From Exit to Entry: Long-term Survival and Transmission of Salmonella


Abstract
:1. Introduction
1.1. Salmonella Taxonomy and Human Disease
2. Results and Discussion
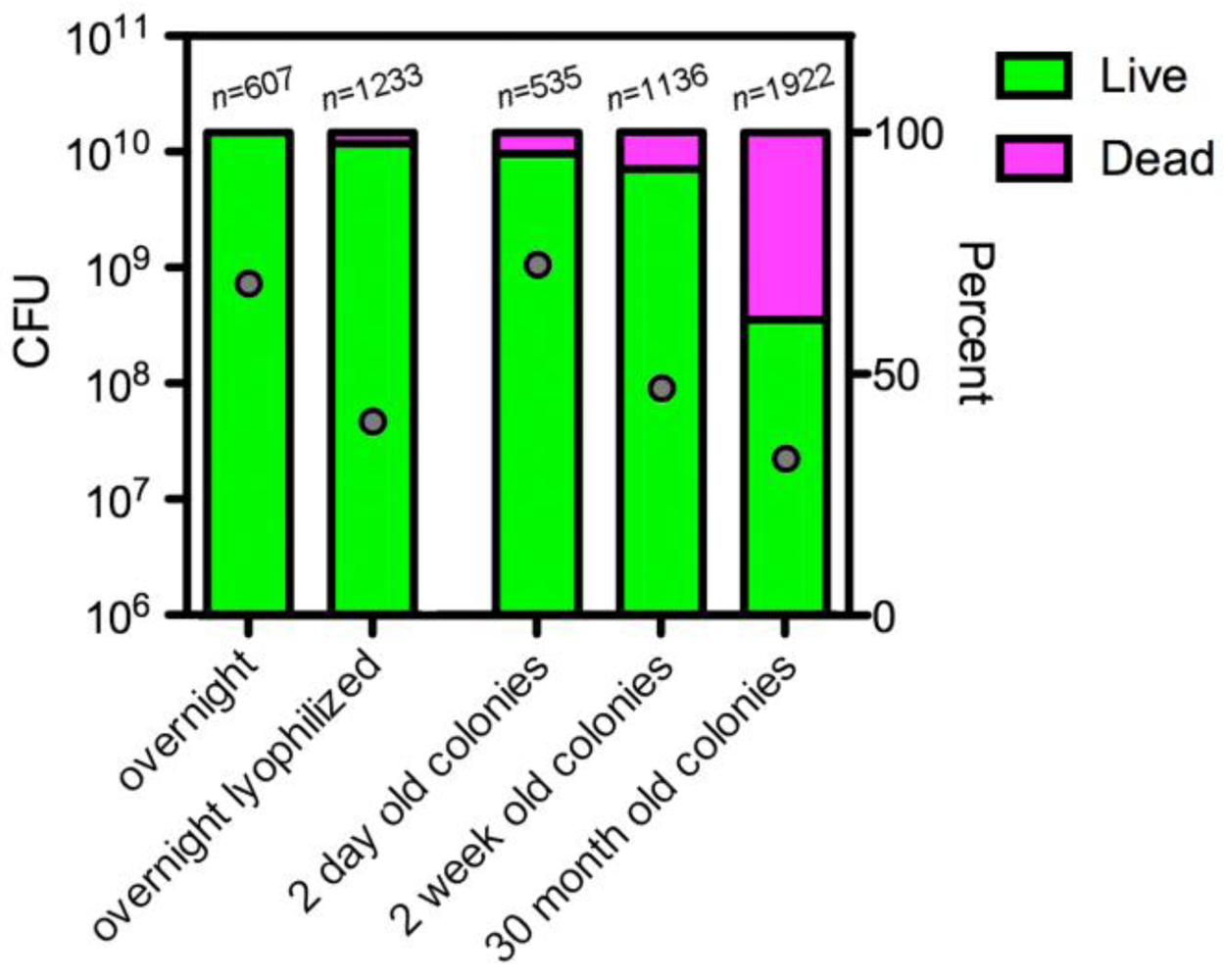
2.1. In vitro Evidence that Salmonella spp. Can Survive for Long Periods of Time under Harsh Conditions
2.1.1. Does Salmonella Enter a Viable, Non-culturable State?

2.2. Salmonella in the Environment: Lessons from Outbreaks
2.2.1. Tomato-related Salmonella Outbreaks

{kind=link}
| Food Vehicle | S. enterica Serovar | Cases | Year | Traceback | Reference |
|---|---|---|---|---|---|
| Tomato | Javiana | 176 | 1990 | Packing plant | [67] |
| Tomato | Montevideo | 100 | 1993 | Packing plant | [67] |
| Tomato | Baildon | 86 | 1998 | Grower/Packing plant | [68] |
| Tomato | Newport | 510 | 2002 | Pond water at grower | [69] |
| Tomato | Braenderup | 125 | 2004 | Packing plant | [70,71] |
| Tomato | Multiserotype | 561 | 2004 | Packing plant | [70,72] |
| Tomato | Braenderup | 82 | 2005 | Grower | [73] |
| Tomato | Newport | 72 | 2005 | Pond water at grower | [69,73] |
| Tomato | Typhimurium | 190 | 2006 | Packing plant | [73,74] |
| Tomato | Newport | 115 | 2006 | Not stated | [73] |
| Alfalfa sprouts | Stanley | >242 | 1995 | Alfalfa Seeds | [75] |
| Alfalfa sprouts | Newport | >133 | 1995 | Alfalfa Seeds | [76] |
| Alfalfa sprouts | Montevideo | 417 | 1996 | Alfalfa Seeds | [77] |
| Alfalfa sprouts | Melegridis | 75 | 1996 | Alfalfa Seeds | [77] |
| Alfalfa/Clover | Senftenberg | 60 | 1997 | Clover Seeds | [77] |
| Alfalfa sprouts | Infantis/Anatum | 109 | 1997 | Alfalfa Seeds | [78] |
| Alfalfa sprouts | Havana | 18 | 1998 | Alfalfa Seeds | [77] |
| Alfalfa sprouts | Cubana | 22 | 1998 | Alfalfa Seeds | [77] |
| Alfalfa sprouts | Mbandaka | 87 | 1999 | Alfalfa Seeds | [79] |
| Alfalfa sprouts | Muenchen | 157 | 1999 | Alfalfa Seeds | [80] |
| Clover sprouts | Typhimurium | 112 | 1999 | Clover seeds | [81] |
| Alfalfa sprouts | Kottbus | 31 | 2003 | Alfalfa seeds | [82] |
| Alfalfa sprouts | Saintpaul | 228 | 2009 | Alfalfa Seeds | [83] |
| Alfalfa sprouts | Newport | 44 | 2010 | Not Stated | [84] |
| Alfalfa/Clover | Typhimuirum (I 4,[5],12:i:) | 140 | 2010 | Not stated | [85] |
| Alfalfa sprouts | Enteriditis | 25 | 2011 | Not stated | [86] |
| Paprika Chips | Saintpaul, Javiana, and Rubislaw | >670 | 1993 | Paprika powder | [87] |
| Toasted Oat Cereal | Agona | 209 | 1998 | Air-handling systemand vitamin spray mixer | [88] |
| Squid/shrimp Crackers | Chester and Oranienberg | 1634 | 1999 | Unknown | [89,90] |
| Chocolate | Durham | 110 | 1970 | Cocoa powder | [91] |
| Chocolate | Eastbourne | 217 | 1973 | Cocoa bean | [92,93] |
| Chocolate | Napoli | 245 | 1982 | Not stated | [94] |
| Chocolate | Nima | 24 | 1985 | Not stated | [95] |
| Chocolate | Typhimurium | 349 | 1987 | Avian wildlife reservoir suspected | [96] |
| Chocolate | Oranienburg | 439 | 2001 | Unknown | [97] |
2.2.3. Salmonella Outbreaks Associated with Processed Foods
2.3. Genomic—and Population—based Studies of Salmonella and Related Enteric Pathogens
3. Commentary: Infectious NTS Isolates in Africa
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Blaser, M.J.; Kirschner, D. The equilibria that allow bacterial persistence in human hosts. Nature 2007, 449, 843–849. [Google Scholar] [CrossRef]
- Savageau, M.A. Escherichia coli habitats, cell types, and molecular mechanisms of gene control. Am. Nat. 1983, 122, 732–744. [Google Scholar]
- White, A.P.; Sibley, K.A.; Sibley, C.D.; Wasmuth, J.D.; Schaefer, R.; Surette, M.G.; Edge, T.A.; Neumann, N.F. Intergenic sequence comparison of Escherichia coli isolates reveals lifestyle adaptations but not host specificity. Appl. Environ. Microbiol. 2011, 77, 7620–7632. [Google Scholar] [CrossRef]
- Santamaria, J.; Toranzos, G.A. Enteric pathogens and soil: A short review. Int. Microbiol. 2003, 6, 5–9. [Google Scholar]
- Le Minor, L.; Popoff, M.Y. Designation of Salmonella enterica sp. Nov., nom. Rev., as the type and only species of the genus Salmonella: Request for an opinion. Int. J. Syst. Bacteriol. 1987, 37, 465–468. [Google Scholar] [CrossRef]
- Reeves, M.W.; Evins, G.M.; Heiba, A.A.; Plikaytis, B.D.; Farmer Iii, J.J. Clonal nature of Salmonella typhi and its genetic relatdeness to other salmonellae as shown by multilocus enzyme electrophoresis, and proposal of Salmonella bongori comb. Nov. J. Clin. Microbiol. 1989, 27, 313–320. [Google Scholar]
- Grimont, P.A.D.; Weill, F.X. Antigenic formulae of the Salmonella serovars, 9th ed.; WHO Collaborating Center for Reference and Research on Salmonella. Institut Pasteur: Paris, France, 2007. [Google Scholar]
- Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Byrd, J.A.; Nisbet, D.J. Gastrointestinal microbial ecology and the safety of our food supply as related to Salmonella. J. Anim. Sci. 2007, 86, 163–172. [Google Scholar] [CrossRef]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-saharan Africa have a distinct genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive nontyphoidal Salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O'Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- Crump, J.A.; Mintz, E.D. Global trends in typhoid and paratyphoid fever. Clin. Infect. Dis. 2010, 50, 241–246. [Google Scholar] [CrossRef]
