Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Strains
2.2. Mosquito Rearing
2.3. Synthetic Pyrethroids, Detoxification Enzyme Inhibitors, and Plant Essential Oils
2.4. Insecticide Susceptibility Testing
2.5. Enhancement of Insecticides Applied in Concert with Plant Oils
2.6. Synergism of Pyrethroids by Plant Essential Oils
2.7. Inhibition of Detoxification Enzymes
3. Results/Discussion
3.1. Enhancement/Synergism
3.2. Inhibition of Detoxification Enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Ann. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R. Insecticide resistance in disease vectors of public health importance. Pest Manag. Sci. 2007, 63, 628–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Global Plan for Insecticide Resistance Management in Malaria Vectors. 2012. Available online: http://www.who.int/malaria/vector_control/gpirm_executive_summary_en.pdf?ua=1 (accessed on 12 July 2018).
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). A Global Brief on Vector-Borne Diseases. 2014. Available online: http://apps.who.int/iris/bitstream/10665/111008/1/WHO_DCO_WHD_2014.1_eng.pdf (accessed on 7 July 2018).
- Narahashi, T. Effect of the insecticide allethrin on membrane potentials of cockroach giant axons. J. Cell. Comp. Physiol. 1962, 59, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Narahashi, T. Effects of Insecticides on Excitable Tissues. Adv. Insect Phys. 1972, 8, 1–93. [Google Scholar]
- Norris, E.; Coats, J. Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. Int. J. Environ. Res. Public Health 2017, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Mount, G.A. A critical review of ultra low-volume aerosols of insecticide applied with vehicle mounted generators for adult mosquito control. J. Am. Mosq. Control. Assoc. 1998, 14, 305–334. [Google Scholar] [PubMed]
- Mount, G.A.; Biery, T.L.; Haile, D.G. A review of ultralow-volume aerial sprays of insecticide for mosquito control. J. Am. Mosq. Control Assoc. 1996, 12, 601–618. [Google Scholar] [PubMed]
- World Health Organization (WHO). Guidelines for Laboratory and Field Testing of Long-Lasting Insecticidal Mosquito Nets; WHO/CDS/WHOPES/GDCPP/2005.11; WHO: Geneva, Switzerland, 2005. [Google Scholar]
