Rapid Increase in the IS26-Mediated cfr Gene in E. coli Isolates with IncP and IncX4 Plasmids and Co-Existing cfr and mcr-1 Genes in a Swine Farm

 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Prevalence of the cfr Gene and Detection of Other Resistance Genes
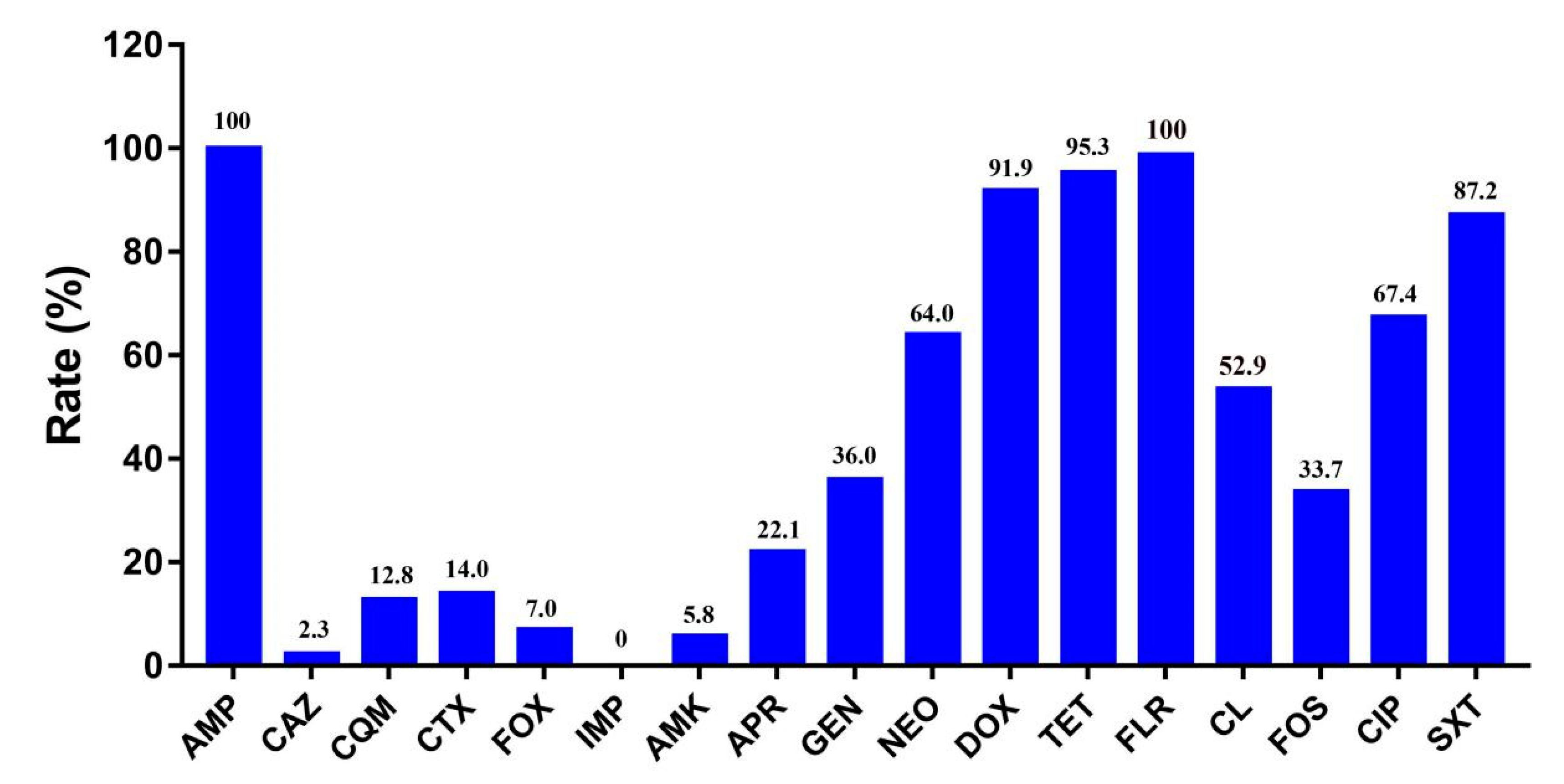
2.2. Antibiotic Susceptibility Testing
2.3. Pulsed-Field Gel Electrophoresis (PFGE) Patterns, Location, and Genetic Context of the cfr Gene
2.4. Co-Transfer of the mcr-1 and cfr Genes
2.5. Plasmid Analysis of pHNEP124
2.6. Plasmid Analysis of pHNEP129
3. Discussion
4. Materials and Methods
4.1. Detection of Multi-Resistant cfr Gene and Other Resistance Genes
4.2. Antibiotic Susceptibility Testing
4.3. Pulsed-Field Gel Electrophoresis (PFGE) and Flanking Regions of the cfr Gene
4.4. S1-PFGE and Southern Blotting Hybridization
4.5. Conjugation Experiment and Transformation Assay
4.6. PBRT and Plasmid Analysis
4.7. Nucleotide Sequence Accession Number
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The Cfr rRNA methyltransferase confers resistance to phenicols, lincosamides, oxazolidinones, pleuromutilins, and streptogramin A antibiotics. Antimiocrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.K.; Mankin, A.S. Transcriptional and translational control of the mlr operon, which confers resistance to seven classes of protein synthesis inhibitors. Antimicrob. Agents Chemother. 2008, 52, 1703–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; Hansen, L.H.; Vester, B.A. New mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef]
- Chen, Y.; Lei, C.; Zuo, L.; Kong, L.; Kang, Z.; Zeng, J.; Zhang, X.; Wang, H. A novel cfr-carrying Tn7 transposon derivative characterized in Morganella morganii of swine origin in China. J. Antimicrob. Chemother. 2018, 74, 603–606. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial resistance in Escherichia coli. Microbiol. Spectrum 2018, 6, ARBA-0026-2017. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhu, Y.; Hua, X.; Chen, F.; Wang, C.; Zhang, Y.; Liu, S.; Zhang, W. F14:A-:B- and IncX4 Inc group cfr-positive plasmids circulating in Escherichia coli of animal origin in Northeast China. Vet. Microbiol. 2018, 217, 53–57. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends. Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef]
