Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to Soil-Borne Entomopathogenic Fungi

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Insects
2.2. Production of Fungal Conidial Suspensions
2.3. Pathogenicity Bioassay
2.4. Virulence Bioassay and Determination of LC50
2.5. Greenhouse Assay
2.6. Statistical Analyses
3. Results
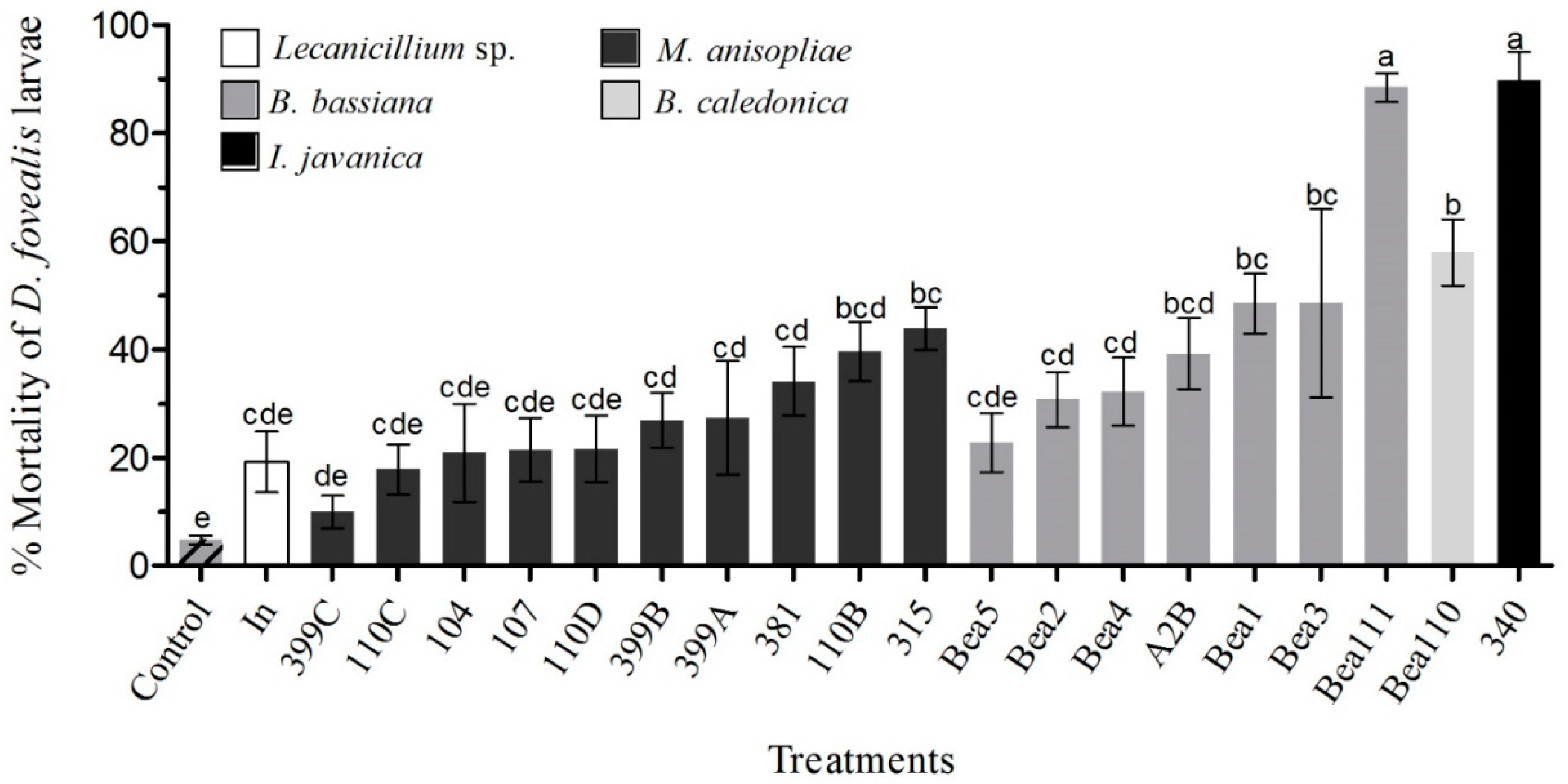
3.1. Pathogenicity Bioassay
3.2. Virulence Bioassay
3.3. Greenhouse Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franco, C.M.; Baptista, M. Duponchelia fovealis Zeller-nova praga em Portugal. Revista Frutas, Legumes e Flores 2010, 110, 34–35. [Google Scholar]
- Bonsignore, C.P.; Vacante, V. Emerging insects and pests in southern Italy. Protezione delle Colture 2009, 4, 25–31. [Google Scholar]
- Efil, E.; Özgür, O.; Efıl, F. New pest, Duponchelia fovealis Zeller on strawberries in Turkey—Damage, distribution and parasitoid. J. Entomol. Zool. Stud. 2014, 2, 328–334. [Google Scholar]
- Zawadneak, M.A.C.; Gonçalves, R.B.; Pimentel, I.C.; Schuber, J.M.; Santos, B.; Poltronieri, A.S.; Solis, M.A. First record of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) in South America. Idesia 2016, 34, 89–93. [Google Scholar] [CrossRef]
- CABI International. Available online: http://www.cabi.org/isc/datasheet/20168 (accessed on 14 March 2018).