- Tengena, J.A.; Adiamoh, M.; D’Alessandro, U.; Larju, L.; Jawara, M.; Jeffries, D.; Malik, N.; Nwakanma, D.; Kaur, H.; Takken, W.; et al. Alternative treatments for indoor residual spraying for malaria control in a village with pyrethroid- and DDT-resistant vectors in the Gambia. PLoS ONE 2013, 8, e74351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agramonte, N.; Bloomquist, J.; Bernier, U.; Apperson, C. Pyrethroid resistance alters the blood-feeding behavior in Puerto Rican Aedes aegypti mosquitoes exposed to treated fabric. PLoS Negl. Trop. Dis. 2017, 11, e0005954. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Insecticide Resistance and Vector Control. 1970. Available online: http://apps.who.int/iris/bitstream/10665/40771/1/WHO_TRS_443_%28part1%29.pdf (accessed on 2 July 2018).
- Smith, L.; Kasai, S.; Scott, J. Voltage-sensitive sodium channel mutations S989P V1016G in Aedes aegypti confer variable resistance to pyrethroids, DDT and oxadiazines. Pest Manag. Sci. 2018, 74, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Namountougou, M.; Diabaté, A.; Etang, J.; Bass, C.; Sawadogo, S.P.; Gnankinié, O.; Baldet, T.; Martin, T.; Chandre, F.; Simard, F.; Dabiré, R.K. First report of the L1014S kdr mutation in wild populations of Anopheles gambiae M and S molecular forms in Burkina Faso (West Africa). Acta Trop. 2013, 125, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.; Hammock, F. Genetic and molecular evidence for a trans-acting regulatory locus controlling glutathione S-transferase-2 expression in Aedes aegypti. Mol. Gen. Genet. 1992, 234, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Molecular biology of insecticide resistance. Toxicol. Lett. 1995, 82, 83–90. [Google Scholar] [CrossRef]
- Berge, J.; Feyereisen, R.; Amichot, M. Cytochrome P450 Monooxygenases and Insecticide Resistance in Insects. Phil. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Jin, T.; Zeng, L.; Lu, Y. Cuticular penetration of β-cypermethrin in insecticide-susceptible and resistant strains of Bactrocera dorsalis. Pestic. Biochem. Physiol. 2012, 103, 189–193. [Google Scholar] [CrossRef]
- Takahashi, O.; Oishi, S.; Fujitani, T.; Tanaka, T.; Yoneyama, M. Chronic toxicity studies of piperonyl butoxide in CD-1 mice: Induction of hepatocellular carcinoma 1. Toxicology 1997, 124, 95–103. [Google Scholar] [CrossRef]
- Tanaka, T.; Fujitani, T.; Takahashi, O.; Oishi, S. Developmental toxicity evaluation of piperonyl butoxide in CD-1 mice. Toxicology Lett. 1994, 71, 123–129. [Google Scholar] [CrossRef]