- Baron, S.; Hadjadj, L.; Rolain, J.M.; Olaitan, A.O. Molecular mechanisms of polymyxin resistance: Knowns and unknowns. Int. J. Antimicrob. Agents 2016, 48, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Q.; Wang, J.; Li, W.; Zhao, L.-Q.; Lu, Y.; Liu, J.-H.; Zeng, Z.-L. Distribution of cfr in Staphylococcus spp. and Escherichia coli Strains from Pig Farms in China and Characterization of a Novel cfr-Carrying F43:A-:B- Plasmid. Front. Microbiol. 2017, 8, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; He, T.; Schwarz, S.; Zhou, D.; Shen, Z.; Wu, C.; Wang, Y.; Ma, L.; Zhang, Q.; Shen, J. Detection of the staphylococcal multiresistance gene cfr in Escherichia coli of domestic-animal origin. J. Antimicrob. Chemother. 2012, 67, 1094–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, H.; Sun, J.; Ma, J.; Li, L.; Fang, L.-X.; Zhang, Q.; Liu, Y.-H.; Liao, X.-P. Identification of the multi-resistance gene cfr in Escherichia coli isolates of animal origin. PLoS ONE 2014, 9, e102378. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Yi, L.X.; Yu, L.F.; Wang, J.; Liu, Y.; Chen, X.; Lv, L.; Yang, J.; Liu, J.-H. Fitness Advantage of mcr-1-Bearing IncI2 and IncX4 Plasmids in Vitro. Front. Microbiol. 2018, 9, 331. [Google Scholar] [CrossRef]
- Walsh, T.R.; Wu, Y. China bans colistin as a feed additive for animals. Lancet Infect. Dis. 2016, 16, 1102–1103. [Google Scholar] [CrossRef]
- Li, W.; Hou, M.; Liu, C.; Xiong, W.; Zeng, Z. Dramatic decrease of the colistin resistance in E. coli from a typical farm following the restriction of use of colistin in China. Int. J. Antimicrob. Agents 2019, 53, 707–708. [Google Scholar] [CrossRef]
- Zhang, R.; Sun, B.; Wang, Y.; Lei, L.; Schwarz, S.; Wu, C. Characterization of a cfr-carrying plasmid from porcine Escherichia coli that closely resembles plasmid pEA3 from the plant pathogen Erwinia amylovora. Antimicrob. Agents Chemother. 2016, 60, 658–661. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-J.; Xu, X.-R.; Schwarz, S.; Wang, X.-M.; Dai, L.; Zheng, H.-J.; Liu, S. Characterization of the IncA/C plasmid pSCEC2 from Escherichia coli of swine origin that harbours the multiresistance gene cfr. J. Antimicrob. Chemother. 2014, 69, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Deng, H.; Li, L.; Chen, M.-Y.; Fang, L.X.; Yang, Q.-E.; Liu, Y.-H.; Liao, X.-P. Complete nucleotide sequence of cfr-carrying IncX4 plasmid pSD11 from Escherichia coli. Antimicrob. Agents Chemother. 2015, 59, 738–741. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Feng, Y.; Lü, X.; McNally, A.; Zong, Z. IncP Plasmid Carrying Colistin Resistance Gene mcr-1 in Klebsiella pneumoniae from Hospital Sewage. Antimicrob. Agents Chemother. 2017, 61, e02229-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Feng, Y.; Lü, X.; McNally, A.; Zong, Z. New Variant of mcr-3 in an Extensively Drug-Resistant Escherichia coli Clinical Isolate Carrying mcr-1 and blaNDM-5. Antimicrob. Agents Chemother. 