- Voetsch, A.C.; Van Gilder, T.J.; Angulo, F.J.; Farley, M.M.; Shallow, S.; Marcus, R.; Cieslak, P.R.; Deneen, V.C.; Tauxe, R.V. Foodnet estimate of the burden of illness caused by nontyphoidal Salmonella infections in the united states. Clin. Infect. Dis. 2004, 38 Suppl. 3, S127–S134. [Google Scholar]
- Betancor, L.; Yim, L.; Martinez, A.; Fookes, M.; Sasias, S.; Schelotto, F.; Thomson, N.; Maskell, D.; Chabalgoity, J.A. Genomic comparison of the closely related Salmonella enterica serovars Enteritidis and Dublin. Open Microbiol. J. 2012, 6, 5–13. [Google Scholar] [CrossRef]
- Gonzalez-Escobedo, G.; Marshall, J.M.; Gunn, J.S. Chronic and acute infection of the gall bladder by Salmonella Typhi: Understanding the carrier state. Nat. Rev. Microbiol. 2011, 9, 9–14. [Google Scholar] [CrossRef]
- Levine, M.M.; Black, R.E.; Lanata, C. Precise estimation of the numbers of chronic carriers of Salmonella typhi in Santiago, Chile, an endemic area. J. Infect. Dis. 1982, 146, 724–726. [Google Scholar] [CrossRef]
- Stokes, A.; Clarke, C. A search for typhoid carriers among 800 convalescents. Lancet 1916, 187, 566–569. [Google Scholar] [CrossRef]
- Vogelsang, T.M.; Boe, J. Temporary and chronic carriers of Salmonella typhi and Salmonella paratyphi B. J. Hyg. 1948, 46, 252–261. [Google Scholar] [CrossRef]
- Buchwald, D.S.; Blaser, M.J. A review of human salmonellosis: Ii. Duration of excretion following infection with nontyphi Salmonella. Rev. Infect. Dis. 1984, 6, 345–356. [Google Scholar] [CrossRef]
- Winfield, M.D.; Groisman, E.A. Role of nonhost environments in the lifestyles of Salmonella and Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 3687–3694. [Google Scholar] [CrossRef]
- Spector, M.P.; Kenyon, W.J. Resistance and survival strategies of Salmonella enterica to environmental stresses. Food Res. Int. 2012, 45, 455–481. [Google Scholar] [CrossRef]
- Davies, R.H.; Breslin, M. Persistence of Salmonella enteritidis phage type 4 in the environment and arthropod vectors on an empty free-range chicken farm. Environ. Microbiol. 2003, 5, 79–84. [Google Scholar] [CrossRef]
- Snow, L.C.; Davies, R.H.; Christiansen, K.H.; Carrique-Mas, J.J.; Cook, A.J.; Evans, S.J. Investigation of risk factors for Salmonella on commercial egg-laying farms in Great Britain, 2004-2005. Vet. Rec. 2010, 166, 579–586. [Google Scholar] [CrossRef]
- Nuccio, S.P.; Thomson, N.R.; Fookes, M.C.; Baumler, A.J. Fimbrial signature arrangements in salmonella. In Salmonella: From genome to function; Caister Academic Press: Norfolk, UK, 2011; pp. 149–161. [Google Scholar]
- Baumler, A.J.; Gilde, A.J.; Tsolis, R.M.; van der Velden, A.W.; Ahmer, B.M.; Heffron, F. Contribution of horizontal gene transfer and deletion events to development of distinctive patterns of fimbrial operons during evolution of Salmonella serotypes. J. Bacteriol. 1997, 179, 317–322. [Google Scholar]
- Doran, J.L.; Collinson, S.K.; Burian, J.; Sarlos, G.; Todd, E.C.; Munro, C.K.; Kay, C.M.; Banser, P.A.; Peterkin, P.I.; Kay, W.W. DNA-based diagnostic tests for Salmonella species targeting agfA, the structural gene for thin, aggregative fimbriae. J. Clin. Microbiol. 1993, 31, 2263–2273. [Google Scholar]
- Collinson, S.K.; Emody, L.; Muller, K.H.; Trust, T.J.; Kay, W.W. Purification and characterization of thin, aggregative fimbriae from Salmonella enteritidis. J. Bacteriol. 1991, 173, 4773–4781. [Google Scholar]
- Collinson, S.K.; Doig, P.C.; Doran, J.L.; Clouthier, S.; Trust, T.J.; Kay, W.W. Thin, aggregative fimbriae mediate binding of Salmonella enteritidis to fibronectin. J. Bacteriol. 1993, 175, 12–18. [Google Scholar]
- Collinson, S.K.; Emody, L.; Trust, T.J.; Kay, W.W. Thin aggregative fimbriae from diarrheagenic Escherichia coli. J. Bacteriol. 1992, 174, 4490–4495. [Google Scholar]
- Romling, U.; Sierralta, W.D.; Eriksson, K.; Normark, S. Multicellular and aggregative behaviour of Salmonella typhimurium strains is controlled by mutations in the agfD promoter. Mol. Microbiol. 1998, 28, 249–264. [Google Scholar] [CrossRef]
- Romling, U.; Bian, Z.; Hammar, M.; Sierralta, W.D.; Normark, S. Curli fibers are highly conserved between Salmonella typhimurium and Escherichia coli with respect to operon structure and regulation. J. Bacteriol. 1998, 180, 722–731. [Google Scholar]
- White, A.P.; Gibson, D.L.; Collinson, S.K.; Banser, P.A.; Kay, W.W. Extracellular polysaccharides associated with thin aggregative fimbriae of Salmonella enterica serovar Enteritidis. J. Bacteriol. 2003, 185, 5398–5407. [Google Scholar] [CrossRef]
- Romling, U.; Rohde, M.; Olsen, A.; Normark, S.; Reinkoster, J. AgfD, the checkpoint of multicellular and aggregative behaviour in Salmonella typhimurium regulates at least two independent pathways. Mol. Microbiol. 2000, 36, 10–23. [Google Scholar] [CrossRef]
- Gibson, D.L.; White, A.P.; Snyder, S.D.; Martin, S.; Heiss, C.; Azadi, P.; Surette, M.; Kay, W.W. Salmonella produces an o-antigen capsule regulated by AgfD and important for environmental persistence. J. Bacteriol. 2006, 188, 7722–7730. [Google Scholar] [CrossRef]