- Fujitani, T.; Ando, H.; Fujitani, K.; Ikeda, T.; Kojima, A.; Kubo, Y.; Ogata, A.; Oishi, S.; Takahashi, H.; Takahashi, O.; et al. Sub-acute toxicity of piperonyl butoxide in F344 rats. Toxicology 1992, 72, 291–298. [Google Scholar] [CrossRef]
- Gross, A.D.; Norris, E.J.; Kimber, M.J.; Bartholomay, L.C.; Coats, J.R. Essential oils enhance the toxicity of permethrin against Aedes aegypti and Anopheles gambiae. Med. Vet. Entomol. 2017, 31, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Estep, A.S.; Sanscrainte, N.D.; Waits, C.M.; Louton, J.E.; Becnel, J.J. Resistance Status and Resistance Mechanisms in a Strain of Aedes aegypti (Diptera: Culicidae) From Puerto Rico. J. Med. Entomol. 2017, 54, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Ponce-García, G.; Del Río-Galvan, S.; Barrera, R.; Saavedra-Rodriguez, K.; Villanueva-Segura, G.F.; Amador, M.; Flores, A.E. Knockdown resistance mutations in Aedes aegypti (Diptera: Culicidae) from Puerto Rico. J. Med. Entomol. 2016, 53, 1410–1414. [Google Scholar] [CrossRef] [PubMed]
- Mutunga, J.M.; Anderson, T.D.; Craft, D.T.; Gross, A.D.; Swale, D.R.; Tong, F.; Wong, D.M.; Carlier, P.R.; Bloomquist, J.R. Carbamate and pyrethroid resistance in the akron strain of Anopheles gambiae. Pestic. Biochem. Phys. 2015, 121, 116–121. [Google Scholar] [CrossRef] [PubMed]
- BEI Resources. Catalog # MRA-913 Anopheles Gambia AKRON 2018. Available online: http://www.beiresources.org/Catalog/livingMosquitoes/MRA-913.aspx accessed 3.7.2018 (accessed on 3 July 2018).
- Reid, W.R.; Thornton, A.; Pridgeon, J.W.; Becnel, J.J.; Tang, F.; Estep, A.; Clark, G.G.; Allen, S.; Liu, N. Transcriptional analysis of four family 4 P450s in a Puerto Rico strain of Aedes aegypti (Diptera: Culicidae) compared with an Orlando strain and their possible functional roles in permethrin resistance. J. Med. Entomol. 2014, 51, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Finney, D. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- World Health Organization (WHO). Test Procedures for Insecticide Resistance Monitoring in Malaria Mosquitoes, 2nd ed.; WHO: Geneva, Switzerland, 2018; Available online: http://apps.who.int/iris/bitstream/handle/10665/250677/9789241511575-eng.pdf?sequence=1 (accessed on 24 June 2018).
