Molecular Analysis of Forensically Important Blow Flies in Thailand


Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.2. DNA Extraction, Amplification, and Sequencing
2.3. DNA Sequence Alignment and Sequence Analysis
2.4. Nucleotide Sequence Accession Numbers
2.5. DNA Marker Assessments (COI and COII) for Species Identification
2.5.1. Distance-Based Analysis
2.5.2. Tree-Based Analysis
3. Results
3.1. Sequence Analysis
3.2. Genetic Variation
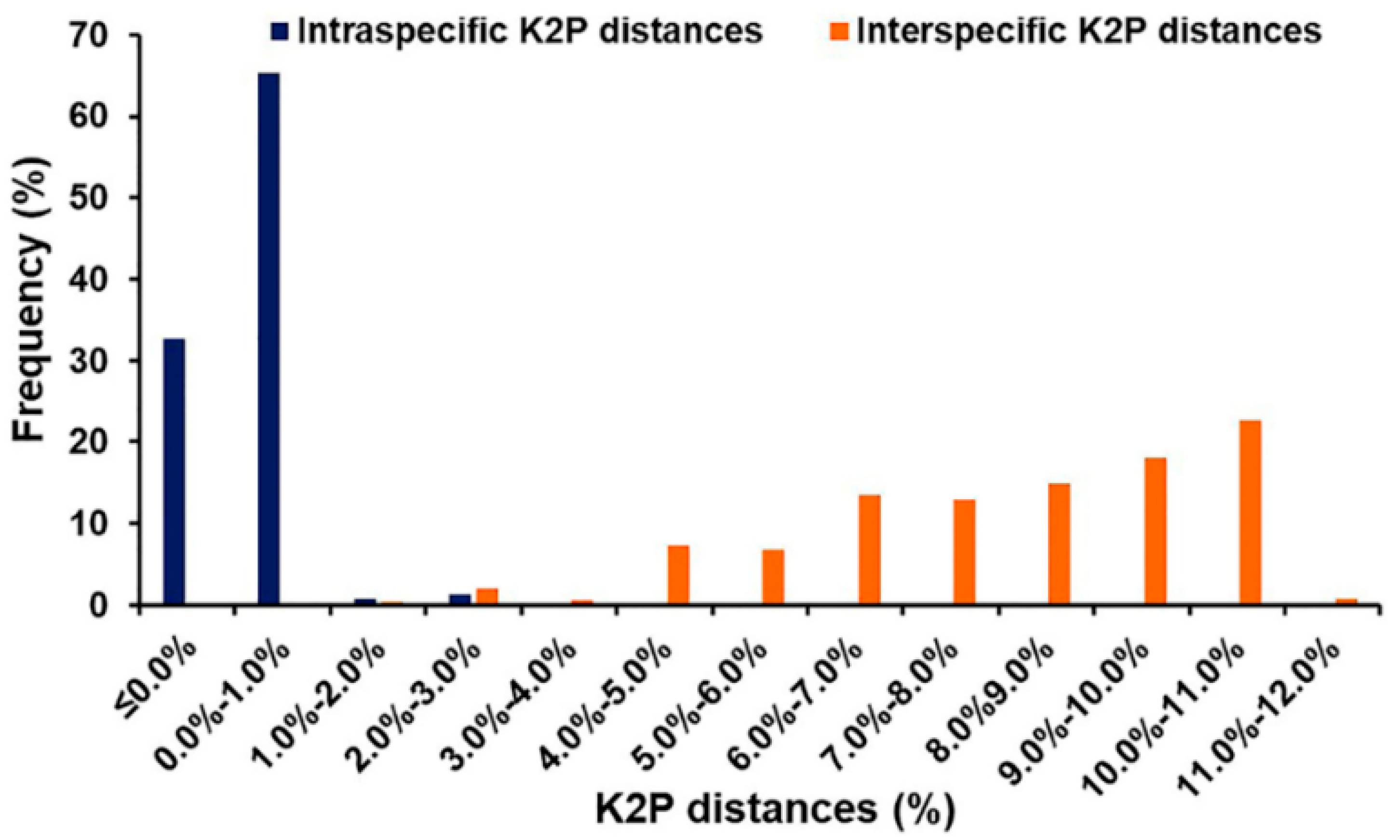
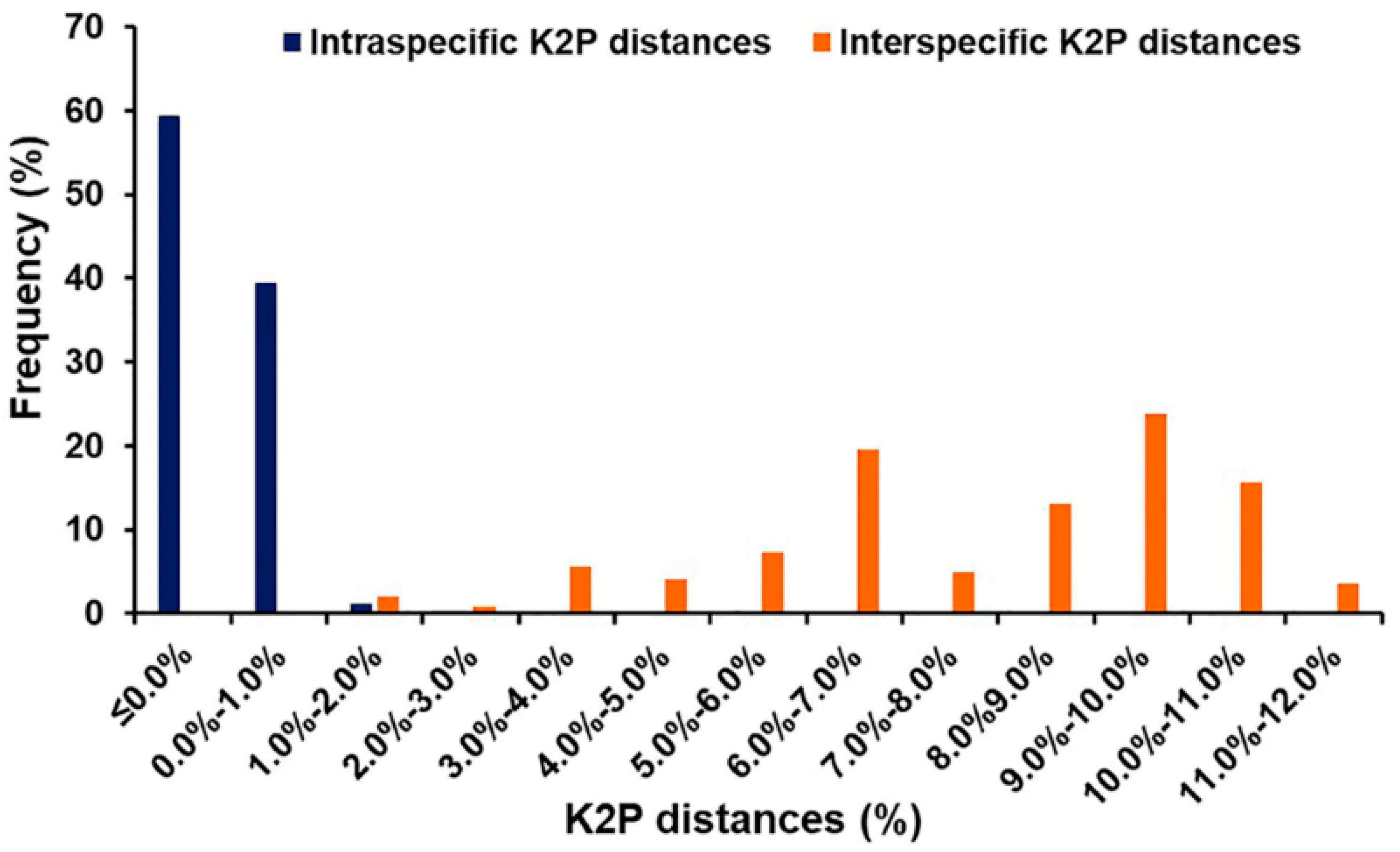
3.3. Distance-Based Identification
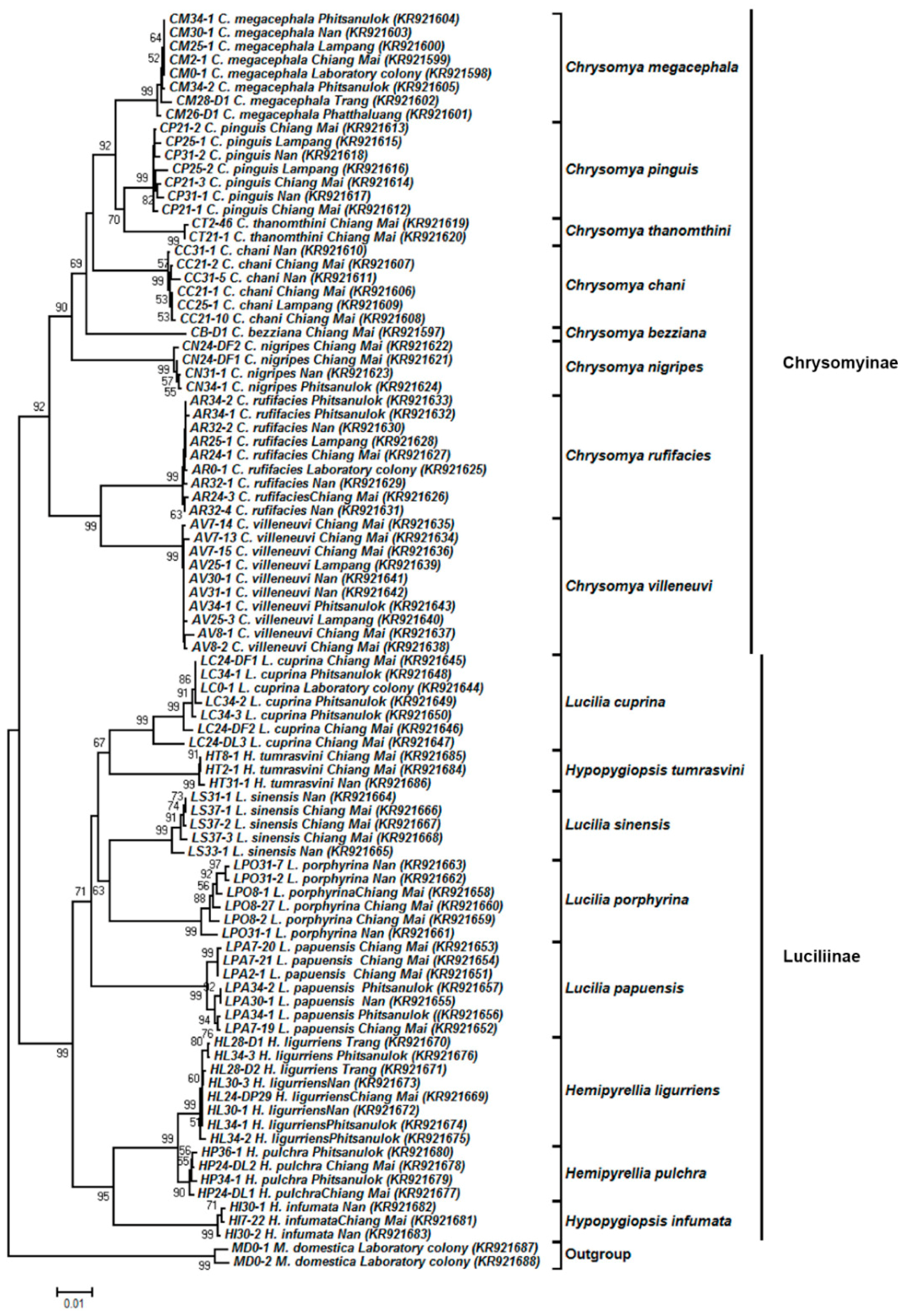
3.4. Tree-Based Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lane, R.P. An investigation into blowfly (Diptera: Calliphoridae) succession on corpses. J. Nat. Hist. 1975, 9, 581–588. [Google Scholar] [CrossRef]
- Catts, E.P.; Goff, M.L. Forensic entomology in criminal investigations. Annu. Rev. Entomol. 1992, 37, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Reibe, S.; Madea, B. How promptly do blowflies colonise fresh carcasses? A study comparing indoor with outdoor locations. Forensic Sci. Int. 2010, 195, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Amendt, J.; Richards, C.S.; Campobasso, C.P.; Zehner, R.; Hall, M.J.R. Forensic entomology: Applications and limitations. Forensic Sci. Med. Pathol. 2011, 7, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.A.; Downton, M.; Wallman, J.F. Thermal attributes of Chrysomya species. Entomol. Exp. Appl. 2009, 133, 260–275. [Google Scholar] [CrossRef]
- Zajac, B.K.; Sontigun, N.; Wannasan, A.; Verhoff, M.A.; Sukontason, K.; Amendt, J.; Zehner, R. Application of DNA barcoding for identifying forensically relevant Diptera from northern Thailand. Parasitol. Res. 2016, 115, 2307–2320. [Google Scholar] [CrossRef] [PubMed]
