
The Global Emergence of Human Babesiosis


Abstract
:1. Introduction

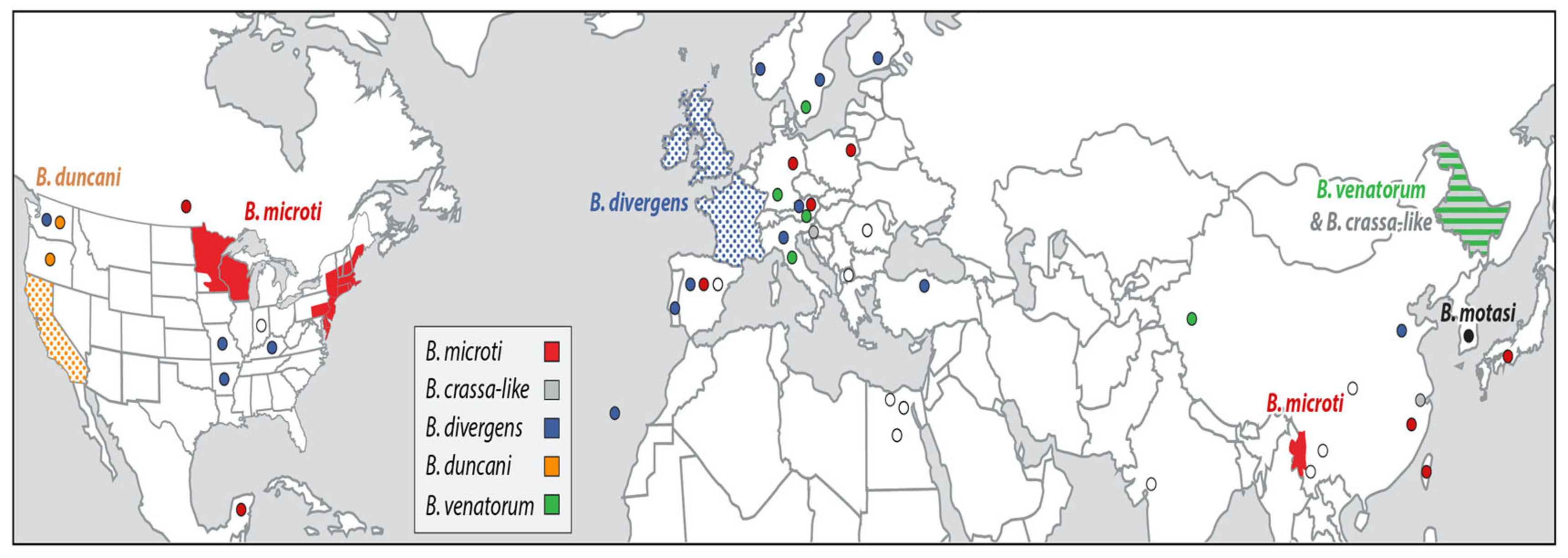{kind=link}
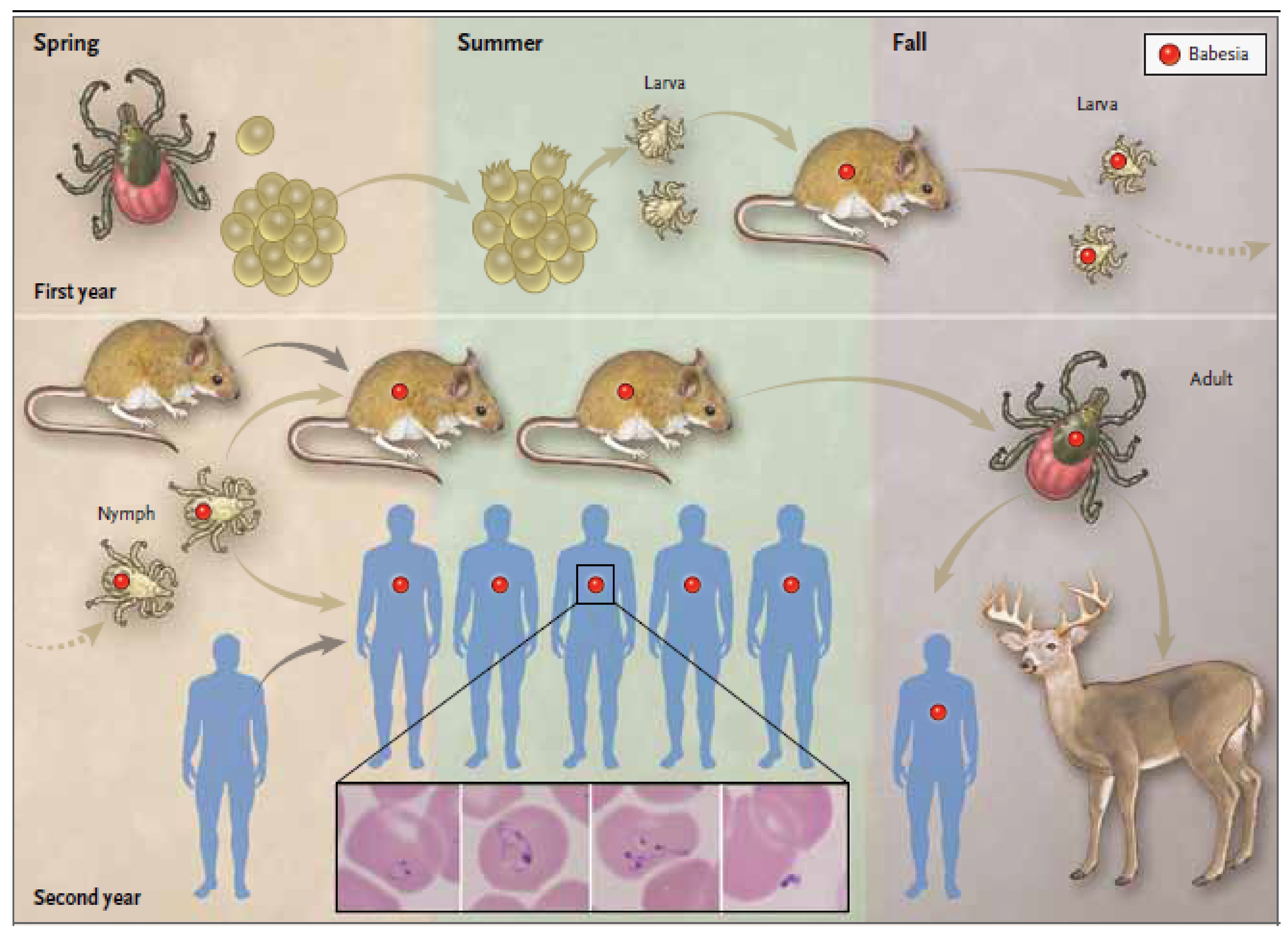{kind=link}
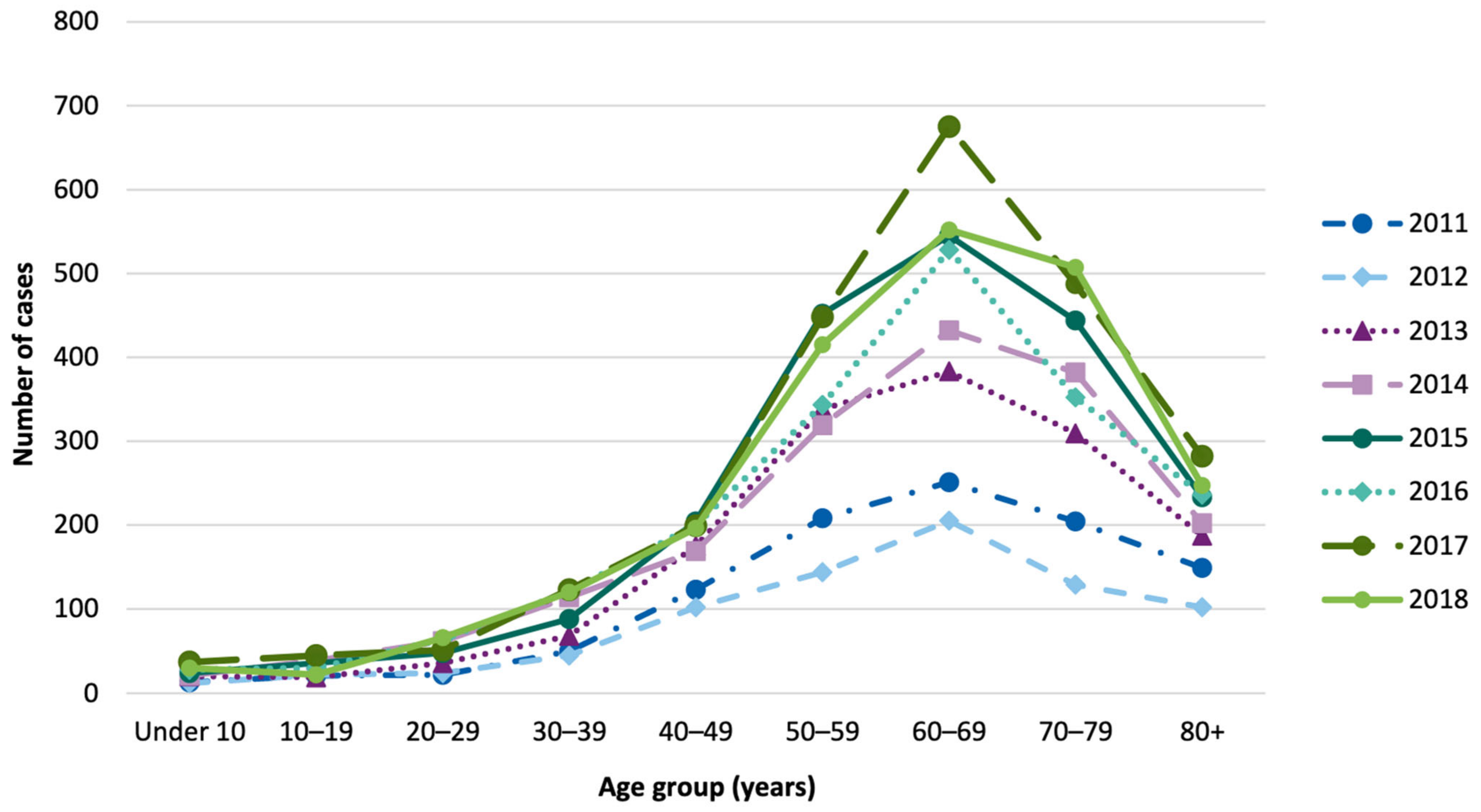{kind=link}
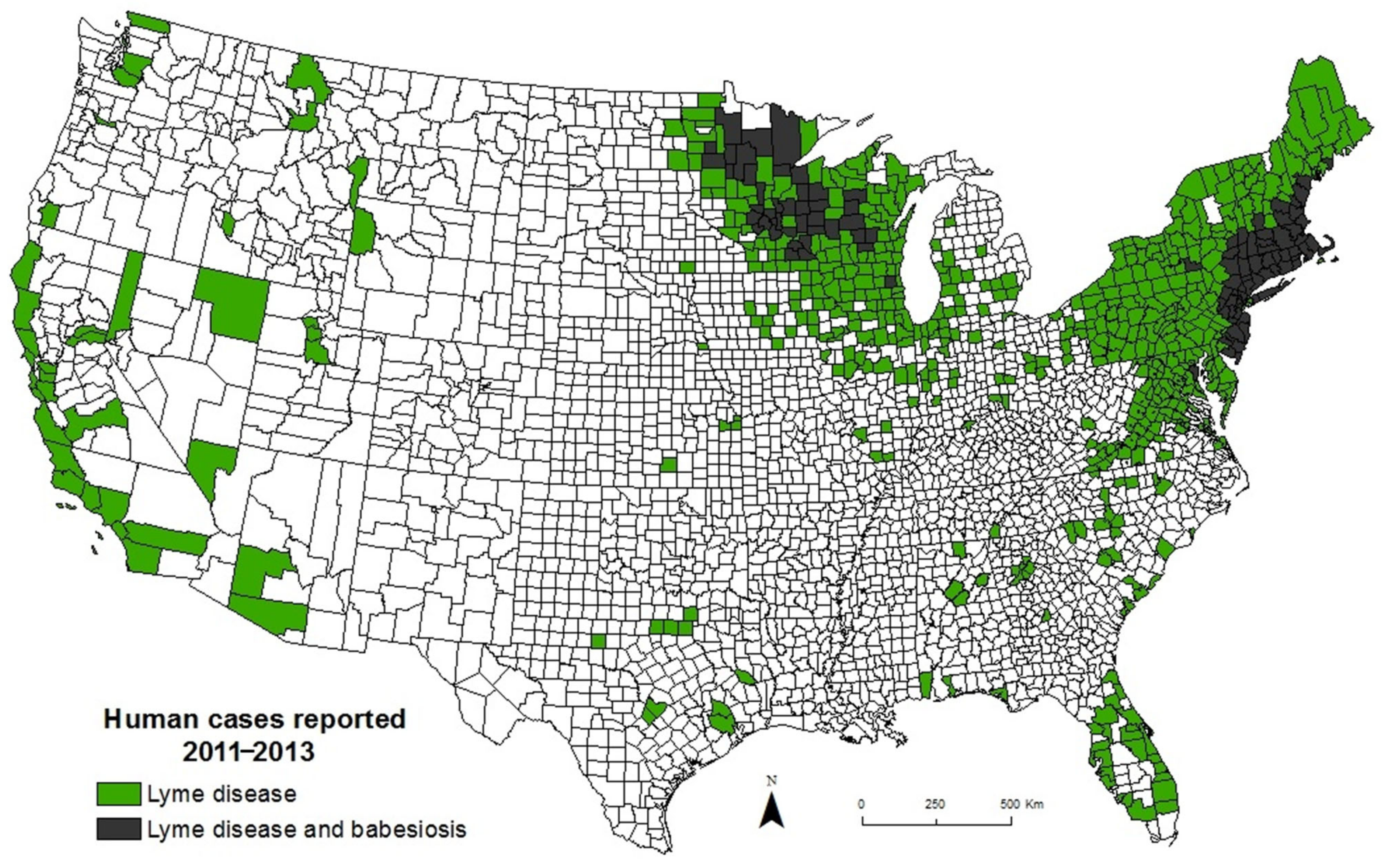{kind=link}
| Babesia Species | Year Case Reported | Major Region of Transmission | Primary Vector |
|---|---|---|---|
| Babesia microti | 1968 [32] | United States (Northeast, northern Midwest) | I. scapularis |
| Babesia divergens | 1957 [33] | Western Europe | I. ricinus |
| Babesia duncani | 1991 [34] | United States (Farwest) | D. albipictus |
| Babesia venatorum (EU1) | 2003 [35] | Europe (Austria, Italy) | I. ricinus |
| China | I. persulcatus | ||
| Babesia motasi (KO-1) | 2007 [26] | South Korea | unknown |
| Babesia crassa-like agent | 2018 [11] | Northeast China | I. persulcatus |
| Genetic variants | |||
| Babesia divergens-like | 1996 [36] | United States | Unknown |
| Babesia microti-like (TW1) | 1997 [30] | Taiwan, Japan | Unknown |
| Continent/Country | Causative Agent (Number of Cases) |
|---|---|
| Africa | Babesia spp. |
| Egypt | Babesia sp. (4) [21] |
| Mozambique | Babesia sp. (2) [8] |
| South Africa | Babesia sp. (2) [29] |
| Asia | B. crassa-like agent, B. divergens B. microti, B. motasi, B. venatorum |
| China | B. crassa-like agent, B. divergens B. microti, B. venatorum |
| India | Babesia sp. (1) [22,23] |
| Japan | B. microti (1) [24] |
| Korea | B. motasi (2) [25,26] |
| Mongolia | B. microti (3) [28] |
| Australia | B. microti |
| New South Wales | B. microti (1) [13] |
| Europe | B. crassa-like agent, B. divergens, B. microti, B. venatorum |
| Canary Islands (Spain) | B. divergens-like agent (1) [18] |
| North America | B. divergens-like, B. duncani, B. microti |
| United States | B. divergens-like, B. duncani, B. microti |