- Mansour, N.A.; Eldefrawi, M.E.; Toppozada, A.; Zeid, M. Toxicological studies on the Egyptian cotton leafworm, Prodenia litura. VI. Potentiation and antagonism of organo-phosphorus and carbamate insecticides. J. Econ. Ent. 1966, 59, 307–311. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Techniques to Detect Insecticide Resistance Mechanisms (Field and Laboratory Manual); WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Anderson, T.D.; Zhu, K.Y. Synergistic and antagonistic effects of atrazine on the toxicity of organophosphorodithioate and organophosphorothioate insecticides to Chironomus tentans (Diptera: Chironomidae). Pestic. Biochem. Physiol. 2004, 80, 54–64. [Google Scholar] [CrossRef]
- Norris, E.; Gross, A.; Dunphy, B.; Bessette, S.; Bartholomay, L.C.; Coats, J.R. Comparison of the insecticidal characteristics of commercially available plant essential oils against Aedes aegypti and Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Immaraju, J.A.; Morse, J.G.; Hobza, R.F. Field evaluation of insecticide rotation and mixtures as strategies for citrus thrips (Thysanoptera: Thripidae) resistance management in California. J. Econ. Entomol. 1990, 83, 306–314. [Google Scholar] [CrossRef]
- Scott, J.A. The molecular genetics of resistance: Resistance as a response to stress. Fla. Entomol. 1995, 78, 399–414. [Google Scholar] [CrossRef]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Bloomquist, J.R. Plant essential oils affect the toxicities of carbaryl and permethrin against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 773–790. [Google Scholar] [CrossRef]
- Shaalan, E.A.S.; Canyon, D.V.; Younes, M.W.F.; Abdel-Wahab, H.; Mansour, A.H. Synergistic efficacy of botanical blends with and without synthetic insecticides against Aedes aegypti and Culex annulirostris mosquitoes. J. Vector Ecol. 2005, 30, 284–288. [Google Scholar] [PubMed]
- Thangam, T.S.; Kathiresan, K. Synergistic effects of insecticides with plant extracts on mosquito larvae. Trop. Biomed. 1990, 7, 135–137. [Google Scholar]
- Pennetier, C.; Bouraima, A.; Chandre, F.; Piameu, M.; Etang, J.; Rossignol, M.; Sidick, I.; Zogo, B.; Lacroix, M.N.; Yadav, R.; et al. Efficacy of Olyset® Plus, a new long-lasting insecticidal net incorporating permethrin and piperonyl butoxide against multi-resistant malaria vectors. PLoS ONE 2013, 8, e75134. [Google Scholar] [CrossRef]