2017, 61, e01757-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Cao, Y.; Yu, P.; Huang, R.; Wang, J.; Wen, Q.; Zhi, C.; Zhang, Q.; Liu, J.-H. Detection of mcr-1 Gene among Escherichia coli Isolates from Farmed Fish and Characterization of mcr-1-Bearing IncP Plasmids. Antimicrob. Agents Chemother. 2018, 62, e02378-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Schwarz, S.; Wang, S.; Chen, L.; Wu, C.; Shen, J. Investigation of a multiresistance gene cfr that fails to mediate resistance to phenicols and oxazolidinones in Enterococcus faecalis. J. Antimicrob. Chemother. 2014, 69, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Brenciani, A.; Morroni, G.; Vincenzi, C.; Manso, E.; Mingoia, M.; Giovanetti, E.; Varaldo, P.E. Detection in Italy of two clinical Enterococcus faecium isolates carrying both the oxazolidinone and phenicol resistance gene optrA and a silent multiresistance gene cfr. J. Antimicrob. Chemother. 2016, 71, 1118–1189. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Kim, Y.H.; Kim, S.E.; Lee, J.H.; Park, C.S.; Kim, H.Y. Identification and distribution of bacillus species in doenjang by whole-cell protein patterns and 16S rRNA gene sequence analysis. J. Microbiol. Biotechnol. 2010, 20, 1210–1214. [Google Scholar] [CrossRef] [Green Version]
- Kehrenberg, C.; Schwarz, S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standard fourth Edition and Supplement; Documents VET01-A4 and VET01-S2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Barton, B.M.; Harding, G.P.; Zuccarelli, A.J. A general method for detecting and sizing large plasmids. Anal. Biochem. 1995, 226, 235–240. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collected Date | Suckling Piglet (N) | Nursery Pig (N) | Fattening Pig (N) | Sow (N) | Boar (N) | Environment (N) | Total | History of Drug Use |
|---|---|---|---|---|---|---|---|---|
| Jun. 2014 | 33 | 35 (3) | 64 | 52 | 0 | 0 | 184 (3, 1.6%) | gentamycin, amoxicillin |
| Jul. 2015 | 28 | 39 (3) | 46 (10) | 38 | 0 | 0 | 151 (13, 8.6%) | florfenicol |
| Jul. 2016 | 8 | 32 (9) | 45 (18) | 51 | 4 (1) | 1 | 141 (28, 19.9%) | florfenicol, enrofloxacin, mequindox, kanamycin |
| Apr. 2017 | 14 | 23 (14) | 51 (24) | 36 (1) | 9 | 8 (2) | 141 (41, 29.1%) | gentamycin, amoxicillin, kanamycin |
| Total | 83 (0) | 129 (29, 22.5%) | 206 (52, 25.2%) | 177 (1, 0.6%) | 13 (1, 7.7%) | 9 (2, 22.2%) | 617 (85, 13.7%) | - |
| Strains | MIC Values | Replicon Type | Resistance Phenotype a | Resistance Genes | |
|---|---|---|---|---|---|
| Florfenicol | Colistin | ||||
