
The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission

 ,
,
Abstract
:1. Introduction
2. Results
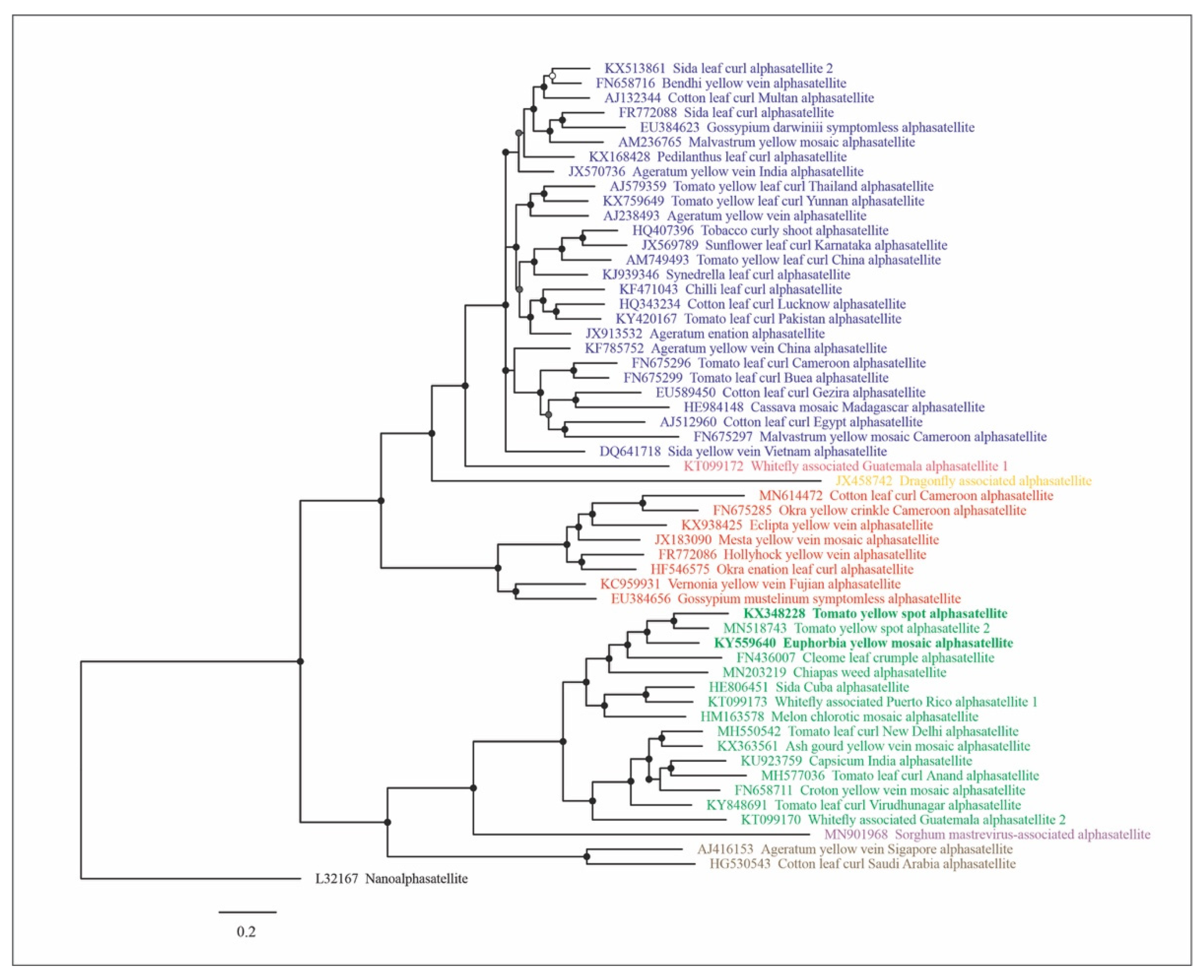
2.1. Phylogeny of Geminivirus-Associated Alphasatellites
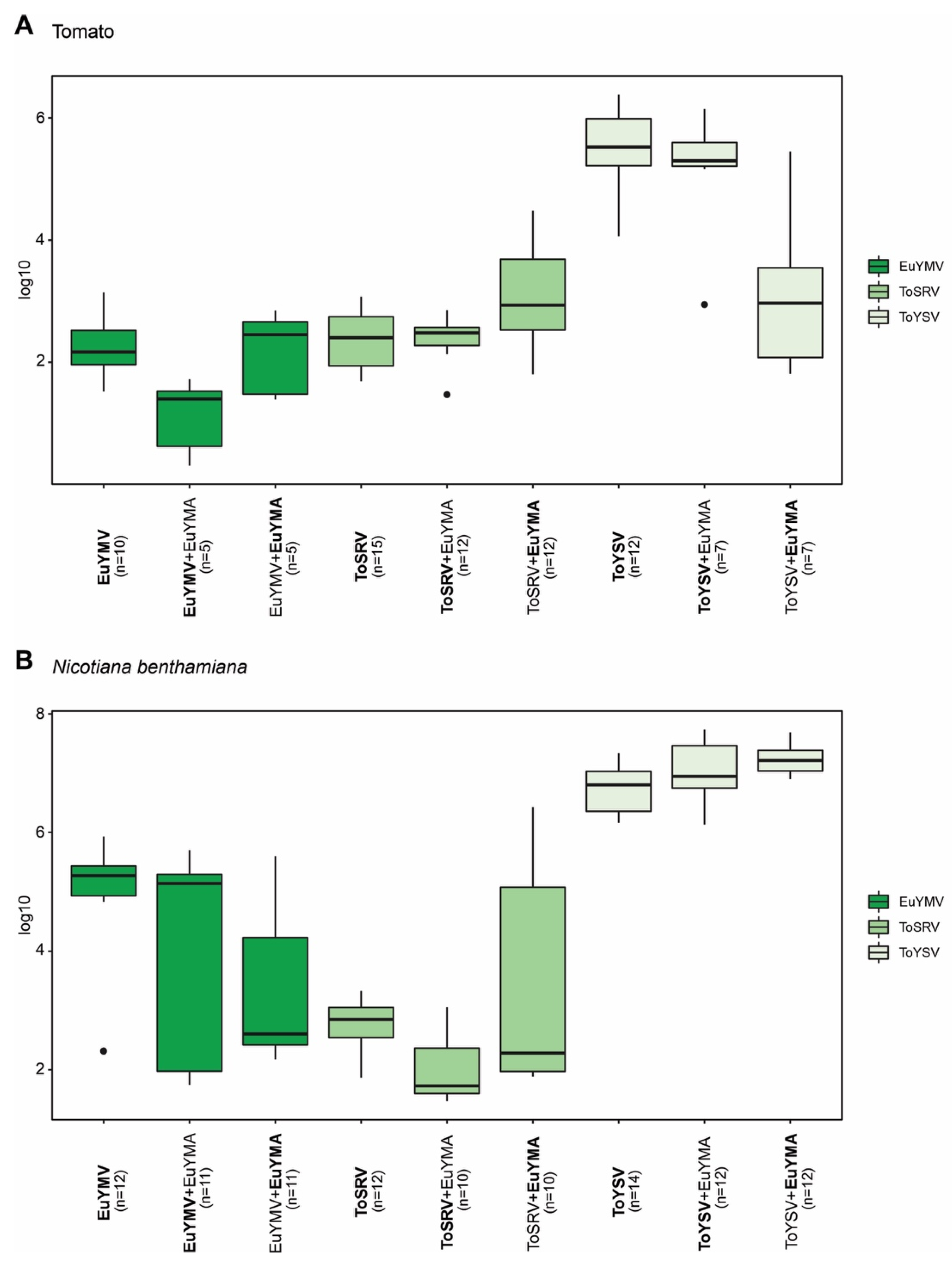
2.2. Effects of EuYMA on Infectivity, Symptoms and Accumulation of EuYMV, ToSRV and ToYSV
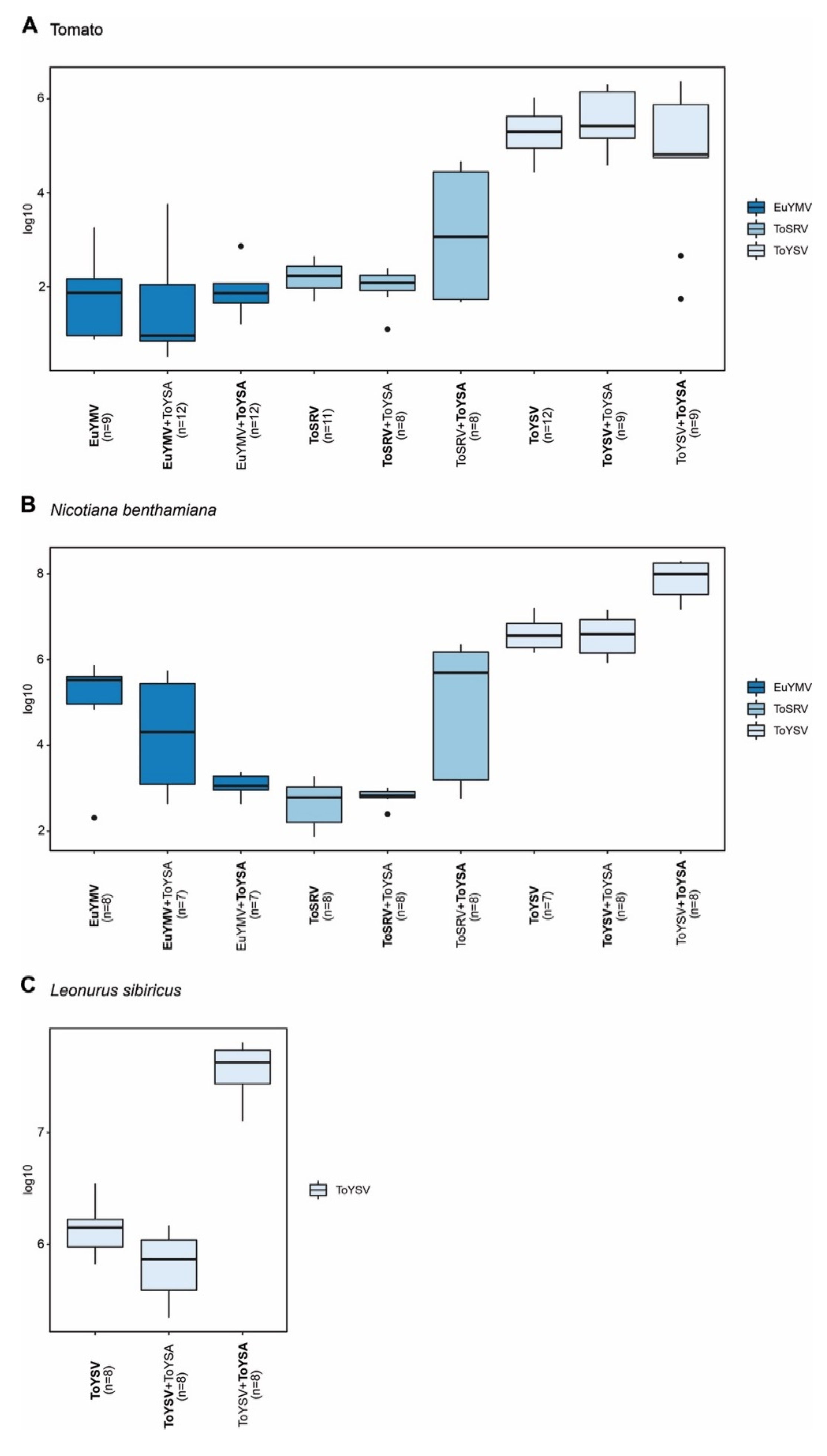
2.3. Effects of ToYSA on Infectivity, Symptoms and Accumulation of EuYMV, ToSRV and ToYSV
2.4. Effect of ToYSA on the Transmission of ToSRV by B. tabaci MEAM1
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analysis
4.2. Construction of the ToYSA Infectious Clone
4.3. Plant Inoculations
4.4. Detection and Quantification of Begomovirus and Alphasatellite Genomic Components
4.5. Whitefly Transmission Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varsani, A.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Brown, J.K.; Zerbini, F.M.; Martin, D.P. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Arch. Virol. 2014, 159, 2193–2203. [Google Scholar] [CrossRef]
- Varsani, A.; Roumagnac, P.; Fuchs, M.; Navas-Castillo, J.; Moriones, E.; Idris, A.; Briddon, R.W.; Rivera-Bustamante, R.F.; Zerbini, F.M.; Martin, D.P. Capulavirus and Grablovirus: Two new genera in the family Geminiviridae. Arch. Virol. 2017, 162, 1819–1831. [Google Scholar] [CrossRef] [Green Version]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Varsani, A.; ICTV Consortium. ICTV Virus Taxonomy Profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.A.; Rivera-Bustamante, R.F.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. A phylogenetic and evolutionary justification for three genera of Geminiviridae. Arch. Virol. 1994, 139, 49–77. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible emergence of new geminiviruses by frequent recombination. Virology 1999, 265, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paximadis, M.; Idris, A.M.; Torres-Jerez, I.; Villarreal, A.; Rey, M.E.C.; Brown, J.K. Characterization of tobacco geminiviruses in the Old and New world. Arch. Virol. 1999, 144, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant’s armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Mahajan, N.; Parameswari, C.; Veluthambi, K. Severe stunting in blackgram caused by the Mungbean yellow mosaic virus. (MYMV) KA27 DNA B component is ameliorated by co-infection or post-infection with the KA22 DNA B: MYMV nuclear shuttle protein is the symptom determinant. Virus. Res. 2011, 157, 25–34. [Google Scholar] [CrossRef]