- Bharti, M.; Singh, B. DNA-based identification of forensically important blow flies (Diptera: Calliphoridae) from India. J. Med. Entomol. 2017, 54, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Ren, L.; Wang, Z.; Deng, J.; Guo, Y.; Chen, C.; Finkelbergs, D.; Cai, J. Identification of forensically important blow flies (Diptera: Calliphoridae) in China based on COI. J. Med. Entomol. 2017, 54, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, L.M.; Stevens, J.R. The molecular systematics of blowflies and screwworm flies (Diptera: Calliphoridae) using 28S rRNA, COX1 and EF-1α: Insights into the evolution of dipteran parasitism. Parsitology 2011, 138, 1760–1777. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kurahashi, H.; Wells, J.D. Molecular phylogeny of the blowfly genus Chrysomya. Med. Vet. Entomol. 2011, 25, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.A.; Lamb, J.; Villet, M.H. Phylogenetic radiation of the greenbottle flies (Diptera, Calliphoridae, Luciliinae). Zookeys 2016, 586, 59–86. [Google Scholar] [CrossRef] [PubMed]
- Saccone, C.; De Giorgi, C.; Gissi, C.; Pesole, G.; Reyes, A. Evolutionary genomics in Metazoa: The mitochondrial DNA as a model system. Gene 1999, 238, 195–209. [Google Scholar] [CrossRef]
- Sperling, F.A.; Anderson, G.S.; Hickey, D.A. A DNA-based approach to the identification of insect species used for postmortem interval estimation. J. Forensic Sci. 1994, 39, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Wallman, J.F.; Donnellan, S.C. The utility of mitochondrial DNA sequences for the identification of forensically important blowflies (Diptera: Calliphoridae) in southeastern Australia. Forensic Sci. Int. 2001, 120, 60–67. [Google Scholar] [CrossRef]
- Wells, J.D.; Sperling, F.A. DNA-based identification of forensically important Chrysomyinae (Diptera: Calliphoridae). Forensic Sci. Int. 2001, 120, 110–115. [Google Scholar] [CrossRef]
- Nelson, L.A.; Wallman, J.F.; Dowton, M. Using COI barcodes to identify forensically and medically important blowflies. Med. Vet. Entomol. 2007, 21, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, B.W.; Liu, T.T.; Fan, H.; Wei, D.; Wen, F.Q.; Bai, P.; Huang, J.; Hou, Y.P. The application of mitochondrial DNA cytochrome oxidase II gene for the identification of forensically important blowflies in western China. Am. J. Forensic Med. Pathol. 