- Nikpour, F.; Vatandoost, H.; Hanafi-Bojd, A.A.; Raeisi, A.; Ranjbar, M.; Enayati, A.A.; Abai, M.R.; Shayeghi, M.R.; Pourreza, A. Evaluation of deltamethrin in combination of piperonyl butoxide (PBO) against pyrethroid resistant, malaria vector, in IRS implementation: An experimental semi-field trial in Iran. Iran. J. Arthropod Borne Dis. 2017, 11, 469–481. [Google Scholar]






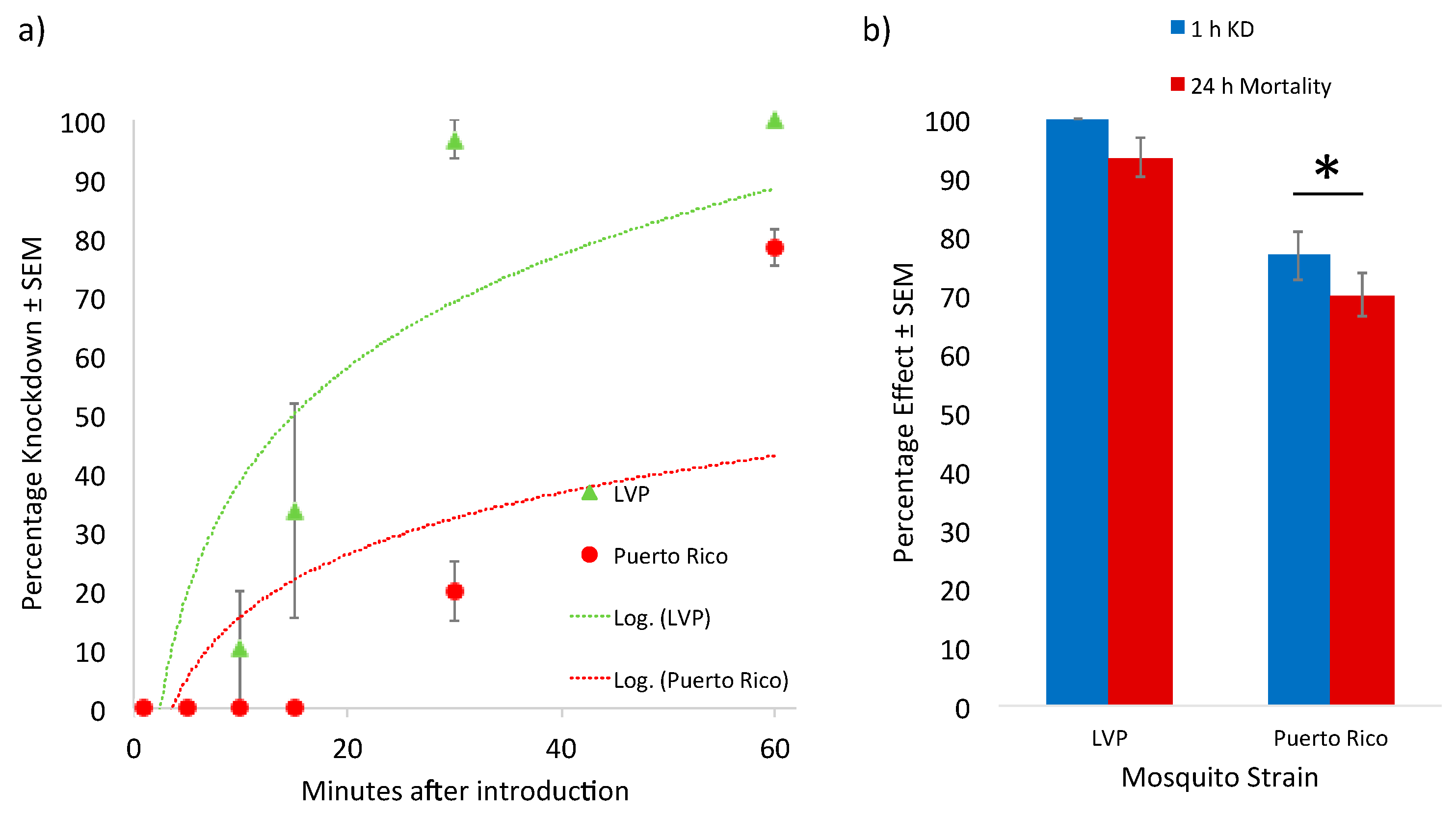{kind=link}
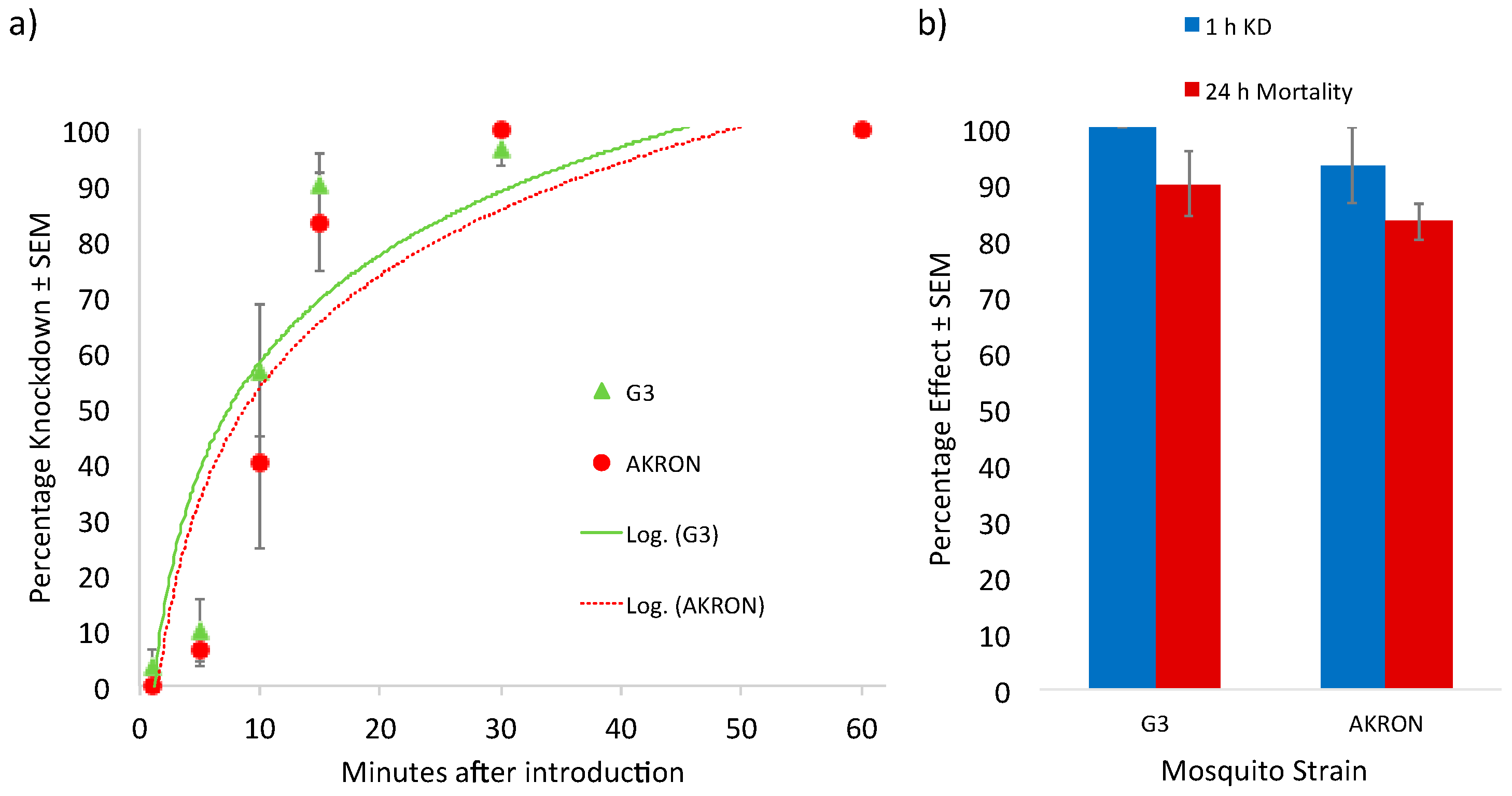{kind=link}
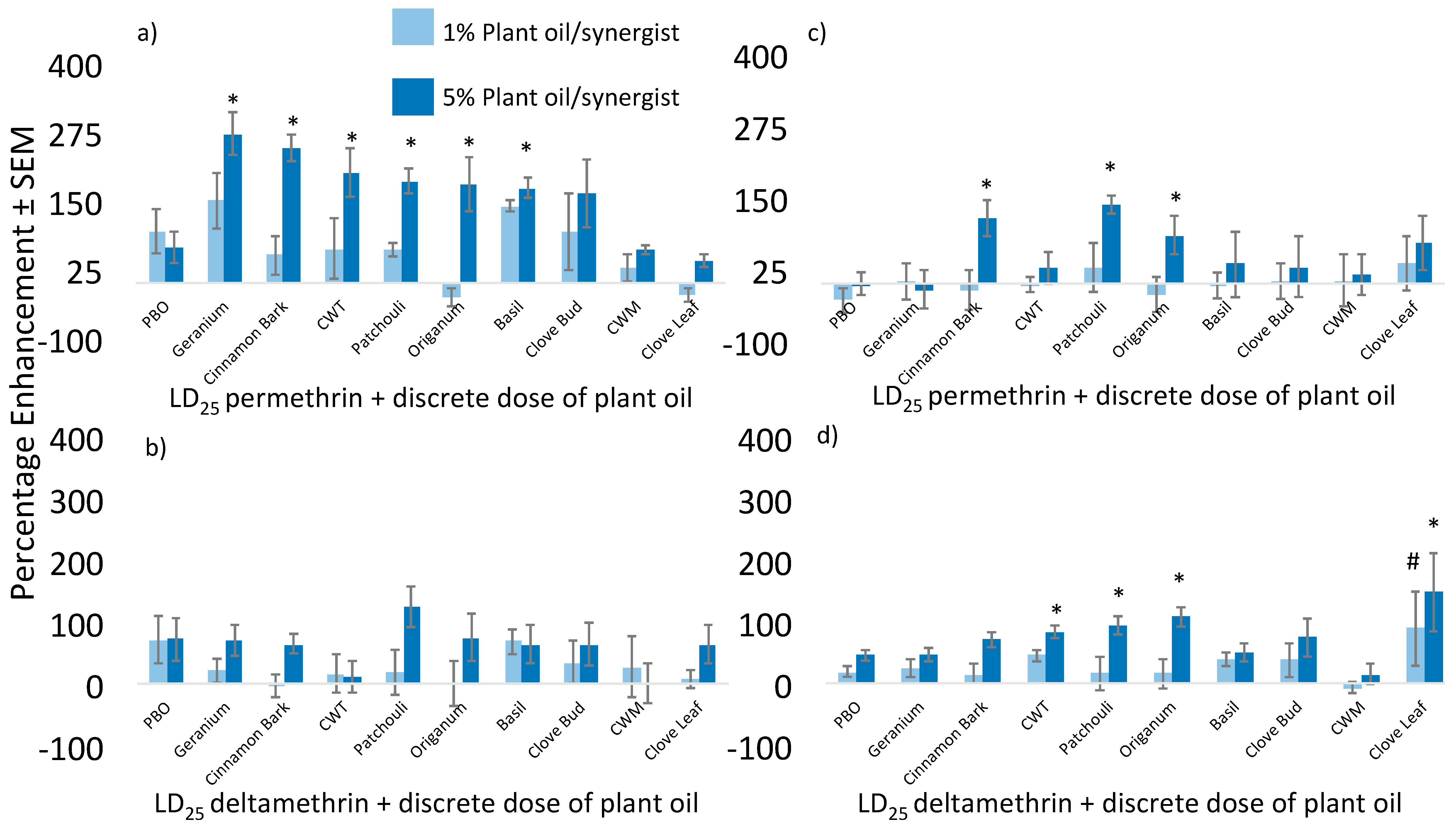{kind=link}
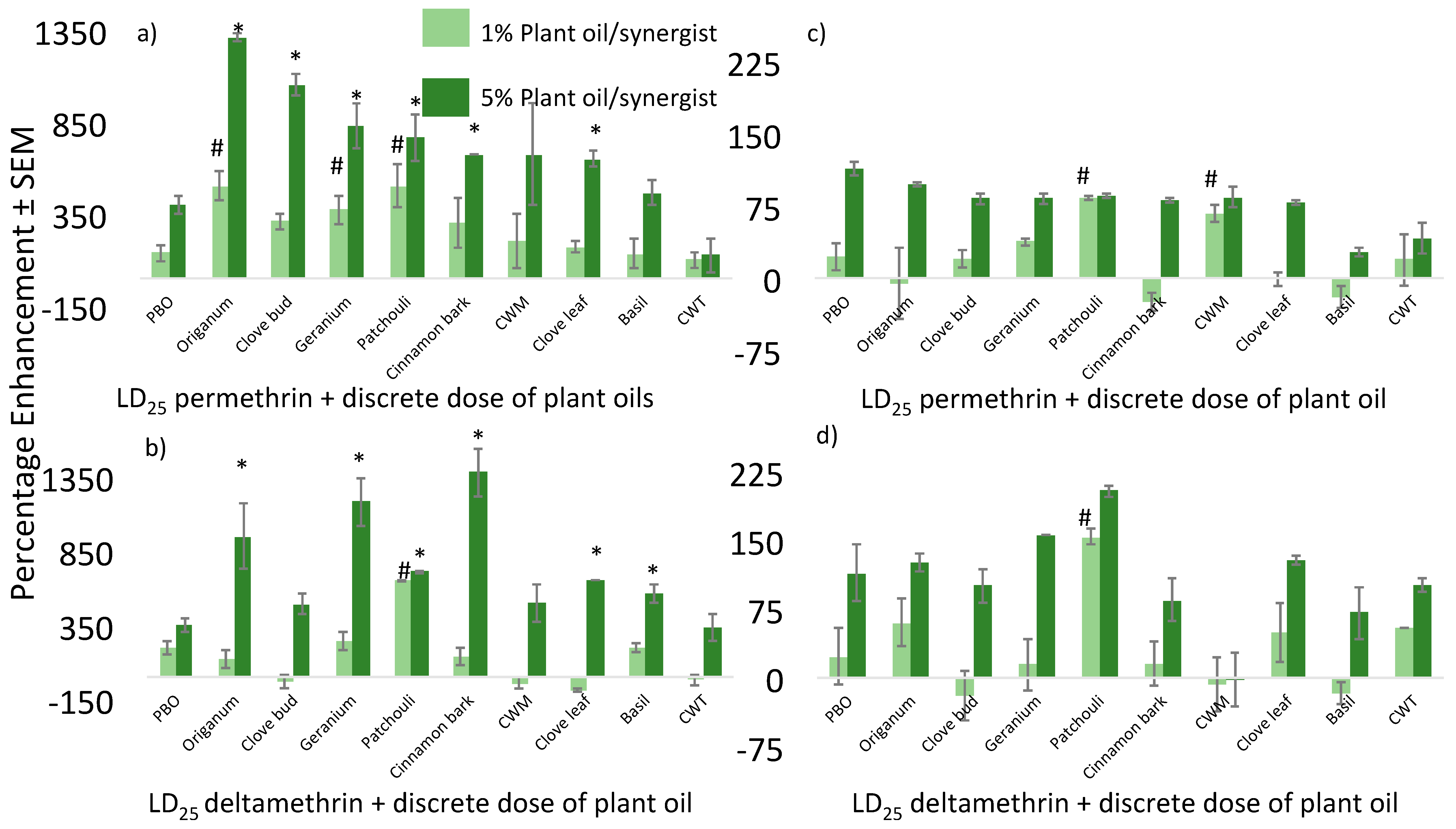{kind=link}
{kind=link}
| Compound | n | Species | Strain | LD50, μg/g (95% CI) | RR |
|---|---|---|---|---|---|
| Permethrin | 1350 | Anopheles gambiae | G3 | 0.632 (0.15–1.5) | |
| 1100 | Anopheles gambiae | AKRON | 3.8 (1.9–5.8) | 6.0 | |
| Deltamethrin | 875 | Anopheles gambiae | G3 | 0.0026 (0.001–0.004) | |
| 1425 | Anopheles gambiae | AKRON | 0.074 (0.06–0.09) | 28.4 | |
| Permethrin | 1300 | Aedes aegypti | Liverpool | 0.42 (0.31–0.51) | |
| 1100 | Aedes aegypti | Puerto Rico | 12.3 (9.6–15.6) | 29.3 | |
| Deltamethrin | 1525 | Aedes aegypti | Liverpool | 0.34 (0.16–1.5) | |
| 1100 | Aedes aegypti | Puerto Rico | 0.601 (0.4–1.03) | 1.8 | |
| Natural pyrethrins | 850 | Aedes aegypti | Liverpool | 1.64 (0.8–2.4) | |
| 700 | Aedes aegypti | Puerto Rico | 7.91 (6.1–11) | 4.8 |
| Strain | Synergist/Plant Oil | 1% Plant Oil/Synergist | 5% Plant Oil/Synergist | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Permethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | Permethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | |||
| Liverpool | PBO | 21.1 | 2 | 23.1 | 36 | 55.84 | 21.1 | 22.3 | 43.4 | 32.7 | −24.65 | |
| Patchouli | 26.7 | 6.7 | 33.4 | 42.7 | 27.84 | 26.7 | 60 | 86.7 | 76 | −12.34 | ||
| CWT a | 21 | 1 | 22 | 34 | 54.55 | 21 | 1 | 22 | 63 | 186.36 | ||
| Geranium | 12.8 | 0 | 12.8 | 29.6 | 131.25 | 12.8 | 17.3 | 30.1 | 43.2 | 43.52 | ||
| CWM b | 34 | 2 | 36 | 43 | 19.44 | 34 | 2 | 36 | 54 | 50.00 | ||
| Clove Bud | 24 | 4 | 28 | 36 | 28.57 | 24 | 35 | 59 | 48 | −18.64 | ||
| Cinnamon Bark | 18.4 | 4 | 22.4 | 27.2 | 21.43 | 18.4 | 36 | 54.4 | 61.6 | 13.24 | ||
| Basil | 33.3 | 60 | 93.3 | 80 | −14.26 | 33.3 | 92 | 125.3 | 90.6 | −27.69 | ||
| Origanum | 28.9 | 22 | 50.9 | 28.9 | −43.22 | 28.9 | 54 | 82.9 | 74.4 | −10.25 | ||
| Clove Leaf | 48.6 | 2 | 50.6 | 38.9 | −23.12 | 48.6 | 50 | 98.6 | 66.3 | −32.76 | ||