- Stocks, S.D.; Hodges, A. A European Pepper Moth or Southern European Marsh Pyralid Duponchelia fovealis (Zeller). 2014. Available online: http://edis.ifas.ufl.edu/pdffiles/IN/IN91000.pdf (accessed on 20 August 2017).
- Zawadneak, M.A.C.; Botton, M.; Schuber, J.M.; Santos, B.; Vidal, H.R. Pragas do morangueiro. In Como Produzir Morangos, 1st ed.; Zawadneak, M.A.C., Schuber, J.M., Mógor, A.F., Eds.; Editora UFPR: Curitiba, Brazil, 2014; pp. 101–145. [Google Scholar]
- Rodrigues, R.; Jaras, L.I.; Poltronieri, A.S.; Pimentel, I.C.; Zawadneak, M.A.C. Seletividade de inseticidas reguladores de crescimento e botânico no parasitismo de três espécies de Trichogramma em ovos de Duponchelia fovealis Zeller (Lepidoptera: Crambidae). EntomoBrasilis 2017, 10, 26–32. [Google Scholar] [CrossRef]
- Amatuzzi, R.F.; Cardoso, N.; Poltronieri, A.S.; Poitevin, C.G.; Dalzoto, P.; Zawadeneak, M.A.; Pimentel, I.C. Potential of endophytic fungi as biocontrol agents of Duponchelia fovealis (Zeller) (Lepidoptera: Crambidae). Braz. J. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Zimmermann, G. Mycopathogens of Soil Insects, 1st ed.; Academic Press: London, UK, 1989; pp. 240–270. [Google Scholar]
- Gonzalez, F.; Tkaczuk, C.; Dinu, M.M.; Fiedler, Z.; Vidal, S.; Zchori-Fein, E.; Messelink, G.J. New opportunities for the integration of microorganisms into biological pest control systems in greenhouse crops. J. Pest Sci. 2016, 89, 295–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, C.W. Entomogenous fungi as microbial pesticides. In New Directions in Biological Control; Baker, R.R., Dunn, P.E., Alan, R.L., Eds.; Wiley-Liss: New York, NY, USA, 1990; pp. 139–159. [Google Scholar]
- Roberts, D.W.; Leger, R.J. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.B.; Lopes, R.B.; Vieira, A.S.; Tamai, M.A. Fungos entomopatogênicos usados no controle de pragas na América Latina. In Controle Microbiano de Pragas na América Latina—Avanços e Desafios, 1st ed.; Alves, S.B., Lopes, R.B., Eds.; Fundação de Estudos Agrários Luiz de Queiroz: Piracicaba, Brazil, 2008; pp. 69–110. [Google Scholar]
- Ortiz-Urquiza, A.; Luo, Z.; Keyhani, N.O. Improving mycoinsecticides for insect biological control. Appl. Microbiol. Biotechnol. 2015, 99, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A. Microbial Control of Insect and Mite Pests from Theory to Practice, 1st ed.; Academic Press: London, UK, 2016; p. 482. [Google Scholar]
- Thomas, S.R.; Elkinton, J.S. Pathogenicity and virulence. J. Invertebr. Pathol. 2004, 85, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Zawadneak, M.A.C.; Gonçalves, R.B.; Poltronieri, A.S.; Santos, B.; Bischoff, A.M.; Borba, A.M.; Pimentel, I.C. Biological parameters of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) reared in the laboratory on two diets. Eur. J. Entomol. 2017, 114, 291–294. [Google Scholar] [CrossRef]
- Alves, S.B.; Moraes, S.A. Quantificação de inoculo de patógenos de insetos. In Controle Microbiano de Inseto, 2nd ed.; Alves, S.B., Ed.; Piracicaba Fundação de Estudos Agrários Luiz de Queiroz: Piracicaba, Brazil, 1998; Volume 1, pp. 765–778. [Google Scholar]
- Goettel, M.S.; Inglis, G.D. Fungi: Hyphomycetes. In Manual of Techiniques in Insect Pathology, 1st ed.; Lacey, L.A., Ed.; Academic Press: London, UK, 1997; pp. 213–250. [Google Scholar]
- Peixoto, N.P.A.S.; Azevedo, J.L.; Araújo, W.L. Microrganismos Endofíticos Revista Biotecnologia Biotecnologia; Ciência & Desenvolvimento: Brasília, Brazil, 2002; pp. 62–76. [Google Scholar]