2007, 28, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, H.; Bunchu, N. The blow flies recorded from Thailand, with the description of a new species of Isomyia Walker (Diptera, Calliphoridae). Jpn. J. Syst. Entomol. 2011, 17, 237–278. [Google Scholar]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Tippanun, J.; Lertthamnongtham, S.; Vogtsberger, R.C.; Olson, J.K. Survey of forensically-relevant fly species in Chiang Mai, northern Thailand. J. Vector Ecol. 2003, 28, 135–138. [Google Scholar] [PubMed]
- Bunchu, N.; Sukontason, K.; Sanit, S.; Chidburee, P.; Kurahashi, H.; Sukontason, K.L. Occurrence of blow fly species (Diptera: Calliphoridae) in Phitsanulok province, northern Thailand. Trop. Biomed. 2012, 29, 532–543. [Google Scholar] [PubMed]
- Moophayak, K.; Klong-klaew, T.; Sukontason, K.; Kurahashi, H.; Tomberlin, J.K.; Sukontason, K.L. Species composition of carrion blow flies in northern Thailand: Altitude appraisal. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Klong-klaew, T.; Sontigun, N.; Sanit, S.; Samerjai, C.; Sukontason, K.; Kurahashi, H.; Koehler, P.G.; Pereira, R.M.; Limsopatham, K.; Suwannayod, S.; et al. Field evaluation of a semi-automatic funnel trap targeted the medically important non-biting flies. Acta Trop. 2017, 176, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Narongchai, P.; Kanchai, C.; Vichairat, K.; Sribanditmongkol, P.; Bhoopat, T.; Kurahashi, H.; Chockjamsai, M.; Piangjai, S.; Bunchu, N.; et al. Forensic entomology cases in Thailand: A review of cases from 2000 to 2006. Parasitol. Res. 2007, 101, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Monum, T.; Sukontason, K.L.; Sribanditmongkol, P.; Sukontason, K.; Samerjai, C.; Limsopatham, K.; Suwannayod, S.; Klong-klaew, T.; Wannasan, A. Forensically important blow flies Chrysomya pinguis, C. villeneuvi, and Lucilia porphyrina (Diptera: Calliphoridae) in a case of human remains in Thailand. Korean J. Parasitol. 