| GDE5P101J | 2 | 4 | IncI2 | CL | cfr mcr-1 |
| GDE5P123J | 4 | 4 | IncI2 | AMP CL | cfr mcr-1 |
| GDE5P132J | 2 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE5P137J | 4 | 8 | IncX4 | AMP APR CL | cfr mcr-1 |
| GDE5P138J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE5P141J | 4 | 8 | IncHI2 | CL | cfr mcr-1 |
| GDE5P142J | 8 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE5P145J | 2 | 8 | IncX4 | CL | cfr mcr-1 |
| GDE5P148J | 8 | 4 | - | AMP CL | cfr mcr-1 |
| GDE5P165J | 8 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P75J | 8 | 4 | IncHI2 | AMP CTX CL FOS | cfr mcr-1 |
| GDE6P85J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P103J | 4 | 4 | - | AMP CL | cfr mcr-1 |
| GDE6P110J | 8 | 2 | IncX4 | CL | cfr mcr-1 |
| GDE6P115J | 2 | 4 | - | AMP CL | cfr mcr-1 |
| GDE6P124J | 8 | 4 | IncP b | AMP CL | cfr mcr-1 |
| GDE6P128J | 8 | 4 | IncP | AMP CL | cfr mcr-1 |
| GDE6P129J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P130J | 4 | 4 | IncX4 | AMP CL | cfr mcr-1 |
| GDE6P133J | >128 | 4 | IncX4 | AMP FLR CL | cfr mcr-1 floR |
| GDE6P143J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P151J | >128 | 4 | IncX4 | AMP GEN FLR CL | cfr mcr-1 floR |
| GDE6P164J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P165J | 4 | 4 | IncX4 | CL | cfr mcr-1 |
| GDE6P168J | 4 | 4 | IncX4 | AMP CL | cfr mcr-1 |
| GDE6P169J | 4 | 4 | IncX4 | AMP CL | cfr mcr-1 |
| GDE7P75J | 16 | 4 | IncX4 | CL SXT | cfr mcr-1 |
| GDE7P100J | 8 | 4 | IncHI2 IncX4 | AMP APR DOX CL SXT | cfr mcr-1 |
| GDE7P128J | 8 | 8 | IncI2 | AMP CL SXT | cfr mcr-1 |
| GDE7P166J | 16 | 2 | IncX4 | AMP DOX CL | cfr mcr-1 |
| coli C600 | 4 | 0.125 | - c | - | - |
| ATCC 25922 | 2 | 0.5 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Liu, J.; Chen, L.; Liu, X.; Xiong, W.; Liu, J.-H.; Zeng, Z. Rapid Increase in the IS26-Mediated cfr Gene in E. coli Isolates with IncP and IncX4 Plasmids and Co-Existing cfr and mcr-1 Genes in a Swine Farm. Pathogens 2021, 10, 33. https://doi.org/10.3390/pathogens10010033
Ma Z, Liu J, Chen L, Liu X, Xiong W, Liu J-H, Zeng Z. Rapid Increase in the IS26-Mediated cfr Gene in E. coli Isolates with IncP and IncX4 Plasmids and Co-Existing cfr and mcr-1 Genes in a Swine Farm. Pathogens. 2021; 10(1):33. https://doi.org/10.3390/pathogens10010033
Chicago/Turabian StyleMa, Zhenbao, Jiao Liu, Lin Chen, Xiaoqin Liu, Wenguang Xiong, Jian-Hua Liu, and Zhenling Zeng. 2021. "Rapid Increase in the IS26-Mediated cfr Gene in E. coli Isolates with IncP and IncX4 Plasmids and Co-Existing cfr and mcr-1 Genes in a Swine Farm" Pathogens 10, no. 1: 33. https://doi.org/10.3390/pathogens10010033