Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii





Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Colony
2.2. Bacteria Isolation and Culture Conditions
2.3. Larval Oral Bioassay
2.4. Larval and Pupal Dipping Bioassay
2.5. Pupal Direct-Spray Bioassay
2.6. Adult Insect Bioassay
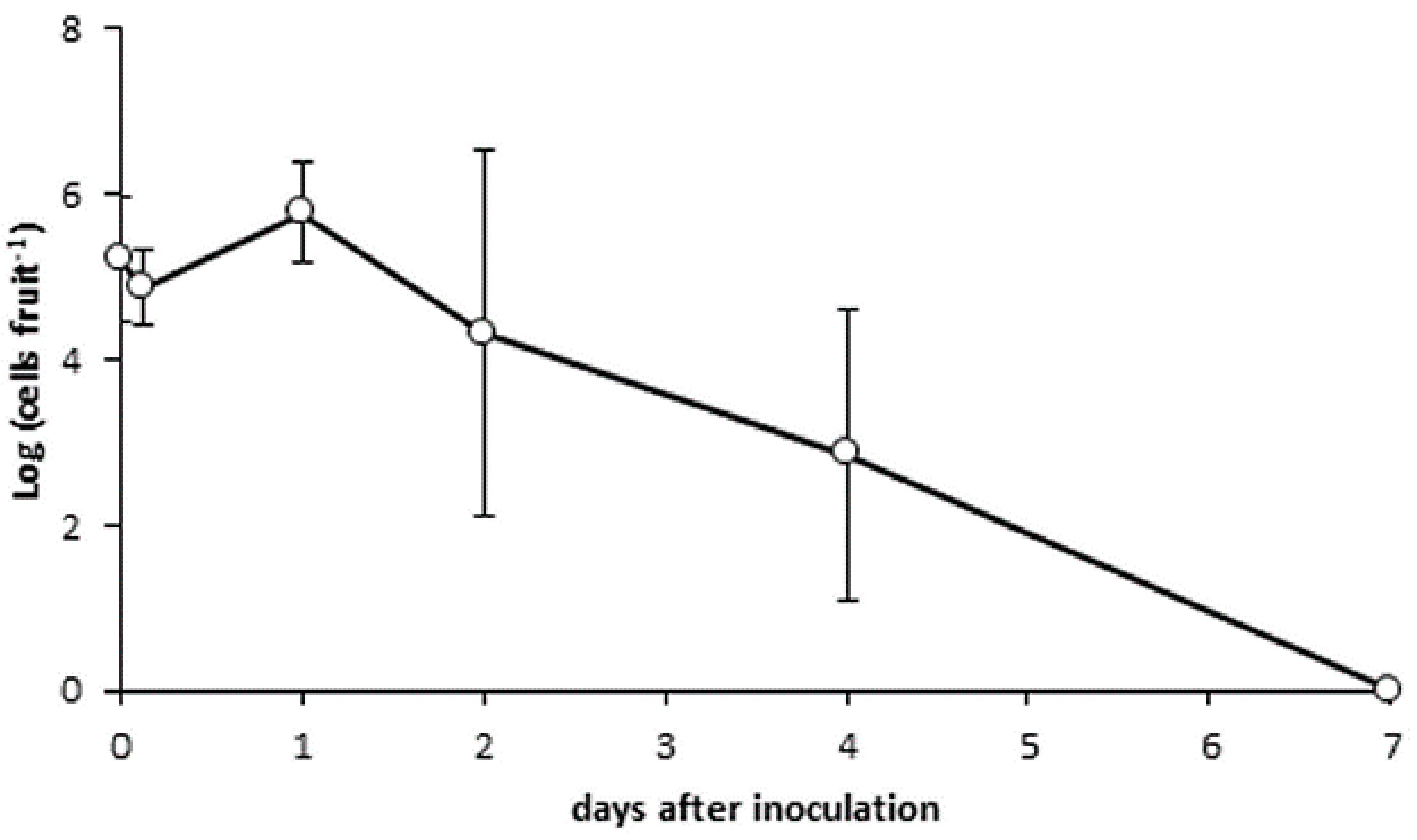
2.7. Bacterial Persistence on Cherry Fruit
2.8. Data Analysis
3. Results
3.1. Larval Oral Bioassay
3.2. Larval and Pupal Dipping Bioassay
3.3. Pupal Direct-Spray Bioassay
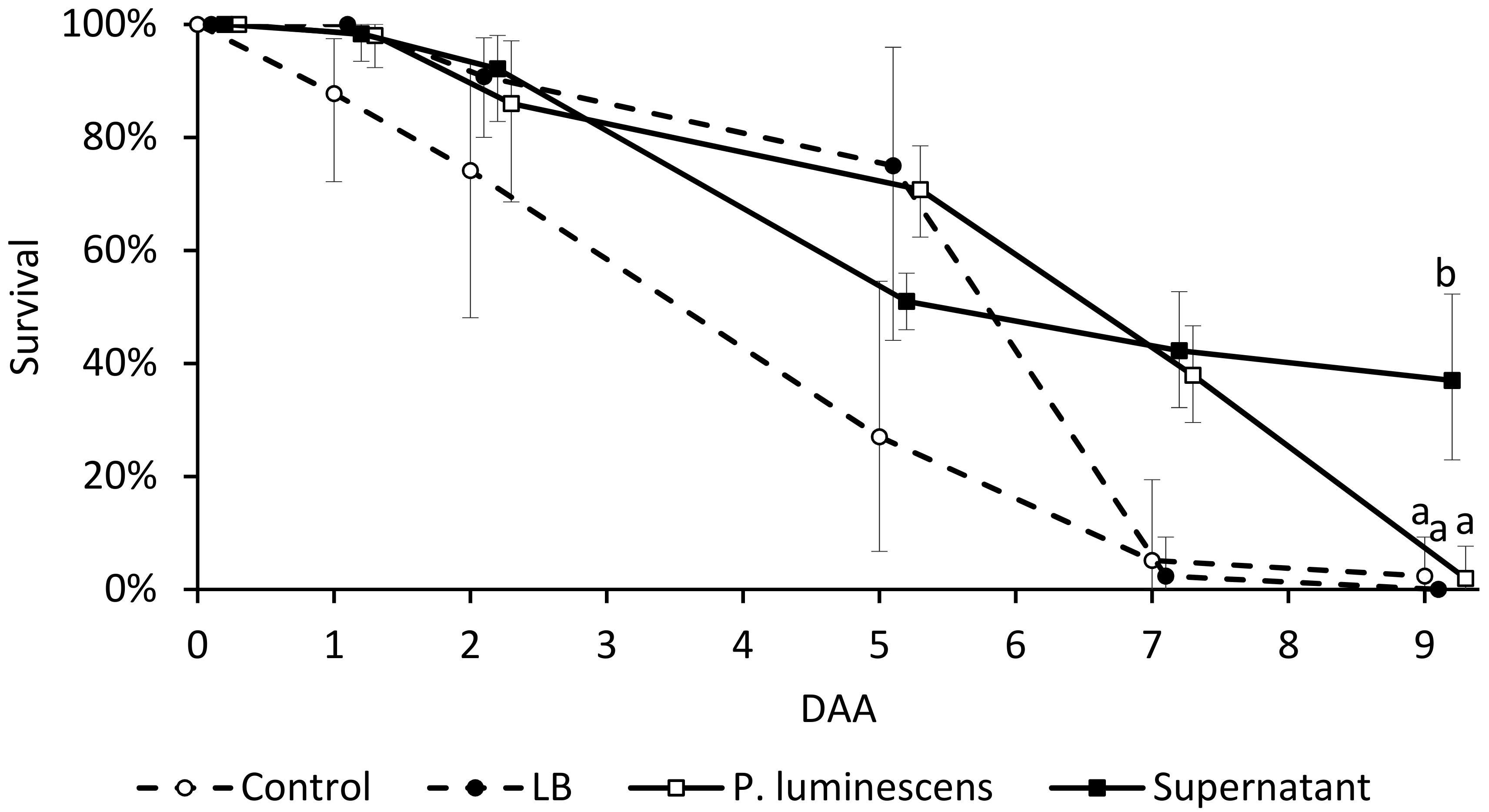
3.4. Adult Insect Bioassay
3.5. Bacterial Persistence on Cherry Skin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cini, A.; Ioratti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.D.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted wing drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Sato, R. Bionomics of the cherry drosophila, Drosophila suzukii Matsumura (Diptera: Drosophilidae) in Fukushima prefecture (Japan). Ann. Rep. Soc. Plant Prot. North Jpn. 1995, 46, 164–172. [Google Scholar]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Haviland, D.R.; Beers, E.H. Chemical control programs for Drosophila suzukii that comply with international limitations on pesticide residues for exported sweet cherries. J. Integr. Pest Manag. 2012, 3, F1–F6. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Rahoo, A.M.; Mukhtar, T.; Gowen, S.R.; Pembroke, B. Virulence of entomopathogenic bacteria Xenorhabdus bovienii and Photorhabdus luminescens against Galleria mellonella larvae. Pak. J. Zool. 2011, 43, 543–548. [Google Scholar]
- Miller, B.; Anfora, G.; Buffington, M.; Daane, K.M.; Dalton, D.T.; Hoelmer, K.M.; Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Loni, A.; et al. Seasonal occurrence of resident parasitoids associated with Drosophila suzukii in two small fruit production regions of Italy and the USA. Bull. Insectol. 2015, 68, 255–263. [Google Scholar]