2017, 55, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.L.; Bhoopat, T.; Wannasan, A.; Sontigun, N.; Sanit, S.; Amendt, J.; Samerjai, C.; Sukontason, K. Chrysomya chani Kurahashi (Diptera: Calliphoridae), a blow fly species of forensic importance: Morphological characters of the third larval instar and a case report from Thailand. Forensic Sci. Res. 2018, 3, 83–93. [Google Scholar] [CrossRef]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Boonchu, N.; Kurahashi, H.; Hope, M.; Olson, J.K. Identification of forensically important fly eggs using a potassium permanganate staining technique. Micron 2004, 35, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Sukontason, K.L.; Ngern-klun, R.; Sripakdee, D.; Piangjai, S. Differentiation of the third instar of forensically important fly species in Thailand. Ann. Entomol. Soc. Am. 2004, 97, 1069–1075. [Google Scholar] [CrossRef]
- Preativatanyou, K.; Sirisup, N.; Payungporn, S.; Poovorawan, Y.; Thavara, U.; Tawatsin, A.; Sungpradit, S.; Siriyasatien, P. Mitochondrial DNA-based identification of some forensically important blowflies in Thailand. Forensic Sci. Int. 2010, 202, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Bogdanowicz, S.M.; Wallner, W.E.; Bell, J.; Odell, T.M.; Harrison, R.G. Asian gypsy moths (Lepidoptera: Lymantriidae) in North America: Evidence from molecular data. Ann. Entomol. Soc. Am. 1993, 86, 710–715. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallman, J.F.; Leys, R.; Hogendoorn, K. Molecular systematics of Australian carrion-breeding blowflies (Diptera: Calliphoridae) based on mitochondrial DNA. Invertebr. Syst. 2005, 19, 1–15. [Google Scholar] [CrossRef]
- Guo, Y.D.; Cai, J.F.; Xiong, F.; Wang, H.J.; Wen, J.F.; Li, J.B.; Chen, Y.Q. The utility of mitochondrial DNA fragments for genetic identification of forensically important sarcophagid flies (Diptera: Sarcophagidae) in China. Trop. Biomed. 2012, 29, 51–60. [Google Scholar] [PubMed]