- Brustolini, O.J.; Machado, J.P.; Condori-Apfata, J.A.; Coco, D.; Deguchi, M.; Loriato, V.A.; Pereira, W.A.; Alfenas-Zerbini, P.; Zerbini, F.M.; Inoue-Nagata, A.K.; et al. Sustained NIK-mediated antiviral signalling confers broad-spectrum tolerance to begomoviruses in cultivated plants. Plant. Biotechnol. J. 2015, 13, 1300–1311. [Google Scholar] [CrossRef] [Green Version]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. The role of AV2 (“precoat”) and coat protein in viral replication and movement in tomato leaf curl geminivirus. Virology 1996, 224, 390–404. [Google Scholar] [CrossRef] [Green Version]
- Mansoor, S.; Briddon, R.W.; Zafar, Y.; Stanley, J. Geminivirus disease complexes: An emerging threat. Trends Plant. Sci. 2003, 8, 128–134. [Google Scholar] [CrossRef]
- Zhou, X. Advances in understanding begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Fiallo-Olive, E.; Chirinos, D.; Geraud-Pouey, F.; Briddon, R.W.; Navas-Castillo, J. Characterization of non-coding DNA satellites associated with sweepoviruses (genus Begomovirus, Geminiviridae)—Definition of a distinct class of begomovirus-associated satellites. Front. Microbiol. 2016, 7, 162. [Google Scholar] [CrossRef] [Green Version]
- Paprotka, T.; Metzler, V.; Jeske, H. The first DNA 1-like alphasatellites in association with New World begomoviruses in natural infections. Virology 2010, 404, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romay, G.; Chirinos, D.; Geraud-Pouey, F.; Desbiez, C. Association of an atypical alphasatellite with a bipartite New World begomovirus. Arch. Virol. 2010, 155, 1843–1847. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Tovar, R.; Navas-Castillo, J. Deciphering the biology of deltasatellites from the New World: Maintenance by New World begomoviruses and whitefly transmission. New Phytol. 2016, 212, 680–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briddon, R.W.; Martin, D.P.; Roumagnac, P.; Navas-Castillo, J.; Fiallo-Olive, E.; Moriones, E.; Lett, J.M.; Zerbini, F.M.; Varsani, A. Alphasatellitidae: A new family with two subfamilies for the classification of geminivirus- and nanovirus-associated alphasatellites. Arch. Virol. 