- Abbott, W.S.A. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 58, 265–267. [Google Scholar] [CrossRef]
- LeOra Software. Polo Plus Probit and Logit Analysis; Version 1.0; LeOra Software: Berkeley, CA, USA, 2002. [Google Scholar]
- Henderson, C.F.; Tilton, E.W. Tests with acaricides against the brown wheat mite. J. Econ. Entomol. 1955, 48, 157–161. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E.; Avery, P.B.; Jaronski, S.T.; Vandenberg, J.D. Comparative virulence of Beauveria bassiana isolates against lepidopteran pests of vegetable crops. J. Invertebr. Pathol. 2010, 103, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Batta, Y.S. Efficacy of endophytic and applied Metarhizium anisopliae (Metch.) Sorokin (Ascomycota: Hypocreales) against larvae of Plutella xylostella L. (Yponomeutidae: Lepidoptera) infesting Brassica napus plants. Crop Prot. 2013, 44, 128–134. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Zhao, Z.; Humber, R.A. Laboratory and field efficacy of entomopathogenic fungi for the management of the sweet potato weevil, Cylas formicarius (Coleoptera: Brentidae). J. Invertebr. Pathol. 2014, 122, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Castro, T.; Mayerhofer, J.; Enkerli, J.; Eilenberg, J.; Meyling, N.V.; Moral, R.A.; Demétrio, C.G.B.; Delalibera, I. Persistence of Brazilian isolates of the entomopathogenic fungi Metarhizium anisopliae and M. robertsii in strawberry crop soil after soil drench application. Agric. Ecosyst. Environ. 2016, 233, 361–369. [Google Scholar] [CrossRef]
- Godonou, I.; Jamesa, B.; Atcha-Ahowéa, C.; Vodouhèb, S.; Kooymanc, C.; Ahanchédéb, A.; Koriea, S. Potential of Beauveria bassiana and Metarhizium anisopliae isolates from Benin to control Plutella xylostella L. (Lepidoptera: Plutellidae). Crop. Prot. 2009, 28, 220–224. [Google Scholar] [CrossRef]
- Glare, T.R.; Reay, S.D.; Nelson, T.L.; Moore, R. Beauveria caledonica is a naturally occurring pathogen of forest beetles. Mycol. Res. 2008, 112, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Ahmed, S. Entomopathogenic fungi disturbed the larval growth and feeding performance of Ocinara varians Walker (Lepidoptera: Bombycidae) larvae. Insect Sci. 2009, 16, 511–517. [Google Scholar] [CrossRef]
- Nussenbaum, A.L.; Lecuona, R.E. Selection of Beauveria bassiana sensu lato and Metarhizium anisopliae sensu lato isolates as microbial control agents against the boll weevil (Anthonomus grandis) in Argentina. J. Invertebr. Pathol. 2012, 110, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schrank, A.; Vainstein, M.H. Metarhizium anisopliae enzymes and toxins. Toxicon 2010, 56, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Maranhao, E.A.A.; Valverde-García, P.; Santiago-Álvarez, C. Selection of Beauveria bassiana isolates for control of the whiteflies Bemisia tabaci and Trialeurodes vaporariorum on the basis of their virulence, thermal requirements and toxicogenic activity. Biol. Control 2006, 36, 274–287. [Google Scholar] [CrossRef]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Huang, Z.; Ali, S.; Ren, S.X.; Wu, J.H. Effect of Isaria fumosoroseus on mortality and fecundity of Bemisia tabaci and Plutella xylostella. Insect Sci. 2010, 17, 140–148. [Google Scholar] [CrossRef]
- Cabanillas, H.E.; Jones, W.A. Pathogenicity of Isaria poprawskii (Ascomycota: Hypocreales: Cordycipitaceae) against the glassy-winged sharpshooter, Homalodisca vitripennis (Hemiptera: Cicadellidae), under laboratory conditions. Crop Prot. 2013, 50, 46–52. [Google Scholar] [CrossRef]