- Lessinger, A.C.; Junqueira, A.C.M.; Conte, F.F.; Azeredo-Espin, A.M.L. Analysis of a conserved duplicated tRNA gene in the mitochondrial genome of blow flies. Gene 2004, 339, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Collura, R.V.; Auerbach, M.R.; Stewart, C.B. A quick, direct method that can differentiate expressed mitochondrial genes from their nuclear pseudogenes. Curr. Biol. 1996, 6, 1337–1339. [Google Scholar] [CrossRef]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, L.A.R. Mitochondrial pseudogenes in insect DNA barcoding: Differing points of view on the same issue. Biota Neotrop. 2012, 12, 301–308. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Zhang, Y.; Piao, H.; Yu, D.H.; Jeong, H.J.; Yoo, G.Y.; Chung, U.; Jo, T.H.; Hwang, J.J. Use of cytochrome c oxidase subunit I (COI) nucleotide sequences for identification of the Korean Luciliinae fly species (Diptera: Calliphoridae) in forensic investigations. J. Korean Med. Sci. 2009, 24, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.M.; Adham, F.K.; Picard, C.J. Survey of the genetic diversity of forensically important Chrysomya (Diptera: Calliphoridae) from Egypt. J. Med. Entomol. 2015, 52, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.D.; Stevens, J.R. Application of DNA-based methods in forensic entomology. Annu. Rev. Entomol. 2008, 53, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Virgilio, M.; Jordaens, K.; Breman, F.C.; Backeljau, T.; De Meyer, M. Identifying insects with incomplete DNA barcode libraries, African fruit flies (Diptera: Tephritidae) as a test case. PLoS ONE 2012, 7, e31581. [Google Scholar] [CrossRef] [PubMed]
- Sonet, G.; Jordaens, K.; Braet, Y.; Bourguignon, L.; Dupont, E.; Backeljau, T.; De Meyer, M.; Desmyter, S. Utility of GenBank and the Barcode of Life Data Systems (BOLD) for the identification of forensically important Diptera from Belgium and France. Zookeys 2013, 365, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.M. Reliability of long vs short COI markers in identification of forensically important flies. Croat. Med. J. 2014, 55, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GilArriortua, M.; Saloña Bordas, M.I.; Köhnemann, S.; Pfeiffer, H.; de Pancorbo, M.M. Molecular differentiation of Central European blowfly species (Diptera, Calliphoridae) using mitochondrial and nuclear genetic markers. Forensic Sci. Int. 2014, 242, 274–282. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Sequences | No. of Polymorphic Sites | No. of Haplotypes | Intraspecific Divergence (Minimum–Maximum) |
|---|---|---|---|---|
| C. bezziana | 1 | N/A | 1 | N/A |
| C. megacephala | 15 | 8 | 4 | 0.1 (0.0–0.6) |
| C. chani | 8 | 6 | 5 | 0.1 (0.0–0.4) |
| C. pinguis | 7 | 14 | 7 | 0.4 (0.2–0.6) |
| C. thanomthini | 2 | 1 | 2 | 0.1 |
| C. nigripes | 4 | 4 | 4 | 0.2 (0.1–0.2) |
| C. rufifacies | 14 | 5 | 6 | 0.1 (0.0–0.2) |
| C. villeneuvi | 14 | 8 | 6 | 0.1 (0.0–0.4) |
| L. cuprina | 8 | 31 | 5 | 0.7 (0.0–2.4) |
| L. papuensis | 9 | 12 | 5 | 0.5 (0.0–0.8) |
| L. porphyrina | 6 | 23 | 6 | 0.7 (0.2–1.1) |
| L. sinensis | 5 | 11 | 4 | 0.4 (0.0–0.8) |
| H. ligurriens | 10 | 6 | 6 | 0.1 (0.0–0.4) |
| H. pulchra | 4 | 6 | 4 | 0.3 (0.2–0.4) |
| H. infumata | 3 | 4 | 3 | 0.2 (0.2–0.3) |
| H. tumrasvini | 3 | 2 | 2 | 0.1 (0.0–0.2) |
| Species | No. of Sequences | No. of Polymorphic Sites | No. of Haplotypes | Intraspecific Divergence (Minimum–Maximum) |
|---|---|---|---|---|
| C. bezziana | 1 | N/A | 1 | N/A |
| C. megacephala | 13 | 0 | 1 | 0.0 |
| C. chani | 8 | 0 | 1 | 0.0 |
| C. pinguis | 7 | 3 | 4 | 0.2 (0.0–0.3) |
| C. thanomthini | 2 | 1 | 2 | 0.2 |
| C. nigripes | 3 | 4 | 3 | 0.4 (0.3–0.6) |
| C. rufifacies | 14 | 4 | 4 | 0.1 (0.0–0.5) |
| C. villeneuvi | 14 | 2 | 3 | 0.1 (0.0–0.3) |
| L. cuprina | 8 | 1 | 2 | 0.0 (0.0–0.2) |
| L. papuensis | 9 | 5 | 3 | 0.4 (0.0–0.8) |
| L. porphyrina | 6 | 14 | 5 | 0.8 (0.0–1.9) |
| L. sinensis | 5 | 5 | 3 | 0.3 (0.0–0.8) |
| H. pulchra | 4 | 2 | 3 | 0.2 (0.0–0.3) |
| H. infumata | 3 | 1 | 2 | 0.1 (0.0–0.2) |
| H. tumrasvini | 3 | 1 | 2 | 0.1 (0.0–0.2) |
| No. | Species | Interspecific Distance | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | ||