2018, 163, 2587–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, K.; Bedford, I.D.; Stanley, J. Adaptation from whitefly to leafhopper transmission of an autonomously replicating nanovirus-like DNA component associated with ageratum yellow vein disease. J. Gen. Virol. 2002, 83, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Mansoor, S.; Khan, S.H.; Bashir, A.; Saeed, M.; Zafar, Y.; Malik, K.A.; Briddon, R.; Stanley, J.; Markham, P.G. Identification of a novel circular single-stranded DNA associated with cotton leaf curl disease in Pakistan. Virology 1999, 259, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Saunders, K.; Stanley, J. A nanovirus-like DNA component associated with yellow vein disease of Ageratum conyzoides: Evidence for interfamilial recombination between plant DNA viruses. Virology 1999, 264, 142–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, K.; Bedford, I.D.; Briddon, R.W.; Markham, P.G.; Wong, S.M.; Stanley, J. A unique virus complex causes Ageratum yellow vein disease. Proc. Natl Acad Sci. USA 2000, 97, 6890–6895. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Wang, Z.Q.; Liu, X.; Zhao, L.; Zhou, X.; Xie, Y. Identification and analysis of potential genes regulated by an alphasatellite (TYLCCNA) that contribute to host resistance against tomato yellow leaf curl China Virus and its betasatellite (TYLCCNV/TYLCCNB) infection in Nicotiana benthamiana. Viruses 2019, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, Q.; Amin, I.; Mansoor, S.; Wassenegger, M.; Briddon, R.W. Geminivirus-associated alphasatellites suppress transcriptional not post-transcriptional gene silencing. Virus Dis. 2016, 27, 419. [Google Scholar]
- Jeske, H.; Kober, S.; Schäfer, B.; Strohmeier, S. Circomics of Cuban geminiviruses reveals the first alphasatellite DNA in the Caribbean. Virus Genes 2014, 49, 312–324. [Google Scholar] [CrossRef]
- Rosario, K.; Marr, C.; Varsani, A.; Kraberger, S.; Stainton, D.; Moriones, E.; Polston, J.E.; Breitbart, M. Begomovirus-associated satellite DNA diversity captured through vector-enabled metagenomic (VEM) surveys using whiteflies (Aleyrodidae). Viruses 2016, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Idris, A.M.; Shahid, M.S.; Briddon, R.W.; Khan, A.J.; Zhu, J.K.; Brown, J.K. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. J. Gen. Virol. 2011, 92, 706–717. [Google Scholar] [CrossRef]
