Paulo, Brazil, 2013. [Google Scholar]
- Solomon, M.E. Control of humidity with potassium hydroxide, sulphuric acid, or other solutions. Bull. Entomol. Res. 1951, 42, 543–554. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman and Halln: London, UK, 1989; 511p. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing, version 3.2.2; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 5 February 2017).
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-normal plots and overdispersed models in R: The hnp package. J. Stat. Softw. 2017, 81, 23. [Google Scholar] [CrossRef]
- Smitley, D.R.; Brooks, W.M.; Kennedy, G.G. Environmental effects on production of primary and secondary conidia, infection, and pathogenesis of Neozygites floridana, a pathogen of the twospotted spider mite, Tetranychus urticae. J. Invertebr. Pathol. 1986, 47, 325–332. [Google Scholar] [CrossRef]
- Oduor, G.I.; de Moraes, G.J.; Van der Geest, L.P.S.; Yaninek, J.S. Production and germination of primary conidia of Neozygites floridana (zygomycetes: Entomophthorales) under constant temperatures, humidities, and photoperiods. J. Invertebr. Pathol. 1996, 68, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Oduor, G.I.; de Moraes, G.J.; Yaninek, J.S.; van der Geest, L.P.S. Effect of temperature, humidity and photoperiod on mortality of Mononychellus tanajoa (acari: Tetranychidae) infected by Neozygites cf floridana (zygomycetes: Entomophthorales). Exp. Appl. Acarol. 1995, 19, 571–579. [Google Scholar] [CrossRef]
- Boulard, T.; Mermier, M.; Fargues, J.; Smits, N.; Rougier, M.; Roy, J.C. Tomato leaf boundary layer climate: Implications for microbiological whitefly control in greenhouses. Agric. For. Meteorol. 2002, 110, 159–176. [Google Scholar] [CrossRef]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, T.; Moral, R.D.A.; Demétrio, C.G.B.; Delalibera, I.; Klingen, I. Prediction of Sporulation and Germination by the Spider Mite Pathogenic Fungus Neozygites floridana (Neozygitomycetes: Neozygitales: Neozygitaceae) Based on Temperature, Humidity and Time. Insects 2018, 9, 69. https://doi.org/10.3390/insects9020069
Castro T, Moral RDA, Demétrio CGB, Delalibera I, Klingen I. Prediction of Sporulation and Germination by the Spider Mite Pathogenic Fungus Neozygites floridana (Neozygitomycetes: Neozygitales: Neozygitaceae) Based on Temperature, Humidity and Time. Insects. 2018; 9(2):69. https://doi.org/10.3390/insects9020069
Chicago/Turabian StyleCastro, Thiago, Rafael De Andrade Moral, Clarice Garcia Borges Demétrio, Italo Delalibera, and Ingeborg Klingen. 2018. "Prediction of Sporulation and Germination by the Spider Mite Pathogenic Fungus Neozygites floridana (Neozygitomycetes: Neozygitales: Neozygitaceae) Based on Temperature, Humidity and Time" Insects 9, no. 2: 69. https://doi.org/10.3390/insects9020069
APA StyleCastro, T., Moral, R. D. A., Demétrio, C. G. B., Delalibera, I., & Klingen, I. (2018). Prediction of Sporulation and Germination by the Spider Mite Pathogenic Fungus Neozygites floridana (Neozygitomycetes: Neozygitales: Neozygitaceae) Based on Temperature, Humidity and Time. Insects, 9(2), 69. https://doi.org/10.3390/insects9020069