| 1 | C. bezziana | - | 3.6 | 5.1 | 3.8 | 3.3 | 5.0 | 6.3 | 6.6 | 9.2 | 10.9 | 9.6 | 11.1 | - | 9.4 | 10.0 | 9.4 |
| 2 | C. megacephala | 4.9 | - | 4.4 | 1.8 | 2.2 | 5.3 | 6.4 | 6.3 | 9.6 | 10.6 | 9.3 | 11.2 | - | 10.1 | 10.8 | 8.5 |
| 3 | C. chani | 5.6 | 4.5 | - | 3.9 | 4.8 | 5.3 | 5.8 | 6.5 | 8.9 | 10.0 | 9.7 | 10.7 | - | 7.6 | 8.6 | 7.0 |
| 4 | C. pinguis | 5.4 | 2.6 | 4.3 | - | 2.5 | 5.5 | 6.9 | 7.2 | 8.7 | 10.6 | 8.7 | 11.1 | - | 8.9 | 10.0 | 8.4 |
| 5 | C. thanomthini | 5.6 | 3.4 | 4.5 | 2.8 | - | 6.1 | 6.6 | 6.9 | 9.7 | 10.9 | 9.4 | 11.3 | - | 9.6 | 10.9 | 9.3 |
| 6 | C. nigripes | 6.6 | 5.4 | 5.9 | 6.2 | 6.2 | - | 6.3 | 6.3 | 10.0 | 11.6 | 10.5 | 11.4 | - | 8.4 | 10.1 | 9.1 |
| 7 | C. rufifacies | 8.1 | 7.0 | 6.8 | 7.9 | 7.9 | 7.9 | - | 3.4 | 9.6 | 10.3 | 9.3 | 10.5 | - | 8.4 | 9.7 | 8.3 |
| 8 | C. villeneuvi | 7.6 | 6.7 | 7.4 | 7.8 | 7.5 | 7.7 | 4.7 | - | 9.6 | 10.8 | 9.5 | 9.9 | - | 8.7 | 9.7 | 8.8 |
| 9 | L. cuprina | 9.6 | 8.8 | 7.9 | 9.6 | 9.0 | 9.7 | 10.3 | 10.1 | - | 9.4 | 5.7 | 7.0 | - | 5.7 | 6.3 | 4.8 |
| 10 | L. papuensis | 10.5 | 10.7 | 8.3 | 10.7 | 10.1 | 9.9 | 10.7 | 10.0 | 6.7 | - | 8.7 | 6.0 | - | 9.6 | 9.9 | 7.5 |
| 11 | L. porphyrina | 11.1 | 10.2 | 9.2 | 10.4 | 9.8 | 11.0 | 10.7 | 10.4 | 6.2 | 7.8 | - | 8.0 | - | 7.4 | 7.1 | 6.3 |
| 12 | L. sinensis | 8.7 | 8.7 | 8.6 | 9.5 | 9.1 | 9.4 | 10.1 | 9.1 | 5.3 | 5.9 | 5.4 | - | - | 8.9 | 8.9 | 5.8 |
| 13 | H. ligurriens | 9.7 | 9.0 | 9.0 | 10.0 | 9.7 | 9.9 | 10.5 | 10.0 | 6.1 | 8.6 | 7.5 | 6.1 | - | - | - | - |
| 14 | H. pulchra | 9.3 | 9.0 | 8.4 | 9.9 | 9.4 | 9.7 | 10.2 | 9.6 | 6.1 | 8.3 | 7.2 | 5.9 | 1.1 | - | 4.2 | 6.1 |
| 15 | H. infumata | 9.4 | 10.2 | 9.4 | 10.0 | 10.3 | 10.0 | 11.0 | 10.3 | 7.6 | 9.0 | 9.0 | 7.1 | 5.6 | 5.5 | - | 5.7 |
| 16 | H. tumrasvini | 9.8 | 9.5 | 8.5 | 10.4 | 10.1 | 9.7 | 9.6 | 10.0 | 4.9 | 6.8 | 7.0 | 5.1 | 6.0 | 5.8 | 7.6 | - |
| Criteria | COI | COII |
|---|---|---|
| No. of sequences | 114 | 101 |
| No. of species | 16 | 15 |
| No. of sequences with closest match at 0% difference | 57 | 80 |