- Wang, X.-G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Renkema, J.M.; Telfer, Z.; Gariepy, T.; Hallett, R.H. Dalotia coriaria as a predator of Drosophila suzukii: Functional responses, reduced fruit infestation and molecular diagnostics. Biol. Control 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Woltz, J.; Donahue, K.; Bruck, D.; Lee, J. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 2015, 139, 759–770. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Collins, D.A.; Blackburn, L.F.; Audsley, N.; Bell, H.A. Preliminary screening of potential control products against Drosophila suzukii. Insects 2014, 5, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Audsley, N. Further screening of entomopathogenic fungi and nematodes as control agents for Drosophila suzukii. Insects 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Lázaro, J.M.; Mellín-Rosas, M.A.; González-Padilla, V.D.; Sánchez-González, J.A.; Moreno-Carrillo, G.; Arredondo-Bernal, H.C. Susceptibility of Drosophila suzukii Matsumura (Diptera: Drosophilidae) to entomophatogenic fungi. Southwest. Entomol. 2014, 39, 201–203. [Google Scholar] [CrossRef]
- Wise, J.C.; Van Woerkom, A.H.; Wheeler, C.E.; Isaacs, R. IR-4 Attract N Kill tactics for control of spotted wing drosophila in blueberry. Arthropod Manag. Tests 2018, 43. [Google Scholar] [CrossRef]
- EU-Pesticides-Database. European Union Pesticides Database. 2018. Available online: http://ec.europa.eu/food/plant/pesticides/eu-311 pesticides-database (accessed on 14 August 2018).
- Mahar, A.; Jan, N.; Mahar, G.M.; Mahar, A.Q. Control of insects with entomopathogenic bacterium Xenorhabdus nematophila and its toxic secretions. Int. J. Agric. Biol. 2008, 10, 52–56. [Google Scholar]
- Ruiu, L. Insect pathogenic bacteria in integrated pest management. Insects 2015, 6, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Fanning, P.D.; Grieshop, M.J.; Isaacs, R. Efficacy of biopesticides on spotted wing drosophila, Drosophila suzukii Matsumura in fall red raspberries. J. Appl. Entomol. 2018, 142, 26–32. [Google Scholar] [CrossRef]