- de Rezende, C.E.; Anriany, Y.; Carr, L.E.; Joseph, S.W.; Weiner, R.M. Capsular polysaccharide surrounds smooth and rugose types of Salmonella enterica serovar Typhimurium DT104. Appl. Environ. Microbiol. 2005, 71, 7345–7351. [Google Scholar] [CrossRef]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.; Gamazo, C.; Penades, J.R.; Lasa, I. Bapa, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol. Microbiol. 2005, 58, 1522–1539. [Google Scholar]
- Chapman, M.R.; Robinson, L.S.; Pinkner, J.S.; Roth, R.; Heuser, J.; Hammar, M.; Normark, S.; Hultgren, S.J. Role of Escherichia coli curli operons in directing amyloid fiber formation. Science 2002, 295, 851–855. [Google Scholar]
- Collinson, S.K.; Parker, J.M.; Hodges, R.S.; Kay, W.W. Structural predictions of AgfA, the insoluble fimbrial subunit of Salmonella thin aggregative fimbriae. J. Mol. Biol. 1999, 290, 741–756. [Google Scholar] [CrossRef]
- Larsen, P.; Nielsen, J.L.; Dueholm, M.S.; Wetzel, R.; Otzen, D.; Nielsen, P.H.r. Amyloid adhesins are abundant in natural biofilms. Environ. Microbiol. 2007, 9, 3077–3090. [Google Scholar] [CrossRef]
- Anriany, Y.A.; Weiner, R.M.; Johnson, J.A.; De Rezende, C.E.; Joseph, S.W. Salmonella enterica serovar Typhimurium DT104 displays a rugose phenotype. Appl. Environ. Microbiol. 2001, 67, 4048–4056. [Google Scholar]
- Scher, K.; Romling, U.; Yaron, S. Effect of heat, acidification, and chlorination on Salmonella enterica serovar Typhimurium cells in a biofilm formed at the air-liquid interface. Appl. Environ. Microbiol. 2005, 71, 1163–1168. [Google Scholar] [CrossRef]
- Solano, C.; Garcia, B.; Valle, J.; Berasain, C.; Ghigo, J.M.; Gamazo, C.; Lasa, I. Genetic analysis of Salmonella enteritidis biofilm formation: Critical role of cellulose. Mol. Microbiol. 2002, 43, 793–808. [Google Scholar] [CrossRef]
- White, A.P.; Gibson, D.L.; Kim, W.; Kay, W.W.; Surette, M.G. Thin aggregative fimbriae and cellulose enhance long-term survival and persistence of Salmonella. J. Bacteriol. 2006, 188, 3219–3227. [Google Scholar]
- Stocki, S.L.; Annett, C.B.; Sibley, C.D.; McLaws, M.; Checkley, S.L.; Singh, N.; Surette, M.G.; White, A.P. Persistence of Salmonella on egg conveyor belts is dependent on the belt type but not on the rdar morphotype. Poult. Sci. 2007, 86, 2375–2383. [Google Scholar] [CrossRef]
- Uhlich, G.A.; Cooke, P.H.; Solomon, E.B. Analyses of the red-dry-rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents. Appl. Environ. Microbiol. 2006, 72, 2564–2572. [Google Scholar]
- Austin, J.W.; Sanders, G.; Kay, W.W.; Collinson, S.K. Thin aggregative fimbriae enhance Salmonella enteritidis biofilm formation. FEMS Microbiol. Lett. 1998, 162, 295–301. [Google Scholar] [CrossRef]
- Ryu, J.H.; Beuchat, L.R. Biofilm formation by Escherichia coli O157:H7 on stainless steel: Effect of exopolysaccharide and curli production on its resistance to chlorine. Appl. Environ. Microbiol. 2005, 71, 247–254. [Google Scholar] [CrossRef]
- Apel, D.; White, A.P.; Grassl, G.A.; Finlay, B.B.; Surette, M.G. Long-term survival of Salmonella enterica serovar Typhmurium reveals an Infect. state that is underrepresented on laboratory media containing bile salts. Appl. Environ. Microbiol. 2009, 75, 4923–4925. [Google Scholar] [CrossRef]
- White, A.P.; Gibson, D.L.; Grassl, G.A.; Kay, W.W.; Finlay, B.B.; Vallance, B.A.; Surette, M.G. Aggregation via the red, dry, and rough morphotype is not a virulence adaptation in Salmonella enterica serovar Typhimurium. Infect. Immun. 2008, 76, 1048–1058. [Google Scholar] [CrossRef]
- Knudsen, G.M.; Nielsen, M.-B.; Grassby, T.; Danino-Appleton, V.; Thomsen, L.E.; Colquhoun, I.J.; Brocklehurst, T.F.; Olsen, J.E.; Hinton, J.C.D. A third mode of surface-associated growth: Immobilization of Salmonella enterica serovar Typhimurium modulates the RpoS-directed transcriptional programme. Environ.Microbiol 2012, 14, 1855–1875. [Google Scholar] [CrossRef]
- White, A.P.; Weljie, A.M.; Apel, D.; Zhang, P.; Shaykhutdinov, R.; Vogel, H.J.; Surette, M.G. A global metabolic shift is linked to Salmonella multicellular development. PLoS One 2010, 5, e11814. [Google Scholar] [CrossRef]
- Grantcharova, N.; Peters, V.; Monteiro, C.; Zakikhany, K.; Romling, U. Bistable expression of csgD in biofilm development of Salmonella enterica serovar Typhimurium. J. Bacteriol. 2010, 192, 456–466. [Google Scholar] [CrossRef]
- Siebring, J.; Sorg, R.A.; Herber, M.; Kuipers, O.P. Take it or leave it: Mechanisms underlying bacterial bistable regulatory networks. In Bacterial Regulatory Networks; Filloux, A.A.M., Ed.; Caister Academic Press: Norfolk, UK, 2012; pp. 305–332. [Google Scholar]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Gunn, J.S. Mechanisms of bacterial resistance and response to bile. Microbes Infect. 2000, 2, 907–913. [Google Scholar] [CrossRef]
- Roszak, D.B.; Grimes, D.J.; Colwell, R.R. Viable but nonrecoverable stage of Salmonella enteritidis in aquatic systems. Can. J. Microbiol. 1984, 30, 334–338. [Google Scholar] [CrossRef]
- Gupte, A.R.; de Rezende, C.L.E.; Joseph, S.W. Induction and resuscitation of viable but nonculturable Salmonella enterica serovar Typhimurium DT104. Appl. Environ. Microbiol. 2003, 69, 6669–6675. [Google Scholar]
- Panutdaporn, N.; Kawamoto, K.; Asakura, H.; Makino, S.I. Resuscitation of the viable but non-culturable state of Salmonella enterica serovar Oranienburg by recombinant resuscitation-promoting factor derived from Salmonella Typhimurium strain LT2. Int. J. Food Microbiol. 2006, 106, 241–247. [Google Scholar] [CrossRef]
