
A New Lineage of Perch Rhabdovirus Associated with Mortalities of Farmed Perch


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
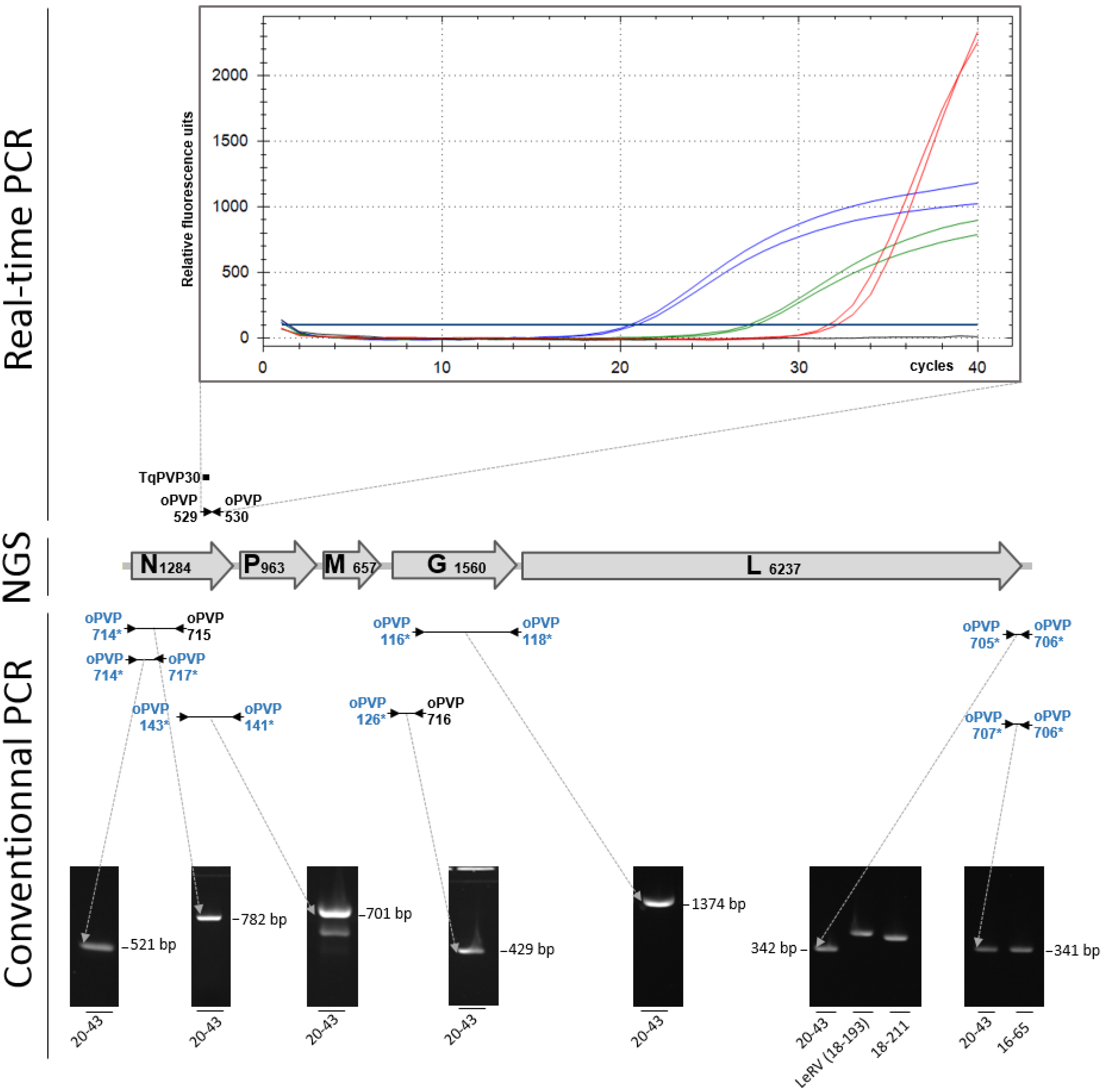
:1. Introduction
2. Results
2.1. Pathology
2.2. Virus Detection
2.3. Genome Sequence
2.4. Phylogenetic Studies
3. Discussion
4. Materials and Methods
4.1. Virus Isolation
4.2. Nucleic Acid Extraction, RT-PCR Detection and Sanger Sequencing
4.3. Next-Generation Sequencing
4.4. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gadd, T.; Viljamaa-Dirks, S.; Holopainen, R.; Koski, P.; Jakava-Viljanen, M. Characterization of perch rhabdovirus (PRV) in farmed grayling Thymallus thymallus. Dis. Aquat. Organ. 2013, 106, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbi, C.; Cabon, J.; Baud, M.; Bourjaily, M.; de Boisseson, C.; Castric, J.; Bigarré, L. Genetic diversity of perch rhabdoviruses isolates based on the nucleoprotein and glycoprotein genes. Arch. Virol. 2011, 156, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Betts, A.M.; Stone, D.M.; Way, K.; Torhy, C.; Chilmonczyk, S.; Benmansour, A.; de Kinkelin, P. Emerging vesiculo-type virus infections of freshwater fishes in Europe. Dis. Aquat. Organ. 2003, 57, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahli, T.; Bellec, L.; von Siebenthal, B.; Cabon, J.; Schmidt-Posthaus, H.; Morin, T. First isolation of a rhabdovirus from perch Perca fluviatilis in Switzerland. Dis. Aquat. Organ. 2015, 116, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ruane, N.M.; Rodger, H.D.; McCarthy, L.J.; Swords, D.; Dodge, M.; Kerr, R.C.; Henshilwood, K.; Stone, D.M. Genetic diversity and associated pathology of rhabdovirus infections in farmed and wild perch Perca fluviatilis in Ireland. Dis. Aquat. Organ. 2014, 112, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannevig, H.; Olesen, N.J.; Jentoft, S.; Kvellestad, A.; Taksdal, T.; Hastein, T. The first isolation of a rhabdovirus from perch (Perca fluviatilis) in Norway. Bull. Eur. Assoc. Fish Pathol. 2001, 21, 186–194. [Google Scholar]
- Bigarré, L.; Plassiart, G.; de Boisseson, C.; Pallandre, L.; Pozet, F.; Ledore, Y.; Fontaine, P.; Lieffrig, F. Molecular investigations of outbreaks of Perch perhabdovirus infections in pike-perch. Dis. Aquat. Organ. 2017, 127, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Nougayrède, P.; de Kinkelin, P.; Chilmonczyk, S.; Vuillaume, A. Isolation of a rhabdovirus from the pike-perch Stizostedion lucioperca (L.1758). Bull. Eur. Assoc. Fish Pathol. 1992, 12, 5–7. [Google Scholar]