| Canada | B. microti (1), B. odocoilei (2) [16,17] |
| Mexico | B. microti (4), Babesia spp. (3) [27,59] |
| South America | B. microti |
| Bolivia | B. microti (9) [14] |
| Brazil | Babesia sp. (1) [15] |
| Colombia | Babesia sp. (1), B. bovis (4), B. bigemina (2) [19] |
| Ecuador | B. microti (1) [20] |
2. Epidemiologic Tools
2.1. Case Surveillance
2.2. Case Reports and Case Series
2.3. Serosurveys
2.4. Ecological Studies
Tick Vector and Mammalian Host Surveillance
2.5. Genomics
2.6. Mathematical Modeling
3. Modes of Transmission and Demographics of Human Babesiosis
4. Human Babesiosis in the Americas
4.1. Overview
4.2. United States
4.2.1. Babesia microti Infection
4.2.2. Babesia duncani Infection
4.2.3. Babesia divergens-Like Infection
4.2.4. Coinfection
4.3. Canada
4.4. Mexico
4.5. South America
5. Human Babesiosis in Europe
5.1. Overview
5.2. Babesia divergens
5.3. Babesia venatorum
5.4. Babesia microti
5.5. Babesia crassa-Like Agent
6. Babesiosis in Asia
6.1. Overview
6.2. China
6.2.1. Human Infection
6.2.2. Babesia venatorum
6.2.3. Babesia crassa-Like Agent
6.2.4. Babesia microti
6.2.5. Babesia divergens
6.2.6. Tick-Vectors and Animal Hosts of Babesia spp. in China
6.3. India
6.4. Japan
6.5. Korea
6.6. Mongolia
7. Babesiosis in Africa
8. Babesiosis in Australia
9. Conclusions
10. Patents
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vannier, E.; Krause, P.J. Human babesiosis. N. Engl. J. Med. 2012, 366, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Zintl, A.; Montero, E.; Hunfeld, K.P.; Gray, J. Human babesiosis in Europe. Pathogens 2021, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.Q.; Liu, K.; Li, X.L.; Liang, S.; Yang, Y.; Yao, H.W.; Sun, R.X.; Sun, Y.; Chen, W.J.; Zuo, S.Q.; et al. Emerging tick-borne infections in mainland China: An increasing public health threat. Lancet Infect. Dis. 2015, 15, 1467–1479. [Google Scholar] [CrossRef]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xia, S.; Huang, J.L.; Tambo, E.; Zhuge, H.X.; Zhou, X.N. Human babesiosis, an emerging tick-borne disease in the People’s Republic of China. Parasit. Vectors 2014, 7, 509. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., III; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Kjemtrup, A.M.; Conrad, P.A. Human babesiosis: An emerging tick-borne disease. Int. J. Parasitol. 2000, 30, 1323–1337. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Levine, N.D. Taxonomy of the Piroplasms. Trans. Am. Microsc. Soc. 1971, 90, 2–33. [Google Scholar] [CrossRef]
- Jia, N.; Zheng, Y.C.; Jiang, J.F.; Jiang, R.R.; Jiang, B.G.; Wei, R.; Liu, H.B.; Huo, Q.B.; Sun, Y.; Chu, Y.L.; et al. Human Babesiosis Caused by a Babesia crassa-Like Pathogen: A Case Series. Clin. Infect. Dis. 2018, 67, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.F.; Zheng, Y.C.; Jiang, R.R.; Li, H.; Huo, Q.B.; Jiang, B.G.; Sun, Y.; Jia, N.; Wang, Y.W.; Ma, L.; et al. Epidemiological, clinical, and laboratory characteristics of 48 cases of “Babesia venatorum” infection in China: A descriptive study. Lancet Infect. Dis. 2015, 15, 196–203. [Google Scholar] [CrossRef]
- Senanayake, S.N.; Paparini, A.; Latimer, M.; Andriolo, K.; Dasilva, A.J.; Wilson, H.; Xayavong, M.V.; Collignon, P.J.; Jeans, P.; Irwin, P.J. First report of human babesiosis in Australia. Med. J. Aust. 2012, 196, 350–352. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, S.; Totino, V.; Macchioni, F.; Zuñiga, F.; Rojas, P.; Lara, Y.; Roselli, M.; Bartoloni, A.; Cancrini, G. Human Babesiosis, Bolivia, 2013. Emerg. Infect. Dis. 2016, 22, 1445–1447. [Google Scholar] [CrossRef] [PubMed]