- Reissbrodt, R.; Heier, H.; Tschape, H.; Kingsley, R.A.; Williams, P.H. Resuscitation by ferrioxamine e of stressed Salmonella enterica serovar Typhimurium from soil and water microcosms. Appl. Environ. Microbiol. 2000, 66, 4128–4130. [Google Scholar] [CrossRef]
- Reissbrodt, R.; Rienaecker, I.; Romanova, J.M.; Freestone, P.P.E.; Haigh, R.D.; Lyte, M.; Tschape, H.; Williams, P.H. Resuscitation of Salmonella enterica serovar Typhimurium and enterohemorrhagic Escherichia coli from the viable but nonculturable state by heat-stable enterobacterial autoinducer. Appl. Environ. Microbiol. 2002, 68, 4788–4794. [Google Scholar] [CrossRef]
- Caro, A.; Got, P.; Baleux, B. Physiological changes of Salmonella typhimurium cells under osmotic and starvation conditions by image analysis. FEMS Microbiol. Lett. 1999, 179, 265–273. [Google Scholar] [CrossRef]
- Lesne, J.; Berthet, S.; Binard, S.; Rouxel, A.; Humbert, F. Changes in culturability and virulence of Salmonella typhimurium during long-term starvation under desiccating conditions. Int. J. Food Microbiol. 2000, 60, 195–203. [Google Scholar] [CrossRef]
- Smith, R.J.; Newton, A.T.; Harwood, C.R.; Barer, M.R. Active but nonculturable cells of Salmonella enterica serovar Typhimurium do not infect or colonize mice. Microbiol. 2002, 148, 2717–2717. [Google Scholar]
- Kamruzzaman, M.; Udden, S.M.N.; Cameron, D.E.; Calderwood, S.B.; Nair, G.B.; Mekalanos, J.J.; Faruque, S.M. Quorum-regulated biofilms enhance the development of conditionally viable, environmental Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2010, 107, 1588–1593. [Google Scholar]
- CDC. Vital signs: Incidence and trends of infection with pathogens transmitted commonly through food-foodborne diseases active surveillance network, 10 U.S. Sites, 1996-2010. MMWR 2011, 60, 749–755. [Google Scholar]
- CDC. Preliminary foodnet data on the incidence of infection with pathogens transmitted commonly through food, 10 states. MMWR 2009, 59, 418–422. [Google Scholar]
- Hedberg, C.W.; Angulo, F.J.; White, K.E.; Langkop, C.W.; Schell, W.L.; Stobierski, M.G.; Schuchat, A.; Besser, J.M.; Dietrich, S.; et al. Outbreaks of salmonellosis associated with eating uncooked tomatoes: Implications for public health. Epidemiol. Infect. 1999, 122, 385–393. [Google Scholar] [CrossRef]
- Cummings, K.; Barrett, E.; Mohle-Boetani, J.C.; Brooks, J.T.; Farrar, J.; Hunt, T.; Fiore, A.; Komatsu, K.; Werner, S.B.; Slutsker, L. A multistate outbreak of Salmonella enterica serotype Baildon associated with domestic raw tomatoes. Emerg. Infect. Dis. 2001, 7, 1046–1048. [Google Scholar] [CrossRef]
- Greene, S.K.; Daly, E.R.; Talbot, E.A.; Demma, L.J.; Holzbauer, S.; Patel, N.J.; Hill, T.A.; Walderhaug, M.O.; Hoekstra, R.M.; Lynch, M.F.; Painter, J.A. Recurrent multistate outbreak of Salmonella newport associated with tomatoes from contaminated fields, 2005. Epidemiol. Infect. 2008, 136, 157–165. [Google Scholar]
- CDC. Outbreaks of Salmonella infections associated with eating roma tomatoes-united states and canada, 2004. MMWR 2005, 54, 325–328. [Google Scholar]
- Gupta, S.K.; Nalluswami, K.; Snider, C.; Perch, M.; Balasegaram, M.; Burmeister, D.; Lockett, J.; Sandt, C.; Hoekstra, R.M.; Montgomery, S. Outbreak of Salmonella Braenderup infections associated with roma tomatoes, northeastern United States, 2004: A useful method for subtyping exposures in field investigations. Epidemiol. Infect. 2007, 135, 1165–1173. [Google Scholar]
- Sandt, C.H.; Krouse, D.A.; Cook, C.R.; Hackman, A.L.; Chmielecki, W.A.; Warren, N.G. The key role of pulsed-field gel electrophoresis in investigation of a large multiserotype and multistate food-borne outbreak of Salmonella infections centered in Pennsylvania. J. Clin. Microbiol. 2006, 44, 3208–3212. [Google Scholar] [CrossRef]
- CDC. Multistate outbreaks of Salmonella infections associated with raw tomatoes eaten in restaurants--United States, 2005-2006. MMWR 2007, 56, 909–911. [Google Scholar]
- Behravesh, C.B.; Blaney, D.; Medus, C.; Bidol, S.A.; Phan, Q.; Soliva, S.; Daly, E.R.; Smith, K.; Miller, B.; Taylor, T.; et al. Multistate outbreak of Salmonella serotype Typhimurium infections associated with consumption of restaurant tomatoes, USA, 2006: Hypothesis generation through case exposures in multiple restaurant clusters. Epidemiol. Infect. 2012, 140, 2053–2061. [Google Scholar] [CrossRef]
- Mahon, B.E.; Ponka, A.; Hall, W.N.; Komatsu, K.; Dietrich, S.E.; Siitonen, A.; Cage, G.; Hayes, P.S.; Lambert-Fair, M.A.; Bean, N.H.; Griffin, P.M.; Slutsker, L. An international outbreak of Salmonella infections caused by alfalfa sprouts grown from contaminated seeds. J. Infect. Dis. 1997, 175, 876–882. [Google Scholar] [CrossRef]
- Van Beneden, C.A.; Keene, W.E.; Strang, R.A.; Werker, D.H.; King, A.S.; Mahon, B.; Hedberg, K.; Bell, A.; Kelly, M.T.; Balan, V.K.; Mac Kenzie, W.R.; Fleming, D. Multinational outbreak of Salmonella enterica serotype Newport infections due to contaminated alfalfa sprouts. J. Am. Med. Assoc. 1999, 281, 158–162. [Google Scholar]