- Shipp, J.L.; Zhang, Y.; Hunt, D.W.A.; Ferguson, G. Influence of humidity and greenhouse microclimate on the efficacy of Beauveria bassiana (Balsamo) for control of greenhouse arthropod pests. Environ. Entomol. 2003, 32, 1154–1163. [Google Scholar] [CrossRef]
- Senthil, K.C.M.; Jacob, T.K.; Devasahayam, S.; D’Silva, S.; Nandeesh, P.G. Characterization and virulence of Beauveria bassiana associated with auger beetle (Sinoxylon anale) infecting allspice (Pimenta dioica). J. Invertebr. Pathol. 2016, 139, 67–73. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Species | Isolate | GenBank | Soil Source | Coordinates and Sites |
|---|---|---|---|---|
| Beauveria bassiana | Bea1 | KY471,648 | Conventional corn (Zea mays) crop systems | 25°53′190″ S, 49°43′195″ W Araucária |
| Bea2 | KY471,649 | |||
| Bea3 | KY471,650 | |||
| Bea4 | KY471,651 | |||
| Bea5 | KY471,652 | |||
| Beauveria caledonica | Bea110 | KY471,655 | Conventional strawberry (Fragaria × ananassa) crop systems | 25°53′376″ S, 49°44′654″ W Araucária |
| Beauveria bassiana | Bea111 | KY471,653 | Conventional corn (Zea mays) crop systems | |
| A2B | KY471,654 | |||
| Lecanicillium sp. | In1 | KY471,666 | Organic strawberry (Fragaria × ananassa) crop systems | 25°74′010″ S, 49°89′425″ W Lapa |
| Metarhizium anisopliae | 104 | KY471,656 | Native forest | 25°34′154″ S, 48°90′083″ W Morretes |
| 107 | KY471,657 | |||
| Isaria javanica | Isa340 | KY488,507 | ||
| Metarhizium anisopliae | 110B | KY471,658 | Banana trees (Musa spp.) in native forest | 25°38′776″ S, 48°86′026″ W Morretes |
| 110C | KY471,659 | |||
| 110D | KY471,660 | |||
| 315 | KY471,661 | |||
| 381 | KY471,662 | |||
| 399A | KY471,663 | |||
| 399B | KY471,664 | |||
| 399C | KY471,665 |
| Treatment | Bea1 | Bea110 | Bea111 | Isa340 | CG716 |
|---|---|---|---|---|---|
| Number of larvae | 601 | 557 | 609 | 645 | 432 |
| LC50 | 9.5 × 108 | 3.85 × 108 | 2.33 × 106 | 9.69 × 105 | 1.41 × 107 |
| Slope (SE) | 0.24 ± 0.06 | 0.25 ± 0.05 | 0.51 ± 0.04 | 0.37 ± 0.03 | 0.21 ± 0.05 |
| 95% FL | 3.7 × 108–5.09 × 109 | 5.13 × 107–5.6 × 109 | 8.88 × 105–5.34 × 106 | 5.79 × 105–3.12 × 106 | 4.48 × 106–8.38 × 107 |
| χ2 (df = 4) | 2.56 | 1.27 | 3.65 | 2.77 | 0.29 |
| LT50 (Days) | 7.1 | 5.3 | 7.2 | 6.4 | 7.5 |
| Slope (SE) 95% FL | 2.10 ± 0.27 6.0 to 9.0 | 2.16 ± 0.25 4.6 to 6.2 | 3.13 ± 0.38 6.4 to 8.6 | 2.81 ± 0.30 5.4 to 8.3 | 2.75 ± 0.39 6.4 to 9.6 |
| Treatment | N° Initial Larvae | % Efficacy (SE) |
|---|---|---|
| Bea111 (2 × 106) * | 197 | 52.68 (6.23) a |
| Isa340 (9 × 105) * | 196 | 45.83 (7.79) a |
| Control | 199 | 17.41 (3.11) b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amatuzzi, R.F.; Poitevin, C.G.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to Soil-Borne Entomopathogenic Fungi. Insects 2018, 9, 70. https://doi.org/10.3390/insects9020070
Amatuzzi RF, Poitevin CG, Poltronieri AS, Zawadneak MAC, Pimentel IC. Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to Soil-Borne Entomopathogenic Fungi. Insects. 2018; 9(2):70. https://doi.org/10.3390/insects9020070
Chicago/Turabian StyleAmatuzzi, Rafaela F., Carolina G. Poitevin, Alex S. Poltronieri, Maria A. C. Zawadneak, and Ida C. Pimentel. 2018. "Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to Soil-Borne Entomopathogenic Fungi" Insects 9, no. 2: 70. https://doi.org/10.3390/insects9020070
APA StyleAmatuzzi, R. F., Poitevin, C. G., Poltronieri, A. S., Zawadneak, M. A. C., & Pimentel, I. C. (2018). Susceptibility of Duponchelia fovealis Zeller (Lepidoptera: Crambidae) to Soil-Borne Entomopathogenic Fungi. Insects, 9(2), 70. https://doi.org/10.3390/insects9020070