- Rech, A.; Bittar, C.M.; de Castro, C.G.; Azevedo, K.R.; dos Santos, R.P.; Machado, A.R.; Schwartsmann, G.; Goldani, L.; Brunetto, A.L. Asymptomatic babesiosis in a child with hepatoblastoma. J. Pediatr. Hematol. Oncol. 2004, 26, 213. [Google Scholar] [CrossRef]
- Kain, K.C.; Jassoum, S.B.; Fong, I.W.; Hannach, B. Transfusion-transmitted babesiosis in Ontario: First reported case in Canada. CMAJ 2001, 164, 1721–1723. [Google Scholar] [PubMed]
- Bullard, J.M.; Ahsanuddin, A.N.; Perry, A.M.; Lindsay, L.R.; Iranpour, M.; Dibernardo, A.; Van Caeseele, P.G. The first case of locally acquired tick-borne Babesia microti infection in Canada. Can. J. Infect. Dis. Med. Microbiol. 2014, 25, e87–e89. [Google Scholar] [CrossRef]
- Olmeda, A.S.; Armstrong, P.M.; Rosenthal, B.M.; Valladares, B.; del Castillo, A.; de Armas, F.; Miguelez, M.; González, A.; Rodríguez Rodríguez, J.A.; Spielman, A.; et al. A subtropical case of human babesiosis. Acta Trop. 1997, 67, 229–234. [Google Scholar] [CrossRef]
- Ríos, L.; Alvarez, G.; Blair, S. Serological and parasitological study and report of the first case of human babesiosis in Colombia. Rev. Soc. Bras. Med. Trop. 2003, 36, 493–498. [Google Scholar] [CrossRef]
- Al Zoubi, M.; Kwak, T.; Patel, J.; Kulkarni, M.; Kallal, C.A. Atypical challenging and first case report of babesiosis in Ecuador. IDCases 2016, 4, 15–17. [Google Scholar] [CrossRef]
- El-Bahnasawy, M.M.; Khalil, H.H.; Morsy, T.A. Babesiosis in an Egyptian boy aquired from pet dog, and a general review. J. Egypt. Soc. Parasitol. 2011, 41, 99–108. [Google Scholar] [PubMed]
- Marathe, A.; Tripathi, J.; Handa, V.; Date, V. Human babesiosis—A case report. Indian J. Med. Microbiol. 2005, 23, 267–269. [Google Scholar] [CrossRef]
- Negi, T.; Kandari, L.S.; Arunachalam, K. Update on prevalence and distribution pattern of tick-borne diseases among humans in India: A review. Parasitol. Res. 2021, 120, 1523–1539. [Google Scholar] [CrossRef] [PubMed]