- Mohle-Boetani, J.C.; Farrar, J.A.; Werner, S.B.; Minassian, D.; Bryant, R.; Abbott, S.; Slutsker, L.; Vugia, D.J. Escherichia coli O157 and Salmonella infections associated with sprouts in California, 1996-1998. Ann. Int. Med. 2001, 135, 239–247. [Google Scholar]
- Pezzino, G.; Miller, C.; Flahart, R.; Potsic, S.R. A multi-state outbreak of Salmonella serotypes Infantis and Anatum - Kansas and Missouri, 1997. Kansas Medicine: J. Kansas Med. Soc. 1998, 98, 10–12. [Google Scholar]
- Gill, C.J.; Keene, W.E.; Mohle-Boetani, J.C.; Farrar, J.A.; Waller, P.L.; Hahn, C.G.; Cieslak, P.R. Alfalfa seed decontamination in a Salmonella outbreak. Emerg. Infect. Dis. 2003, 9, 474–479. [Google Scholar] [CrossRef]
- Proctor, M.E.; Hamacher, M.; Tortorello, M.L.; Archer, J.R.; Davis, J.P. Multistate outbreak of Salmonella serovar Muenchen infections associated with alfalfa sprouts grown from seeds pretreated with calcium hypochlorite. J. Clin. Microbiol. 2001, 39, 3461–3465. [Google Scholar] [CrossRef]
- Brooks, J.T.; Rowe, S.Y.; Shillam, P.; Heltzel, D.M.; Hunter, S.B.; Slutsker, L.; Hoekstra, R.M.; Luby, S.P. Salmonella Typhimurium infections transmitted by chlorine-pretreated clover sprout seeds. Am. J. Epidemiol. 2001, 154, 1020–1028. [Google Scholar] [CrossRef]
- Winthrop, K.L.; Palumbo, M.S.; Farrar, J.A.; Mohle-Boetani, J.C.; Abbott, S.; Beatty, M.E.; Inami, G.; Werner, S.B. Alfalfa sprouts and Salmonella Kottbus infection: A multistate outbreak following inadequate seed disinfection with heat and chlorine. J. Food Prot. 2003, 66, 13–17. [Google Scholar]
- CDC. Outbreak of Salmonella serotype Saintpaul infections associated with eating alfalfa sprouts - United States, 2009. MMWR 2009, 58, 500–503. [Google Scholar]
- CDC. Salmonella newport on alfalfa sprouts. Available online: http://www.cdc.gov/salmonella/newport/index.html (Accessed on 5 June 2012).
- CDC. Salmonella linked to alfalfa sprouts. Available online: http://www.cdc.gov/salmonella/i4512i-/021011/index.html (Accessed on June 5, 2012).
- CDC. Salmonella Enteritidis infections on alfalfa sprouts. Available online: http://www.cdc.gov/salmonella/sprouts-enteritidis0611/index.html (Accessed on 5 June 2012).
- Lehmacher, A.; Bockemuhl, J.; Aleksic, S. Nationwide outbreak of human salmonellosis in germany due to contaminated paprika and paprika-powdered potato chips. Epidemiol. Infect. 1995, 115, 501–511. [Google Scholar] [CrossRef]
- CDC. Multistate outbreak of Salmonella serotype Agona infections linked to toasted oats cereal--United States, April-May, 1998. MMWR 1998, 47, 462–464. [Google Scholar]
- Hiramatsu, R.; Matsumoto, M.; Sakae, K.; Miyazaki, Y. Ability of shiga toxin-producing Escherichia coli and Salmonella spp. To survive in a desiccation model system and in dry foods. Appl. Environ. Microbiol. 2005, 71, 6657–6663. [Google Scholar] [CrossRef]
- Tsuji, H.; Hamada, K. Outbreak of salmonellosis caused by ingestion of cuttlefish chips contaminated by both Salmonella Chester and Salmonella Oranienburg. Jpn. J.Infect. Dis. 1999, 52, 138–139. [Google Scholar]
- Gastrin, B.; Kampe, A.; Nystrom, K.G.; Oden-Johanson, B.; Wessel, G.; Zetterberg, B. Salmonella durham epidemic caused by contaminated cocoa. Lakartidningen 1972, 69, 5335–5338. [Google Scholar]
- Craven, P.C.; Mackel, D.C.; Baine, W.B.; Barker, W.H.; Gangarosa, E.J. International outbreak of Salmonella Eastbourne infection traced to contaminated chocolate. Lancet 1975, 1, 788–792. [Google Scholar]
- D'Aoust, J.Y.; Aris, B.J.; Thisdele, P.; Durante, A.; Brisson, N.; Dragon, D.; Lachapelle, G.; Johnston, M.; Laidley, R. Salmonella eastbourne outbreak associated with chocolate. Can. Inst. Food Sci. Technol. J. 1975, 8, 181–184. [Google Scholar]
- Gill, O.N.; Sockett, P.N.; Bartlett, C.L.; Vaile, M.S.; Rowe, B.; Gilbert, R.J.; Dulake, C.; Murrell, H.C.; Salmaso, S. Outbreak of Salmonella napoli infection caused by contaminated chocolate bars. Lancet 1983, 1, 574–577. [Google Scholar]
- Hockin, J.C.; D'Aoust. An international outbreak of Salmonella nima from imported chocolate. J. Food Prot. 1989, 52, 51–54. [Google Scholar]
- Kapperud, G.; Gustavsen, S.; Hellesnes, I.; Hansen, A.H.; Lassen, J.; Hirn, J.; Jahkola, M.; Montenegro, M.A.; Helmuth, R. Outbreak of Salmonella typhimurium infection traced to contaminated chocolate and caused by a strain lacking the 60-megadalton virulence plasmid. J. Clin. Microbiol. 1990, 28, 2597–2601. [Google Scholar]
- Werber, D.; Dreesman, J.; Feil, F.; van Treeck, U.; Fell, G.; Ethelberg, S.; Hauri, A.M.; Roggentin, P.; Prager, R.; Fisher, I.S.T.; Behnke, S.C.; Bartelt, E.; Weise, E.; Ellis, A.; Siitonen, A.; Andersson, Y.; Tschape, H.; Kramer, M.H.; Ammon, A. International outbreak of Salmonella Oranienburg due to German chocolate. BMC Infect. Dis. 2005, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Micallef, S.A.; Rosenberg Goldstein, R.E.; George, A.; Kleinfelter, L.; Boyer, M.S.; McLaughlin, C.R.; Estrin, A.; Ewing, L.; Jean-Gilles Beaubrun, J.; Hanes, D.E.; Kothary, M.H.; Tall, B.D.; Razeq, J.H.; Joseph, S.W.; Sapkota, A.R. Occurrence and antibiotic resistance of multiple Salmonella serotypes recovered from water, sediment and soil on mid-atlantic tomato farms. Environ. Res. 2012, 114, 31–39. [Google Scholar] [CrossRef]