- Dorson, M.; Torchy, C.; Chilmonczyk, S.; de Kinkelin, P. A rhabdovirus pathogenic for perch, Perca fluviatilis L.: Isolation and preliminary study. J. Fish Dis. 1984, 7, 241–245. [Google Scholar] [CrossRef]
- Jorgensen, P.E.V.; Olesen, N.J.; Ahne, W.; Wahli, T.; Meier, W. Isolation of a previously undescribed rhabdovirus from pike Esox lucius. Dis. Aquat. Organ. 1993, 16, 171–179. [Google Scholar] [CrossRef]
- Caruso, C.; Gustinelli, A.; Pastorino, P.; Acutis, P.L.; Prato, R.; Masoero, L.; Peletto, S.; Fioravanti, M.L.; Prearo, M. Mortality outbreak by perch rhabdovirus in European perch (Perca fluviatilis) farmed in Italy: Clinical presentation and phylogenetic analysis. J. Fish Dis. 2019, 42, 773–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallandre, L.; Luo, D.; Feuvrier, C.; Lieffrig, F.; Pozet, F.; Dacheux, L.; Bigarré, L. Revisiting the classification of percid perhabdoviruses using new full-length genomes. Viruses 2020, 12, 649. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.S.; Li, B.; Shen, X.R.; Jiang, R.D.; Zhu, Y.; Wu, J.; Fan, Y.; Bourhy, H.; Hu, B.; Ge, X.Y.; et al. Characterization of Novel Rhabdoviruses in Chinese Bats. Viruses 2021, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallandre, L.; Lautraite, A.; Feuvrier, C.; Pozet, F.; Dacheux, L.; Bigarré, L. A New Lineage of Perch Rhabdovirus Associated with Mortalities of Farmed Perch. Pathogens 2021, 10, 1256. https://doi.org/10.3390/pathogens10101256
Pallandre L, Lautraite A, Feuvrier C, Pozet F, Dacheux L, Bigarré L. A New Lineage of Perch Rhabdovirus Associated with Mortalities of Farmed Perch. Pathogens. 2021; 10(10):1256. https://doi.org/10.3390/pathogens10101256
Chicago/Turabian StylePallandre, Laurane, Armand Lautraite, Claudette Feuvrier, Françoise Pozet, Laurent Dacheux, and Laurent Bigarré. 2021. "A New Lineage of Perch Rhabdovirus Associated with Mortalities of Farmed Perch" Pathogens 10, no. 10: 1256. https://doi.org/10.3390/pathogens10101256
APA StylePallandre, L., Lautraite, A., Feuvrier, C., Pozet, F., Dacheux, L., & Bigarré, L. (2021). A New Lineage of Perch Rhabdovirus Associated with Mortalities of Farmed Perch. Pathogens, 10(10), 1256. https://doi.org/10.3390/pathogens10101256