- Saito-Ito, A.; Tsuji, M.; Wei, Q.; He, S.; Matsui, T.; Kohsaki, M.; Arai, S.; Kamiyama, T.; Hioki, K.; Ishihara, C. Transfusion-acquired, autochthonous human babesiosis in Japan: Isolation of Babesia microti-like parasites with hu-RBC-SCID mice. J. Clin. Microbiol. 2000, 38, 4511–4516. [Google Scholar] [CrossRef]
- Hong, S.H.; Kim, S.Y.; Song, B.G.; Rho, J.R.; Cho, C.R.; Kim, C.N.; Um, T.H.; Kwak, Y.G.; Cho, S.H.; Lee, S.E. Detection and characterization of an emerging type of Babesia sp. similar to Babesia motasi for the first case of human babesiosis and ticks in Korea. Emerg. Microbes Infect. 2019, 8, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Cho, S.H.; Joo, H.N.; Tsuji, M.; Cho, S.R.; Park, I.J.; Chung, G.T.; Ju, J.W.; Cheun, H.I.; Lee, H.W.; et al. First case of human babesiosis in Korea: Detection and characterization of a novel type of Babesia sp. (KO1) similar to ovine babesia. J. Clin. Microbiol. 2007, 45, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Peniche-Lara, G.; Balmaceda, L.; Perez-Osorio, C.; Munoz-Zanzi, C. Human babesiosis, Yucatán State, Mexico, 2015. Emerg. Infect. Dis. 2018, 24, 2061–2062. [Google Scholar] [CrossRef]
- Hong, S.H.; Anu, D.; Jeong, Y.I.; Abmed, D.; Cho, S.H.; Lee, W.J.; Lee, S.E. Molecular detection and seroprevalence of Babesia microti among stock farmers in Khutul City, Selenge Province, Mongolia. Korean J. Parasitol. 2014, 52, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.; Isaäcson, M.; Mohamed, A.; Potgieter, F.; De Waal, D. Human babesiosis-a preliminary report of 2 suspected cases in southern Africa. S. Afr. Med. J. 1990, 78, 699. [Google Scholar] [PubMed]
- Shih, C.M.; Liu, L.P.; Chung, W.C.; Ong, S.J.; Wang, C.C. Human babesiosis in Taiwan: Asymptomatic infection with a Babesia microti-like organism in a Taiwanese woman. J. Clin. Microbiol. 1997, 35, 450–454. [Google Scholar] [CrossRef]