- Gaertner, J.P.; Garres, T.; Becker, J.C.; Jimenez, M.L.; Forstner, M.R.J.; Hahn, D. Temporal analyses of salmonellae in a headwater spring ecosystem reveals the effects of precipitation and runoff events. J. Water Health 2009, 7, 115–121. [Google Scholar] [CrossRef]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, diversity, and seasonality of waterborne salmonellae in a rural watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar]
- Polo, F.; Figueras, M.J.; Inza, I.; Sala, J.; Fleisher, J.M.; Guarro, J. Relationship between presence of Salmonella and indicators of faecal pollution in aquatic habitats. FEMS Microbiol. Lett. 1998, 160, 253–256. [Google Scholar] [CrossRef]
- Polo, F.; Figueras, M.J.; Inza, I.; Sala, J.; Fleisher, J.M.; Guarro, J. Prevalence of Salmonella serotypes in environmental waters and their relationships with indicator organisms. Antonie Van Leeuwenhoek 1999, 75, 285–292. [Google Scholar] [CrossRef]
- Santo Domingo, J.W.; Harmon, S.; Bennett, J. Survival of Salmonella species in river water. Curr. Microbiol. 2000, 40, 409–417. [Google Scholar]
- Gaertner, J.P.; Mendoza, J.A.; Forstner, M.R.J.; Hahn, D. Recovery of Salmonella from biofilms in a headwater spring ecosystem. J. Water Health 2011, 9, 458–466. [Google Scholar]
- Hintz, L.D.; Boyer, R.R.; Ponder, M.A.; Williams, R.C.; Rideout, S.L. Recovery of Salmonella enterica newport introduced through irrigation water from tomato (Lycopersicum esculentum) fruit, roots, stems, and leaves. HortScience 2010, 45, 675–678. [Google Scholar]
- Guo, X.; van Iersel, M.W.; Chen, J.; Brackett, R.E.; Beuchat, L.R. Evidence of association of salmonellae with tomato plants grown hydroponically in inoculated nutrient solution. Appl. Environ. Microbiol. 2002, 68, 3639–3643. [Google Scholar] [CrossRef]
- Miles, J.M.; Sumner, S.S.; Boyer, R.R.; Williams, R.C.; Latimer, J.G.; McKinney, J.M. Internalization of Salmonella enterica serovar Montevideo into greenhouse tomato plants through contaminated irrigation water or seed stock. J. Food Prot. 2009, 72, 849–852. [Google Scholar]
- Gu, G.; Hu, J.; Cevallos-Cevallos, J.M.; Richardson, S.M.; Bartz, J.A.; van Bruggen, A.H.C. Internal colonization of Salmonella enterica serovar Typhimurium in tomato plants. PLoS One 2011, 6, e27340. [Google Scholar]
- Zhuang, R.Y.; Beuchat, L.R.; Angulo, F.J. Fate of Salmonella montevideo on and in raw tomatoes as affected by temperature and treatment with chlorine. Appl. Environ.Microbiol. 1995, 61, 2127–2131. [Google Scholar]
- Backer, H.D.; Mohle-Boetani, J.C.; Werner, S.B.; Abbott, S.L.; Farrar, J.; Vugia, D.J. High incidence of extra-intestinal infections in a Salmonella Havana outbreak associated with alfalfa sprouts. Pub. Health Rep. 2000, 115, 339–339. [Google Scholar] [CrossRef]
- National Advisory Committee on Microbiological Criteria for Food. Microbiological safety evaluations and recommendations on sprouted seeds. Int. J. Food Microbiol. 1999, 52, 123–153. [Google Scholar] [CrossRef]
- Weissinger, W.R.; Beuchat, L.R. Comparison of aqueous chemical treatments to eliminate Salmonella on alfalfa seeds. J. Food Prot. 2000, 63, 1475–1482. [Google Scholar]
- Beuchat, L.R.; Ryu, J.H. Produce handling and processing practices. Emerg. Infect. Dis. 1997, 3, 459–465. [Google Scholar] [CrossRef]
- Jaquette, C.B.; Beuchat, L.R.; Mahon, B.E. Efficacy of chlorine and heat treatment in killing Salmonella stanley inoculated onto alfalfa seeds and growth and survival of the pathogen during sprouting and storage. Appl. Environ. Microbiol. 1996, 62, 2212–2215. [Google Scholar]
- Andrews, W.H.; Mislivec, P.B.; Wilson, C.R.; Bruce, V.R.; Poelma, P.L.; Gibson, R.; Trucksess, M.W.; Young, K. Microbial hazards associated with bean sprouting. J. Assoc. Off. Anal. Chem. 1982, 65, 241–248. [Google Scholar]
- Dong, Y.; Iniguez, A.L.; Ahmer, B.M.M.; Triplett, E.W. Kinetics and strain specificity of rhizosphere and endophytic colonization by enteric bacteria on seedlings of Medicago sativa and Medicago truncatula. Appl. Environ. Microbiol. 2003, 69, 1783–1790. [Google Scholar]
- Barak, J.D.; Gorski, L.; Naraghi-Arani, P.; Charkowski, A.O. Salmonella enterica virulence genes are required for bacterial attachment to plant tissue. Appl. Environ. Microbiol. 2005, 71, 5685–5691. [Google Scholar]
- Podolak, R.; Enache, E.; Stone, W.; Black, D.G.; Elliott, P.H. Sources and risk factors for contamination, survival, persistence, and heat resistance of Salmonella in low-moisture foods. J. Food Prot. 2010, 73, 1919–1936. [Google Scholar]
- Chia, T.W.R.; Goulter, R.M.; McMeekin, T.; Dykes, G.A.; Fegan, N. Attachment of different Salmonella serovars to materials commonly used in a poultry processing plant. Food Microbiol. 2009, 26, 853–859. [Google Scholar] [CrossRef]
- Morita, T.; Kitazawa, H.; Iida, T.; Kamata, S. Prevention of Salmonella cross-contamination in an oilmeal manufacturing plant. J. Appl. Microbiol. 2006, 101, 464–473. [Google Scholar] [CrossRef]
- Marin, C.; Hernandiz, A.; Lainez, M. Biofilm development capacity of Salmonella strains isolated in poultry risk factors and their resistance against disinfectants. Poult. Sci. 2009, 88, 424–431. [Google Scholar] [CrossRef]