| Puerto Rico | PBO | 34 | 3 | 37 | 25 | −32.43 | 34 | 28 | 62 | 33 | −46.77 | |
| Patchouli | 35 | 8 | 43 | 60 | 39.53 | 35 | 48 | 83 | 83 | 0.00 | ||
| CWT a | 30 | 1 | 31 | 29 | −6.45 | 30 | 4 | 34 | 38 | 11.76 | ||
| Geranium | 53 | 1 | 54 | 55 | 1.85 | 53 | 15 | 68 | 48 | −29.41 | ||
| CWM b | 27 | 13 | 40 | 28 | −30.00 | 27 | 55 | 82 | 31 | −62.20 | ||
| Clove Bud | 45 | 4 | 49 | 47 | −4.08 | 45 | 79 | 124 | 57 | −54.03 | ||
| Cinnamon Bark | 31 | 4 | 35 | 28 | −20.00 | 31 | 71 | 102 | 64 | −37.25 | ||
| Basil | 37 | 41 | 78 | 36 | −53.85 | 37 | 11 | 48 | 49 | 2.08 | ||
| Origanum | 35 | 2 | 37 | 29 | −21.62 | 35 | 38 | 73 | 63 | −13.70 | ||
| Clove Leaf | 34 | 2 | 36 | 46 | 27.78 | 34 | 49 | 83 | 58 | −30.12 | ||
| Strain | Synergist/Plant Oil | 1% Plant Oil/Synergist | 5% Plant Oil/Synergist | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Deltamethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | Deltamethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | |||
| Liverpool | PBO | 26 | 3 | 29 | 43 | 48.28 | 26 | 14 | 40 | 43 | 7.50 | |
| CWM b | 29 | 1 | 30 | 37 | 23.33 | 30 | 14 | 44 | 30 | −31.82 | ||
| Clove Leaf | 22 | 2 | 24 | 29 | 20.83 | 23 | 42 | 65 | 43 | −33.85 | ||
| Geranium | 28 | 2 | 30 | 34 | 13.33 | 28 | 18 | 46 | 46 | 0.00 | ||
| Clove Bud | 29 | 2 | 31 | 33 | 6.45 | 29 | 30 | 59 | 39 | −33.90 | ||
| Origanum | 31 | 0 | 31 | 31 | 0.00 | 31 | 38 | 69 | 52 | −24.64 | ||
| CWT a | 27 | 5 | 32 | 31 | −3.13 | 27 | 5 | 32 | 30 | -6.25 | ||
| Patchouli | 31 | 7 | 38 | 36 | −5.26 | 31 | 43 | 74 | 65 | −12.16 | ||
| Cinnamon Bark | 27 | 3 | 30 | 26 | −13.33 | 27 | 36 | 63 | 43 | −31.75 | ||
| Basil | 35 | 48 | 83 | 44 | −46.99 | 35 | 63 | 98 | 43 | −56.12 | ||
| Puerto Rico | PBO | 43 | 2 | 45 | 55 | 22.22 | 43 | 24 | 67 | 67 | 0.00 | |
| CWM b | 54 | 4 | 58 | 50 | −13.79 | 54 | 54 | 108 | 62 | −42.59 | ||
| Clove Leaf | 20 | 2 | 22 | 37 | 68.18 | 20 | 47 | 67 | 53 | −20.90 | ||
| Geranium | 41 | 4 | 45 | 51 | 13.33 | 41 | 13 | 54 | 60 | 11.11 | ||
| Clove Bud | 35 | 4 | 39 | 47 | 20.51 | 35 | 74 | 109 | 65 | −40.37 | ||
| Origanum | 33 | 22 | 55 | 37 | −32.73 | 33 | 49 | 82 | 66 | −19.51 | ||
| CWT a | 38 | 1 | 39 | 55 | 41.03 | 38 | 3 | 41 | 70 | 70.73 | ||
| Patchouli | 33 | 8 | 41 | 41 | 0.00 | 33 | 46 | 79 | 63 | −20.25 | ||
| Cinnamon Bark | 38 | 8 | 46 | 49 | 6.52 | 38 | 64 | 102 | 75 | −26.47 | ||