- Tanyel, E.; Guler, N.; Hokelek, M.; Ulger, F.; Sunbul, M. A case of severe babesiosis treated successfully with exchange transfusion. Int. J. Infect. Dis. 2015, 38, 83–85. [Google Scholar] [CrossRef]
- Scholtens, R.G.; Braff, E.H.; Healey, G.A.; Gleason, N. A case of babesiosis in man in the United States. Am. J. Trop. Med. Hyg. 1968, 17, 810–813. [Google Scholar] [CrossRef]
- Skrabalo, Z.; Deanovic, Z. Piroplasmosis in man; report of a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar] [PubMed]
- Quick, R.E.; Herwaldt, B.L.; Thomford, J.W.; Garnett, M.E.; Eberhard, M.L.; Wilson, M.; Spach, D.H.; Dickerson, J.W.; Telford, S.R., III; Steingart, K.R.; et al. Babesiosis in Washington State: A new species of Babesia? Ann. Intern. Med. 1993, 119, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Cacciò, S.; Gherlinzoni, F.; Aspöck, H.; Slemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; et al. Molecular characterization of a non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.; Persing, D.H.; Précigout, E.A.; Goff, W.L.; Mathiesen, D.A.; Taylor, P.W.; Eberhard, M.L.; Gorenflot, A.F. A fatal case of babesiosis in Missouri: Identification of another piroplasm that infects humans. Ann. Intern. Med. 1996, 124, 643–650. [Google Scholar] [CrossRef]
- Puri, A.; Bajpai, S.; Meredith, S.; Aravind, L.; Krause, P.J.; Kumar, S. Babesia microti: Pathogen genomics, genetic variability, immunodominant antigens, and pathogenesis. Front. Microbiol. 2021, 12, 2416. [Google Scholar] [CrossRef]
- Babes, V. On bacterial haemoglobinuria in cattle. Comptes Rendus l’Acad. Sci. 1888, 107, 692–694. [Google Scholar]
- Spielman, A.; Wilson, M.L.; Levine, J.F.; Piesman, J. Ecology of Ixodes dammini-borne human babesiosis and Lyme disease. Annu. Rev. Entomol. 1985, 30, 439–460. [Google Scholar] [CrossRef] [PubMed]