- Cahenzli, F.; Strack, T.; Daniel, C. Screening of 25 different natural crop protection products against Drosophila suzukii. J. Appl. Entomol. 2018, 142, 563–577. [Google Scholar] [CrossRef]
- Blackburn, M.; Golubeva, E.; Bowen, D.; Ffrench-Constant, R.H. A novel insecticidal toxin from Photorhabdus luminescens, toxin complex a (Tca), and its histopathological effects on the midgut of Manduca sexta. Appl. Environ. Microbiol. 1998, 64, 3036–3041. [Google Scholar] [PubMed]
- Waterfield, N.; Dowling, A.; Sharma, S.; Daborn, P.J.; Potter, U.; Ffrench-Constant, R.H. Oral toxicity of Photorhabdus luminescens W14 toxin complexes in Escherichia coli. Appl. Environ. Microbiol. 2001, 67, 5017–5024. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Fatig, R.O.; Orr, G.L.; Schafer, G.W.; Strickland, J.A.; Sukhapinda, K.; Woodsworth, A.T.; Petell, J.K. Photorhabdus luminescens W-14 insecticidal activity consists of at least two similar but distinct proteins. Purification and characterization of Toxin A and Toxin B. J. Biol. Chem. 1999, 274, 9836–9842. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Bhatnagar, R.K. Insecticidal toxic proteins produced by Photorhabdus luminescens akhurstii, a symbiont of Heterorhabditis indica. J. Nematol. 2002, 34, 23–27. [Google Scholar] [PubMed]
- Mahar, A.N.; Jan, N.D.; Mahar, A.Q.; Mahar, G.M.; Hullio, M.H.; Lanjar, A.G. Efficacy of entomopathogenic bacterium Photorhabdus luminescens and its metabolites against diamondback moth Plutella xylostella larvae on chinese cabbage and artificial diet. Pak. J. Nematol. 2008, 26, 69–82. [Google Scholar]