| Basil | 45 | 13 | 58 | 62 | 6.90 | 45 | 39 | 84 | 67 | −20.24 | ||
| Strain | Synergist/Plant Oil | 1% Plant Oil/Synergist | 5% Plant Oil/Synergist | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Permethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | Permethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | |||
| G3 | PBO | 13.3 | 24.6 | 37.9 | 30.7 | −19.00 | 13.3 | 72 | 85.3 | 64 | −24.97 | |
| Geranium | 8.8 | 13.6 | 22.4 | 41.6 | 85.71 | 8.8 | 72 | 80.8 | 81.6 | 0.99 | ||
| CWM b | 6.6 | 2.7 | 9.3 | 12 | 29.03 | 6.7 | 48 | 54.7 | 42.6 | −22.12 | ||
| Clove Bud | 8.8 | 16 | 24.8 | 31.2 | 25.81 | 8.8 | 77.6 | 86 | 85.6 | −0.93 | ||
| CWT a | 9.3 | 6.7 | 16 | 20 | 25.00 | 9.3 | 64.7 | 74 | 75.3 | 1.76 | ||
| Patchouli | 17.3 | 78.6 | 95.9 | 90.7 | −5.42 | 17.3 | 97.3 | 114.6 | 98.7 | −13.87 | ||
| Clove Leaf | 16 | 54.7 | 70.7 | 60 | −15.13 | 16 | 86.7 | 102.7 | 84 | −18.21 | ||
| Origanum | 12 | 44.7 | 56.7 | 44 | −22.40 | 12 | 95.3 | 107.3 | 98 | −8.67 | ||
| Cinnamon Bark | 13.6 | 29.6 | 43.2 | 30.4 | −29.63 | 13.6 | 81.6 | 95.2 | 100 | 5.04 | ||
| Basil | 12.8 | 36.8 | 49.6 | 32 | −35.48 | 12.8 | 95 | 107.8 | 85.6 | −20.59 | ||
| Akron | PBO | 44.4 | 24 | 68.4 | 53.7 | −21.49 | 44.4 | 47 | 91.4 | 92 | 0.66 | |
| Geranium | 56 | 13.3 | 69.3 | 76 | 9.67 | 56 | 30.7 | 86.7 | 100 | 15.34 | ||
| CWM b | 59 | 28 | 87 | 92 | 5.75 | 59 | 48 | 107 | 100 | −6.54 | ||
| Clove Bud | 56 | 2.7 | 58.7 | 66.7 | 13.63 | 56 | 77.3 | 133.3 | 100 | −24.98 | ||
| CWT a | 59 | 22.7 | 81.7 | 68 | −16.77 | 59 | 85.3 | 144.3 | 79 | −45.25 | ||
| Patchouli | 59 | 68 | 127 | 99 | −22.05 | 59 | 96 | 155 | 100 | −35.48 | ||
| Clove Leaf | 56 | 5.3 | 61.3 | 54.7 | −10.77 | 56 | 82.7 | 138.7 | 97.3 | −29.85 | ||
| Origanum | 52 | 4 | 56 | 49.3 | −11.96 | 52 | 76 | 128 | 98.7 | −22.89 | ||
| Cinnamon Bark | 56 | 37.3 | 93.3 | 42.7 | −54.23 | 56 | 96 | 152 | 98.7 | −35.07 | ||
| Basil | 56 | 21.3 | 77.3 | 45.3 | −41.40 | 56 | 50.7 | 106.7 | 70.7 | −33.74 | ||
| Strain | Synergist/Plant Oil | 1% Plant Oil/Synergist | 5% Plant Oil/Synergist | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Deltamethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | Deltamethrin | Synergist/Oil Alone | Expected Mortality | Observed Mortality | Cotoxicity Factor | |||
| G3 | PBO | 12.5 | 18 | 30.5 | 31.5 | 3.28 | 12.5 | 49 | 61.5 | 45.5 | −26.02 | |