- Western, K.A.; Benson, G.D.; Gleason, N.N.; Healy, G.R.; Schultz, M.G. Babesiosis in a Massachusetts resident. N. Engl. J. Med. 1970, 283, 854–856. [Google Scholar] [CrossRef]
- Walter, K.S.; Pepin, K.M.; Webb, C.T.; Gaff, H.D.; Krause, P.J.; Pitzer, V.E.; Diuk-Wasser, M.A. Invasion of two tick-borne diseases across New England: Harnessing human surveillance data to capture underlying ecological invasion processes. Proc. Biol. Sci. 2016, 283, 20160834. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.T.; John, M.; Visintainer, P.; Wormser, G.P. Increasing incidence and changing epidemiology of babesiosis in the Hudson Valley region of New York State: 2009–2016. Diagn. Microbiol. Infect. Dis. 2020, 96, 114958. [Google Scholar] [CrossRef] [PubMed]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes tick-borne pathogens: Ecological, epidemiological, and clinical consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Carpi, G.; Walter, K.S.; Mamoun, C.B.; Krause, P.J.; Kitchen, A.; Lepore, T.J.; Dwivedi, A.; Cornillot, E.; Caccone, A.; Diuk-Wasser, M.A. Babesia microti from humans and ticks hold a genomic signature of strong population structure in the United States. BMC Genom. 2016, 17, 888. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.M.; Krause, P.J.; Davis, S.; Vannier, E.G.; Fitzpatrick, M.C.; Rollend, L.; Belperron, A.A.; States, S.L.; Stacey, A.; Bockenstedt, L.K.; et al. Borrelia burgdorferi promotes the establishment of Babesia microti in the northeastern United States. PLoS ONE 2014, 9, e115494. [Google Scholar] [CrossRef]