- Blackburn, M.B.; Domek, J.M.; Gelman, D.B.; Hu, J.S. The broadly insecticidal Photorhabdus luminescens toxin complex a (Tca): Activity against the Colorado potato beetle, Leptinotarsa decemlineata, and sweet potato whitefly, Bemisia tabaci. J. Insect Sci. 2005, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2007, 49, 436–451. [Google Scholar]
- Shawer, R.; Tonina, L.; Tirello, P.; Duso, C.; Mori, N. Laboratory and field trials to identify effective chemical control strategies for integrated management of Drosophila suzukii in European cherry orchards. Crop Prot. 2018, 103, 73–80. [Google Scholar] [CrossRef]
- Tonina, L.; Mori, N.; Giomi, F.; Battisti, A. Development of Drosophila suzukii. J. Pest Sci. 2016, 89, 667–678. [Google Scholar] [CrossRef]
- Kehres, J.; Denon, D.; Mauléon, H. A simple technique to estimate, in situ, population densities of an entomopathogenic nematode (Heterorhabditis indica) in sandy soils. Nematology 2001, 3, 285–287. [Google Scholar] [CrossRef]
- Sicard, M.; Hering, S.; Schulte, R.; Gaudriault, S.; Schulenburg, H. The effect of Photorhabdus luminescens (Enterobacteriaceae) on the survival, development, reproduction and behaviour of Caenorhabditis elegans (Nematoda: Rhabditidae). Environ. Microbiol. 2007, 9, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Berry, R.E.; Blouin, M.S. Identification of symbiotic bacteria (Photorhabdus and Xenorhabdus) from the entomopathogenic nematodes Heterorhabditis marelatus and Steinernema oregonense based on 16S rDNA sequence. J. Invertebr. Pathol. 2001, 77, 87–91. [Google Scholar] [CrossRef] [PubMed]
- da Silva, O.S.; Prado, G.R.; da Silva, J.L.R.; Silva, C.E.; da Costa, M.; Heermann, R. Oral toxicity of Photorhabdus luminescens and Xenorhabdus nematophila (Enterobacteriaceae) against Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2013, 112, 2891–2896. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Instructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides. 1981. Available online: http://apps.who.int/iris/bitstream/handle/10665/69615/WHO?sequence=1 (accessed on 14 August 2018).