| No. of allospecific matches at 0% difference | 0 | 0 |
| Best Match (BM) | ||
| - Correct identifications | 100.00% (114) | 100.00% (101) |
| - Ambiguous identifications | 0.00% (0) | 0.00% (0) |
| - Incorrect identifications | 0.00% (0) | 0.00% (0) |
| Calculated threshold for Best Close Match and All Species Barcodes | 0.72% | 0.63% |
| Best Close Match (BCM) | ||
| - Correct identifications | 98.24% (112) | 98.01% (99) |
| - Ambiguous identifications | 0.00% (0) | 0.00% (0) |
| - Incorrect identifications | 0.00% (0) | 0.00% (0) |
| - No match closer than the calculated threshold | 1.75% (2) | 1.98% (2) |
| All Species Barcodes (ASB) | ||
| - Correct identifications | 94.73% (108) | 94.05% (95) |
| - Ambiguous identifications | 3.50% (4) | 3.96% (4) |
| - Incorrect identifications | 0.00% (0) | 0.00% (0) |
| - No match closer than the calculated threshold | 1.75% (2) | 1.98% (2) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sontigun, N.; Sukontason, K.L.; Amendt, J.; Zajac, B.K.; Zehner, R.; Sukontason, K.; Chareonviriyaphap, T.; Wannasan, A. Molecular Analysis of Forensically Important Blow Flies in Thailand. Insects 2018, 9, 159. https://doi.org/10.3390/insects9040159
Sontigun N, Sukontason KL, Amendt J, Zajac BK, Zehner R, Sukontason K, Chareonviriyaphap T, Wannasan A. Molecular Analysis of Forensically Important Blow Flies in Thailand. Insects. 2018; 9(4):159. https://doi.org/10.3390/insects9040159
Chicago/Turabian StyleSontigun, Narin, Kabkaew L. Sukontason, Jens Amendt, Barbara K. Zajac, Richard Zehner, Kom Sukontason, Theeraphap Chareonviriyaphap, and Anchalee Wannasan. 2018. "Molecular Analysis of Forensically Important Blow Flies in Thailand" Insects 9, no. 4: 159. https://doi.org/10.3390/insects9040159
APA StyleSontigun, N., Sukontason, K. L., Amendt, J., Zajac, B. K., Zehner, R., Sukontason, K., Chareonviriyaphap, T., & Wannasan, A. (2018). Molecular Analysis of Forensically Important Blow Flies in Thailand. Insects, 9(4), 159. https://doi.org/10.3390/insects9040159