- Nesse, L.L.; Nordby, K.; Heir, E.; Bergsjoe, B.; Vardund, T.; Nygaard, H.; Holstad, G. Molecular analyses of Salmonella enterica isolates from fish feed factories and fish feed ingredients. Appl. Environ. Microbiol. 2003, 69, 1075–1081. [Google Scholar] [CrossRef]
- Vestby, L.K.; Moretro, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonella are correlated with persistence in fish meal- and feed factories. BMC Vet. Res. 2009, 5, 20. [Google Scholar] [CrossRef]
- Refsum, T.r.; Handeland, K.; Baggesen, D.L.; Holstad, G.; Kapperud, G. Salmonellae in avian wildlife in norway from 1969 to 2000. Appl. Environ. Microbiol. 2002, 68, 5595–5599. [Google Scholar] [CrossRef]
- Solomon, E.B.; Niemira, B.A.; Sapers, G.M.; Annous, B.A. Biofilm formation, cellulose production, and curli biosynthesis by Salmonella originating from produce, animal, and clinical sources. J. Food Prot. 2005, 68, 906–912. [Google Scholar]
- Stepanovic, S.; Cirkovic, I.; Mijac, V.; Svabic-Vlahovic, M. Influence of the incubation temperature, atmosphere and dynamic conditions on biofilm formation by Salmonella spp. Food Microbiol. 2003, 20, 339–343. [Google Scholar] [CrossRef]
- Patel, J.; Sharma, M. Differences in attachment of Salmonella enterica serovars to cabbage and lettuce leaves. Int. J. Food Microbiol. 2010, 139, 41–47. [Google Scholar] [CrossRef]
- Lund, B.M.; Eklund, T. Control of pH and use of organic acids. In Microbiological Safety and Quality of Food; Aspen Publishers: Gaithersburg, MD, 2000; pp. 175–199. [Google Scholar]
- Tamminga, S.K.; Beumer, R.R.; Kampelmacher, E.H.; van Leusden, F.M. Survival of Salmonella eastbourne and Salmonella typhimurium in chocolate. J. Hyg. 1976, 76, 41–47. [Google Scholar] [CrossRef]
- Goepfert, J.M.; Biggie, R.A. Heat resistance of Salmonella typhimurium and Salmonella senftenberg 775W in milk chocolate. Appl. Microbiol. 1968, 16, 1939–1940. [Google Scholar]
- Tamminga, S.K.; Beumer, R.R.; Kampelmacher, E.H.; van Leusden, F.M. Survival of Salmonella eastbourne and Salmonella typhimurium in milk chocolate prepared with artificially contaminated milk powder. J. Hyg. 1977, 79, 333–337. [Google Scholar] [CrossRef]
- Blaser, M.J.; Newman, L.S. A review of human salmonellosis: I. Infective dose. Rev. Infect. Dis. 1982, 4, 1096–1106. [Google Scholar] [CrossRef]
- D'Aoust, J.Y. Salmonella and the chocolate industry. A review. J. Food Prot. 1977, 40, 718–727. [Google Scholar]
- Chan, K.; Baker, S.; Kim, C.C.; Detweiler, C.S.; Dougan, G.; Falkow, S. Genomic comparison of Salmonella enterica serovars and Salmonella bongori by use of an S. enterica serovar Typhimurium DNA microarray. J. Bacteriol. 2003, 185, 553–563. [Google Scholar]
- Didelot, X.; Bowden, R.; Street, T.; Golubchik, T.; Spencer, C.; McVean, G.; Sangal, V.; Anjum, M.F.; Achtman, M.; Falush, D.; Donnelly, P. Recombination and population structure in Salmonella enterica. PLoS Genet. 2011, 7, e1002191. [Google Scholar]
- den Bakker, H.C.; Switt, A.I.M.; Govoni, G.; Cummings, C.A.; Ranieri, M.L.; Degoricija, L.; Hoelzer, K.; Rodriguez-Rivera, L.D.; Brown, S.; Bolchacova, E.; et al. Genome sequencing reveals diversification of virulence factor content and possible host adaptation in distinct subpopulations of Salmonella enterica. BMC Genomics 2011, 12, 425. [Google Scholar] [CrossRef]
- Sangal, V.; Harbottle, H.; Mazzoni, C.J.; Helmuth, R.; Guerra, B.; Didelot, X.; Paglietti, B.; Rabsch, W.; Brisse, S.; Weill, F.X.; Roumagnac, P.; Achtman, M. Evolution and population structure of Salmonella enterica serovar Newport. J. Bacteriol. 2010, 192, 6465–6476. [Google Scholar]
- Holt, K.E.; Parkhill, J.; Mazzoni, C.J.; Roumagnac, P.; Weill, F.-X.; Goodhead, I.; Rance, R.; Baker, S.; Maskell, D.J.; Wain, J.; Dolecek, C.; Achtman, M.; Dougan, G. High-throughput sequencing provides insights into genome variation and evolution in Salmonella Typhi. Nat. Genet. 2008, 40, 987–993. [Google Scholar]
- McClelland, M.; Sanderson, K.E.; Clifton, S.W.; Latreille, P.; Porwollik, S.; Sabo, A.; Meyer, R.; Bieri, T.; Ozersky, P.; McLellan, M.; et al. Comparison of genome degradation in Paratyphi A and Typhi, human-restricted serovars of Salmonella enterica that cause typhoid. Nat. Genet. 2004, 36, 1268–1274. [Google Scholar]
- Kingsley, R.A.; Santos, R.L.; Keestra, A.M.; Adams, L.G.; B√§umler, A.J. Salmonella enterica serotype Typhimurium ShdA is an outer membrane fibronectin-binding protein that is expressed in the intestine. Mol. Microbiol. 2002, 43, 895–905. [Google Scholar] [CrossRef]
- Kingsley, R.A.; van Amsterdam, K.; Kramer, N.; Baumler, A.J. The shdA gene is restricted to serotypes of Salmonella enterica subspecies I and contributes to efficient and prolonged fecal shedding. Infect. Immun. 2000, 68, 2720–2727. [Google Scholar]
- Thomson, N.R.; Clayton, D.J.; Windhorst, D.; Vernikos, G.; Davidson, S.; Churcher, C.; Quail, M.A.; Stevens, M.; Jones, M.A.; Watson, M.; et al. Comparative genome analysis of Salmonella Enteritidis PT4 and Salmonella Gallinarum 287/91 provides insights into evolutionary and host adaptation pathways. Genome Res. 2008, 18, 1624–1637. [Google Scholar] [CrossRef]
- Betancor, L.; Yim, L.; Fookes, M.; Martinez, A.; Thomson, N.R.; Ivens, A.; Peters, S.; Bryant, C.; Algorta, G.; Kariuki, S.; Schelotto, F.; Maskell, D.; Dougan, G.; Chabalgoity, J.A. Genomic and phenotypic variation in epidemic-spanning Salmonella enterica serovar Enteritidis isolates. BMC Microbiol. 2009, 9, 237. [Google Scholar] [CrossRef]