- Kanzawa, T. Studies on Drosophila suzukii Mats. Kofu. Rev. Appl. Entomol. 1939, 29, 622. [Google Scholar]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Yee, W.L.; Alston, D.G. Effects of spinosad, spinosad bait, and chloronicotinyl insecticides on mortality and control of adult and larval western cherry fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 2006, 99, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Waterfield, N.R.; Ciche, T.; Clarke, D. Photorhabdus and a host of hosts. Annu. Rev. Microbiol. 2009, 63, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Sharad-Mohan, A.; Gaur, H.S. Successful management of mango mealy bug, Drosicha mangiferae by Photorhabdus luminescens, a symbiotic bacterium from entomopathogenic nematode Heterorhabditis indica. Int. J. Nematol. 2003, 14, 195–198. [Google Scholar]
- Uma, G.P.; Prabhuraj, A. Bio-efficacy of Photorhabdus luminescens, a symbiotic bacterium against Thrips palmi Karny (Thripidae: Thysanoptera). J. Biopestic. 2010, 3, 458–462. [Google Scholar]
- Musser, F.R.; Nyrop, J.P.; Shelton, A.M. Integrating biological and chemical controls in decision making: European corn borer (Lepidoptera: Crambidae) control in sweet corn as an example. J. Econ. Entomol. 2006, 99, 1538–1549. [Google Scholar] [CrossRef] [PubMed]
- Satinder, K.B.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochem. 2006, 41, 323–342. [Google Scholar]
- Ramesh, A.K.; Prabhuraj, A.; Ashoka, J.; Hanchinal, S.G.; Sharanagouda, H. Generation and evaluation of nanoparticles of supernatant of Photorhabdus luminescens (Thomas and Poinar) against mite and aphid pests of cotton for enhanced efficacy. Curr. Sci. 2017, 112, 2312–2316. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Concentration (Cells mL−1) | Larvae | Pupae | Adults | Immatures | Total Individuals | |
|---|---|---|---|---|---|---|
| 2 DAA | 4 DAA | 10 DAA | 10 DAA | 10 DAA | 10 DAA | |
| 3.5 × 108 | 10.0 (10.0–10.0) a | 36.7 (8.0–65.0) a | 10.0 (−33.0–53.0) a | 50.0 (25.2–74.8) a | 46.7 (8.7–84.6) a | 96.7 (82.3–111) a |
| 3.5 × 107 | 3.3 (−11–17.6) a | 33.3 (−4.6–71.3) ab | 3.3 (−11.0–177.0) a | 53.3 (−22.6–129.0) a | 36.7 (−15–88.4) a | 90.0 (65.1–114.8) ab |
| 3.5 × 106 | 6.7 (−22–35.4) a | 10.0 (−14.8–34.8) c | 10.0 (−14.8–34.8) a | 53.3 (−22.6–129.0) a | 20.0 (−14.8–34.8) ab | 73.3 (44.6–102.0) bc |
| 3.5 × 105 | 6.7 (−22–35.4) a | 13.3 (−1.0–27.7) bc | 0.0 (0.0–0.0) a | 53.3 (1.6–105.0) a | 13.3 (−1.0–27.7) b | 66.7 (9.3–124.0) c |
| 3.5 × 104 | 0.0 (0.0–0.0) a | 6.7 (−22.0–35.4) c | 6.7 (−22.0–35.4) a | 46.7 (18.0–75.4) a | 13.3 (−15.4–42.0) b | 60.0 (60.0–60.0) c |
| 3.5 × 103 | 0.0 (0.0–0.0) a | 10.0 (−33.0–53.0) c | 3.3 (−11.0–17.7) a | 46.7 (8.7–84.6) a | 13.3 (−24.6–51.3) b | 60.0 (10.3–109.7) c |
| Control | 3.3 (−11.0–17.7) a | 6.7 (−7.6–21.0) c | 0.0 (0.0–0.0) a | 6.7 (−7.7–21.0) b | 6.7 (−7.7–21.0) b | 13.3 (−1.0–27.7) d |
| Treatment on Larvae | Larvae, 4 DAA | Immatures, 10 DAA | Total Individuals, 10 DAA |
| Bacterial supernatant | 24 (11.6–32.4) a | 44 (28.4–55.6) a | 74 (51.6–92.4) a |