- Mar, T.B.; Mendes, I.R.; Lau, D.; Fiallo-Olive, E.; Navas-Castillo, J.; Alves, M.S.; Zerbini, F.M. Interaction between the New World begomovirus Euphorbia yellow mosaic virus and its associated alphasatellite: Effects on infection and transmission by the whitefly Bemisia tabaci. J. Gen. Virol. 2017, 98, 1552–1562. [Google Scholar] [CrossRef]
- Ferro, C.G.; Silva, J.P.; Xavier, C.A.D.; Godinho, M.T.; Lima, A.T.M.; Mar, T.B.; Lau, D.; Zerbini, F.M. The ever increasing diversity of begomoviruses infecting non-cultivated hosts: New species from Sida spp. and Leonurus sibiricus, plus two New World alphasatellites. Ann. Appl. Biol. 2017, 170, 204–218. [Google Scholar] [CrossRef]
- Duarte, M.F.; Pereira Carvalho, R.C.; Dos Reis, L.N.A.; Rojas, M.J.; Gilbertson, R.; Costa, H.; Boiteux, L.S.; Fonseca, M.E.N. Natural infection of tomatoes (Solanum lycopersicum) by Euphorbia yellow mosaic virus isolates across four Brazilian states. Plant. Dis. 2020, 105, 518. [Google Scholar] [CrossRef]
- Catarino, A.M.; Sousa, T.F.; Lima, E.J.S.P.; Zerbini, F.M.; Sande, O.F.L.; Nascimento, M.B.; Cruz, J.C.; Hanada, R.E.; Nascimento, Á.R.; Assis, L.A.G.; et al. Molecular detection of Euphorbia yellow mosaic virus. infecting chili pepper. Trop. Plant. Pathol. 2020, 45, 454–460. [Google Scholar] [CrossRef]
- Rodríguez-Pardina, P.E.; Hanada, K.; Laguna, I.G.; Zerbini, F.M.; Ducasse, D.A. Molecular characterisation and relative incidence of bean- and soybean-infecting begomoviruses in northwestern Argentina. Ann. Appl. Biol. 2011, 158, 69–78. [Google Scholar] [CrossRef]
- Fernandes, N.A.N.; Boiteux, L.S.; Fonseca, M.E.N.; Gonzales-Segnana, L.; Kitajima, E.W. Report of Tomato yellow spot virus. infecting Leonurus sibiricus in Paraguay and within tomato fields in Brazil. Plant. Dis. 2014, 98, 1445. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.A.; Barreto, S.S.; Costa, T.M.; Rocha, G.A.; Dianese, E.C.; Gilbertson, R.L.; Inoue-Nagata, A.K. First report of Tomato severe rugose virus., a tomato-infecting begomovirus, in soybean plants in Brazil. Plant. Dis. 2017, 101, 1959. [Google Scholar] [CrossRef]
- Macedo, M.A.; Barreto, S.S.; Costa, T.M.; Maliano, M.R.; Rojas, M.R.; Gilbertson, R.L.; Inoue-Nagata, A.K. First report of common beans as a non-symptomatic host of Tomato severe rugose virus in Brazil. Plant. Dis. 2017, 101, 261. [Google Scholar] [CrossRef]