| Geranium | 6.7 | 10.6 | 17.3 | 22.7 | 31.21 | 6.7 | 62.7 | 69.4 | 85.3 | 22.91 | ||
| Patchouli | 18.4 | 92 | 110.4 | 91.2 | −17.39 | 18.4 | 97 | 115.4 | 97.6 | −15.42 | ||
| Basil | 13.3 | 8 | 21.3 | 20 | −6.10 | 13.3 | 45.3 | 58.6 | 44 | −24.91 | ||
| Cinnamon Bark | 6.7 | 26.7 | 33.4 | 16 | −52.10 | 6.7 | 77.3 | 84 | 98.7 | 17.50 | ||
| Origanum | 10.7 | 21.3 | 32 | 23.2 | −27.50 | 10.7 | 80 | 90.7 | 96 | 5.84 | ||
| Clove Leaf | 14.7 | 6.7 | 21.4 | 53 | 147.66 | 6.7 | 85.3 | 92 | 84 | −8.70 | ||
| Clove Bud | 17.6 | 17.6 | 35.2 | 13.6 | −61.36 | 17.6 | 81.6 | 99.2 | 80 | −19.35 | ||
| CWT a | 16 | 13.6 | 29.6 | 14 | −52.70 | 16 | 72.8 | 88.8 | 49.3 | −44.48 | ||
| CWM b | 9.3 | 8 | 17.3 | 6 | −65.32 | 9.3 | 63.3 | 72.6 | 49.3 | −32.09 | ||
| Akron | PBO | 45.5 | 27 | 72.5 | 62 | −14.48 | 45.5 | 49 | 94.5 | 94 | −0.53 | |
| Geranium | 43 | 11 | 54 | 48 | −11.11 | 43 | 39 | 82 | 100 | 21.95 | ||
| Patchouli | 36 | 74 | 110 | 79 | −28.18 | 36 | 99 | 135 | 93 | −31.11 | ||
| Basil | 42.4 | 29 | 71.4 | 37.6 | −47.34 | 42.4 | 49 | 91.4 | 63.2 | −30.85 | ||
| Cinnamon Bark | 49.1 | 33 | 82.1 | 55.4 | −-32.52 | 49.1 | 96 | 145.1 | 84.5 | −41.76 | ||
| Origanum | 43 | 2.6 | 45.6 | 65 | 42.54 | 43 | 84 | 127 | 89 | −29.92 | ||
| Clove Leaf | 45.7 | 4 | 49.7 | 62.8 | 26.36 | 45.7 | 81.3 | 127 | 90.9 | −28.43 | ||
| Clove Bud | 47.2 | 17.3 | 64.5 | 39.2 | −39.22 | 47.2 | 76 | 123.2 | 88 | −28.57 | ||
| CWT a | 52 | 22.7 | 74.7 | 69.3 | −7.23 | 52 | 89.3 | 141.3 | 84 | −40.55 | ||
| CWM b | 47.3 | 27 | 74.3 | 44.7 | −39.84 | 47.3 | 54 | 101.3 | 46.7 | −53.90 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norris, E.J.; Johnson, J.B.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes. Insects 2018, 9, 132. https://doi.org/10.3390/insects9040132
Norris EJ, Johnson JB, Gross AD, Bartholomay LC, Coats JR. Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes. Insects. 2018; 9(4):132. https://doi.org/10.3390/insects9040132
Chicago/Turabian StyleNorris, Edmund J., Jacob B. Johnson, Aaron D. Gross, Lyric C. Bartholomay, and Joel R. Coats. 2018. "Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes" Insects 9, no. 4: 132. https://doi.org/10.3390/insects9040132
APA StyleNorris, E. J., Johnson, J. B., Gross, A. D., Bartholomay, L. C., & Coats, J. R. (2018). Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes. Insects, 9(4), 132. https://doi.org/10.3390/insects9040132