| Supernatant 10-fold dilution | 12 (−1.6–25.6) b | 18 (1.8–34.2) b | 34 (15.2–52.8) b |
| Control | 4 (−2.8–10.8) b | 12 (1.6–22.4) b | 28 (17.6–38.4) b |
| Treatment on Pupae | Pupae, 9 DAA | Adults, 9 DAA | Total Individuals, 9 DAA |
| Bacterial supernatant | 38 (17.6–58.4) a | 36 (15.2–56.8) a | 74 (59.8–88.2) a |
| Supernatant 10-fold dilution | 26 (9.3–42.7) a | 16 (9.2–22.8) b | 42 (25.8–58.2) b |
| Control | 26 (19.2–32.8) a | 6 (−0.8–12.8) b | 32 (26.4–37.6) b |
| Concentration (Cells mL−1) | Larvae | Pupae | Adults | Immatures | Total Individuals | |
|---|---|---|---|---|---|---|
| 2 DAA | 4 DAA | 9 DAA | 10 DAA | 10 DAA | 10 DAA | |
| 3.5 × 108 | 6.7 (−7.7–21.0) a | 10.0 (10.0–10.0) a | 60.0 (−5.0–125.7) a | 16.7 (−35.0–68.4) a | 70.0 (4.3–135.7) a | 86.7 (58.0–115.4) a |
| 3.5 × 107 | 3.3 (−11.0–17.7) a | 10.0 (−14.0–34.8) a | 36.7 (22.3–51) ab | 16.7 (2.3–31.0) a | 46.7 (8.7–84.6) ab | 63.3 (11.6–115) ab |
| 3.5 × 106 | 3.3 (−11.0–17.7) a | 3.3 (−11.0–17.7) a | 16.7 (−55.0–88.4) b | 30.0 (−13.0–73.0) a | 20.0 (−45.7–85.7) b | 50.0 (−15.7–115.7) bc |
| Control | 3.3 (−11.0–17.7) a | 3.3 (−11.0–17.7) a | 13.3 (−1.0–27.7) b | 10.0 (10.0–10.0) a | 16.7 (2.3–31) b | 26.7 (12.3–41.0) c |
| Concentration (Cells mL−1) | Pupae | Adults | Total Individuals |
|---|---|---|---|
| 3.5 × 108 | 46.7 (32.3–61.0) a | 26.7 (−45.0–98.4) a | 73.3 (16.0–130.7) a |
| 3.5 × 107 | 46.7 (8.7–84.6) a | 23.3 (−39.2–85.8) a | 70.0 (4.3–135.7) a |
| 3.5 × 106 | 43.3 (5.4–81.3) ab | 23.3 (9.0–37.7) a | 66.7 (15.0–118.4) ab |
| Control | 23.3 (9.0–37.7) b | 6.7 (−7.7–21.0) a | 30.0 (5.2–54.8) b |
| Concentration (Cells mL−1) | Pupae | Adults | Total Individuals |
|---|---|---|---|
| 3.5 × 108 | 63.3 (6.0–120.7) a | 36.7 (−20.7–94.0) a | 100.0 (100.0–100.0) a |
| 3.5 × 107 | 50.0 (−15.7–115.7) a | 50.0 (−15.7–115.7) a | 100.0 (100.0–100.0) a |
| 3.5 × 106 | 63.3 (34.6–92.0) a | 30.0 (5.2–54.8) ab | 93.3 (79.0–107.7) ab |
| 3.5 × 105 | 63.3 (49.0–77.7) a | 26.7 (12.3–41.0) ab | 90.0 (65.2–114.8) ab |
| 3.5 × 104 | 56.7 (42.3–71.0) a | 30.0 (5.2–54.8) ab | 86.7 (58.0–115.4) ab |
| 3.5 × 103 | 50.0 (7.0–93.0) a | 33.3 (19.0–47.7) ab | 83.3 (45.4–121.3) b |
| Control | 10.0 (10.0–10.0) b | 10.0 (10.0–10.0) b | 20.0 (20.0–20.0) c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shawer, R.; Donati, I.; Cellini, A.; Spinelli, F.; Mori, N. Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects 2018, 9, 148. https://doi.org/10.3390/insects9040148
Shawer R, Donati I, Cellini A, Spinelli F, Mori N. Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects. 2018; 9(4):148. https://doi.org/10.3390/insects9040148
Chicago/Turabian StyleShawer, Rady, Irene Donati, Antonio Cellini, Francesco Spinelli, and Nicola Mori. 2018. "Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii" Insects 9, no. 4: 148. https://doi.org/10.3390/insects9040148
APA StyleShawer, R., Donati, I., Cellini, A., Spinelli, F., & Mori, N. (2018). Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects, 9(4), 148. https://doi.org/10.3390/insects9040148