- Barbosa, J.C.; Barreto, S.S.; Inoue-Nagata, A.K.; Reis, M.S.; Firmino, A.C.; Bergamin, A.; Rezende, J.A.M. Natural infection of Nicandra physaloides by Tomato severe rugose virus in Brazil. J. Gen. Plant. Pathol. 2009, 75, 440–443. [Google Scholar] [CrossRef]
- Souza-Dias, J.A.C.; Sawazaki, H.E.; Pernambuco, P.C.A.; Elias, L.M.; Maluf, H. Tomato severe rugose virus: Another begomovirus causing leaf deformation and mosaic symptoms on potato in Brazil. Plant. Dis. 2008, 92, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, D.N.; Krause-Sakate, R.; Hasegawa, J.M.; Cezar, M.A.; Dziuba, P.H.; Pavan, M.A. First report of Tomato severe rugose virus infecting pepper plants in Brazil. Fitopatol. Bras. 2006, 31, 321. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.C.; Manhani, G.G.; Alfenas, P.F.; Calegario, R.F.; Fontes, E.P.B.; Zerbini, F.M. Tomato yellow spot Virus, a tomato-infecting begomovirus from Brazil with a closer relationship to viruses from Sida sp., forms pseudorecombinants with begomoviruses from tomato but not from Sida. J. Gen. Virol. 2006, 87, 3687–3696. [Google Scholar] [CrossRef]
- Silva, F.N.; Lima, A.T.M.; Rocha, C.S.; Castillo-Urquiza, G.P.; Alves-Júnior, M.; Zerbini, F.M. Recombination and pseudorecombination driving the evolution of the begomoviruses tomato severe rugose virus (ToSRV) and tomato rugose mosaic virus (ToRMV): Two recombinant DNA-A components sharing the same DNA-B. Virol. J. 2014, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- Calegario, R.F.; Ferreira, S.S.; Andrade, E.C.; Zerbini, F.M. Characterization of Tomato yellow spot virus. (ToYSV), a novel tomato-infecting begomovirus from Brazil. Pesqui. Agropecu. Bras. 2007, 42, 1335–1343. [Google Scholar] [CrossRef]
- Alves-Junior, M.; Alfenas-Zerbini, P.; Andrade, E.C.; Esposito, D.A.; Silva, F.N.; Cruz, A.C.F.; Ventrella, M.C.; Otoni, W.C.; Zerbini, F.M. Synergism and negative interference during co-infection of tomato and Nicotiana benthamiana with two bipartite begomoviruses. Virology 2009, 387, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Nawaz-ul-Rehman, M.S.; Nahid, N.; Mansoor, S.; Briddon, R.W.; Fauquet, C.M. Post-transcriptional gene silencing suppressor activity of two non-pathogenic alphasatellites associated with a begomovirus. Virology 2010, 405, 300–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kon, T.; Rojas, M.R.; Abdourhamane, I.K.; Gilbertson, R.L. Roles and interactions of begomoviruses and satellite DNAs associated with okra leaf curl disease in Mali, West Africa. J. Gen. Virol. 