- Fricke, W.F.; Mammel, M.K.; McDermott, P.F.; Tartera, C.; White, D.G.; LeClerc, J.E.; Ravel, J.; Cebula, T.A. Comparative genomics of 28 Salmonella enterica isolates: Evidence for crispr-mediated adaptive sublineage evolution. J. Bacteriol. 2011, 193, 3556–3568. [Google Scholar] [CrossRef]
- Hoffmann, M.; Zhao, S.; Luo, Y.; Li, C.; Folster, J.P.; Whichard, J.; Allard, M.W.; Brown, E.W.; McDermott, P.F. Genome sequences of five Salmonella enterica serovar Heidelberg isolates associated with a 2011 multistate outbreak in the united states. J. Bacteriol. 2012, 194, 3274–3275. [Google Scholar] [CrossRef]
- Okoro, C.K.; Kingsley, R.A.; Quail, M.A.; Kankwatira, A.M.; Feasey, N.A.; Parkhill, J.; Dougan, G.; Gordon, M.A. High-resolution single nucleotide polymorphism analysis distinguishes recrudescence and reinfection in recurrent invasive nontyphoidal Salmonella Typhimurium disease. Clin. Infect. Dis. 2012, 54, 955–963. [Google Scholar]
- Richardson, E.J.; Limaye, B.; Inamdar, H.; Datta, A.; Manjari, K.S.; Pullinger, G.D.; Thomson, N.R.; Joshi, R.R.; Watson, M.; Stevens, M.P. Genome sequences of Salmonella enterica serovar Typhimurium, Choleraesuis, Dublin, and Gallinarum strains of well- defined virulence in food-producing animals. J. Bacteriol. 2011, 193, 3162–3163. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, W.Q.; Sanderson, K.E.; Liu, S.L. Comparison of salmonella genomes. In Salmonella: From genome to function; Caister Academic Press: Norfolk, UK, 2011; pp. 49–67. [Google Scholar]
- Leekitcharoenphon, P.; Lukjancenko, O.; Friis, C.; Aarestrup, F.M.; Ussery, D.W. Genomic variation in Salmonella enterica core genes for epidemiological typing. BMC Genomics 2012, 13. [Google Scholar] [CrossRef]
- Didelot, X.; Achtman, M.; Parkhill, J.; Thomson, N.R.; Falush, D. A bimodal pattern of relatedness between the salmonella paratyphi a and typhi genomes: Convergence or divergence by homologous recombination? Genome Res. 2007, 17, 61–68. [Google Scholar]
- Holt, K.E.; Perkins, T.T.; Dougan, G.; Kingsley, R.A. Genomics and pathogenesis of Salmonella enterica serovars Typhi and Paratyphi A. In Salmonella: From genome to function; Caister Academic Press: Norfolk, UK, 2011; pp. 107–121. [Google Scholar]
- Romling, U.; Bokranz, W.; Rabsch, W.; Zogaj, X.; Nimtz, M.; Tschape, H. Occurrence and regulation of the multicellular morphotype in Salmonella serovars important in human disease. Int.J. Med. Microbiol. 2003, 293, 273–285. [Google Scholar] [CrossRef]
- White, A.P.; Surette, M.G. Comparative genetics of the rdar morphotype in Salmonella. J. Bacteriol. 2006, 188, 8395–8406. [Google Scholar] [CrossRef]
- Mahajan, R.K.; Khan, S.A.; Chandel, D.S.; Kumar, N.; Hans, C.; Chaudhry, R. Fatal case of Salmonella enterica subsp. arizonae gastroenteritis in an infant with microcephaly. J. Clin. Microbiol. 2003, 41, 5830–5832. [Google Scholar]
- Meric, G.; Kemsley, E.K.; Falush, D.; Saggers, E.J.; Lucchini, S. Phylogenetic distribution of traits associated with plant colonization in Escherichia coli. Environ. Microbiol. 2012. [CrossRef]
- Sakellaris, H.; Hannink, N.K.; Rajakumar, K.; Bulach, D.; Hunt, M.; Sasakawa, C.; Adler, B. Curli loci of Shigella spp. Infect. Immun. 2000, 68, 3780–3783. [Google Scholar] [CrossRef]
- Bokranz, W.; Wang, X.; Tschape, H.; Romling, U. Expression of cellulose and curli fimbriae by Escherichia coli isolated from the gastrointestinal tract. J. Med. Microbiol. 2005, 54, 1171–1182. [Google Scholar] [CrossRef]
- Crawford, R.W.; Rosales-Reyes, R.; Ramirez-Aguilar, M.d.l.L.; Chapa-Azuela, O.; Alpuche-Aranda, C.; Gunn, J.S. Gallstones play a significant role in Salmonella spp. gallbladder colonization and carriage. Proc. Natl. Acad. Sci. USA 2010, 107, 4353–4358. [Google Scholar]
- Lawley, T.D.; Bouley, D.M.; Hoy, Y.E.; Gerke, C.; Relman, D.A.; Monack, D.M. Host transmission of Salmonella enterica serovar Typhimurium is controlled by virulence factors and indigenous intestinal microbiota. Infect. Immun. 2007, 76, 403–416. [Google Scholar]
- Gordon, M.A. Invasive nontyphoidal Salmonella disease. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef]
Supplementary
Graphical Representation of Waldner et al. Manuscript

© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Waldner, L.L.; MacKenzie, K.D.; Köster, W.; White, A.P. From Exit to Entry: Long-term Survival and Transmission of Salmonella. Pathogens 2012, 1, 128-155. https://doi.org/10.3390/pathogens1020128
Waldner LL, MacKenzie KD, Köster W, White AP. From Exit to Entry: Long-term Survival and Transmission of Salmonella. Pathogens. 2012; 1(2):128-155. https://doi.org/10.3390/pathogens1020128
Chicago/Turabian StyleWaldner, Landon L., Keith D. MacKenzie, Wolfgang Köster, and Aaron P. White. 2012. "From Exit to Entry: Long-term Survival and Transmission of Salmonella" Pathogens 1, no. 2: 128-155. https://doi.org/10.3390/pathogens1020128
APA StyleWaldner, L. L., MacKenzie, K. D., Köster, W., & White, A. P. (2012). From Exit to Entry: Long-term Survival and Transmission of Salmonella. Pathogens, 1(2), 128-155. https://doi.org/10.3390/pathogens1020128