2009, 90, 1001–1013. [Google Scholar] [CrossRef]
- Mar, T.B.; Xavier, C.A.D.; Lima, A.T.M.; Nogueira, A.M.; Silva, J.C.F.; Ramos-Sobrinho, R.; Lau, D.; Zerbini, F.M. Genetic variability and population structure of the New World begomovirus Euphorbia yellow mosaic virus. J. Gen. Virol. 2017, 98, 1537–1551. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Inoue-Nagata, A.K.; Albuquerque, L.C.; Rocha, W.B.; Nagata, T. A simple method for cloning the complete begomovirus genome using the bacteriophage phi29 DNA polymerase. J. Virol. Met. 2004, 116, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.T.M. Caracterização de Dois Begomovírus (Tomato severe rugose Virus e Tomato yellow vein streak Virus) que Infectam Tomateiro e Obtenção de Clones Infecciosos. Master’s Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2007. [Google Scholar]
- Aragão, F.J.L.; Barros, L.M.G.; Brasileiro, A.C.M.; Ribeiro, S.G.; Smith, F.D.; Sanford, J.C.; Faria, J.C.; Rech, E.L. Inheritance of foreign genes in transgenic bean (Phaseolus vulgaris L.) co-transformed via particle bombardment. Theor Appl. Genet. 1996, 93, 142–150. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small amounts of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Royston, P. Algorithm AS 181: The W test for Normality. Appl. Stat. 1982, 31, 176–180. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 14 August 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tomato | ||||||
| EuYMV | ToSRV | ToYSV | ||||
| alone | +EuYMA | alone | +EuYMA | alone | +EuYMA | |
| Virus detection * | 11/25 # (44%) | 6/25 (24%) | 21/25 (84%) | 25/25 (100%) | 22/25 (88%) | 25/25 (100%) |
| EuYMA detection * | 6/25 (24%) | 18/25 (72%) | 13/25 (52%) | |||
| Nicotiana benthamiana | ||||||
| EuYMV | ToSRV | ToYSV | ||||
| alone | +EuYMA | alone | +EuYMA | alone | +EuYMA | |
| Virus detection | 15/21 (71%) | 18/19 (95%) | 18/18 (100%) | 15/21 (71%) | 19/20 (95%) | 20/20 (100%) |
| EuYMA detection | 12/19 (63%) | 14/21 (67%) | 19/20 (95%) | |||
| Tomato | ||||||
| EuYMV | ToSRV | ToYSV | ||||
| alone | +ToYSA | alone | +ToYSA | alone | +ToYSA | |
| Virus detection * | 22/40 # (55%) | 27/39 (69%) | 23/39 (59%) | 14/36 (39%) | 24/40 (60%) | 25/39 (64%) |
| ToYSA detection * | 26/39 (67%) | 7/36 (19%) | 15/39 (38%) | |||
| Nicotiana benthamiana | ||||||
| EuYMV | ToSRV | ToYSV | ||||
| alone | +ToYSA | alone | +ToYSA | alone | +ToYSA | |
| Virus detection | 13/24 (54%) | 13/23 (57%) | 17/21 (81%) | 14/20 (70%) | 21/23 (91%) | 25/25 (100%) |
| ToYSA detection | 10/23 (43%) | 12/20 (60%) | 22/25 (88%) | |||
| Leonurus sibiricus | ||||||
| EuYMV | ToSRV | ToYSV | ||||
| alone | +ToYSA | alone | +ToYSA | alone | +ToYSA | |
| Virus detection | 0/23 | 0/29 | 0/24 | 0/25 | 19/28 (68%) | 21/30 (70%) |
| ToYSA detection | n.d. ¥ | n.d. | 17/30 (57%) | |||
| Number of Infected Plants/Number of Inoculated Plants (%) | |||||
|---|---|---|---|---|---|
| Treatments | ToSRV * | ToSRV and ToYSA # | ToSRV and ToYSA ¥ | ToYSV & | ToYSV and ToYSA £ |
| Exp. 1 | 30/30 (100) | 13/28 (46) | 5/28 (18) | 0/15 (0) | 0/15 (0) |
| Exp. 2 | 22/25 (88) | 24/26 (92) | 0/26 (0) | n.d. $ | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogueira, A.M.; Nascimento, M.B.; Barbosa, T.M.C.; Quadros, A.F.F.; Gomes, J.P.A.; Orílio, A.F.; Barros, D.R.; Zerbini, F.M. The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission. Pathogens 2021, 10, 1244. https://doi.org/10.3390/pathogens10101244
Nogueira AM, Nascimento MB, Barbosa TMC, Quadros AFF, Gomes JPA, Orílio AF, Barros DR, Zerbini FM. The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission. Pathogens. 2021; 10(10):1244. https://doi.org/10.3390/pathogens10101244
Chicago/Turabian StyleNogueira, Angélica M., Monique B. Nascimento, Tarsiane M. C. Barbosa, Ayane F. F. Quadros, João Paulo A. Gomes, Anelise F. Orílio, Danielle R. Barros, and Francisco Murilo Zerbini. 2021. "The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission" Pathogens 10, no. 10: 1244. https://doi.org/10.3390/pathogens10101244
APA StyleNogueira, A. M., Nascimento, M. B., Barbosa, T. M. C., Quadros, A. F. F., Gomes, J. P. A., Orílio, A. F., Barros, D. R., & Zerbini, F. M. (2021). The Association between New World Alphasatellites and Bipartite Begomoviruses: Effects on Infection and Vector Transmission. Pathogens, 10(10), 1244. https://doi.org/10.3390/pathogens10101244