- Menis, M.; Whitaker, B.I.; Wernecke, M.; Jiao, Y.; Eder, A.; Kumar, S.; Xu, W.; Liao, J.; Wei, Y.; MaCurdy, T.E.; et al. Babesiosis Occurrence Among United States Medicare Beneficiaries, Ages 65 and Older, During 2006–2017: Overall and by State and County of Residence. Open Forum Infect. Dis. 2021, 8, ofaa608. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.R., III; Spielman, A.; Sikand, V.; Ryan, R.; Christianson, D.; Burke, G.; Brassard, P.; Pollack, R.; Peck, J.; et al. Concurrent Lyme disease and babesiosis. Evidence for increased severity and duration of illness. JAMA 1996, 275, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Conrad, P.A.; Kjemtrup, A.M.; Carreno, R.A.; Thomford, J.; Wainwright, K.; Eberhard, M.; Quick, R.; Telford, S.R., III; Herwaldt, B.L. Description of Babesia duncani n.sp. (Apicomplexa: Babesiidae) from humans and its differentiation from other piroplasms. Int. J. Parasitol. 2006, 36, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.F.; Michelson, M.L.; Holman, P.J. Acute babesiosis caused by Babesia divergens in a resident of Kentucky. N. Engl. J. Med. 2002, 347, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; de Bruyn, G.; Pieniazek, N.J.; Homer, M.; Lofy, K.H.; Slemenda, S.B.; Fritsche, T.R.; Persing, D.H.; Limaye, A.P. Babesia divergens-like infection, Washington State. Emerg. Infect. Dis. 2004, 10, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Burgess, M.J.; Rosenbaum, E.R.; Pritt, B.S.; Haselow, D.T.; Ferren, K.M.; Alzghoul, B.N.; Rico, J.C.; Sloan, L.M.; Ramanan, P.; Purushothaman, R.; et al. Possible Transfusion-Transmitted Babesia divergens-like/MO-1 Infection in an Arkansas Patient. Clin. Infect. Dis. 2017, 64, 1622–1625. [Google Scholar] [CrossRef] [PubMed]
- Herc, E.; Pritt, B.; Huizenga, T.; Douce, R.; Hysell, M.; Newton, D.; Sidge, J.; Losman, E.; Sherbeck, J.; Kaul, D.R. Probable Locally Acquired Babesia divergens-Like Infection in Woman, Michigan, USA. Emerg. Infect. Dis. 2018, 24, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Ruebush, T.K., 2nd; Juranek, D.D.; Chisholm, E.S.; Snow, P.C.; Healy, G.R.; Sulzer, A.J. Human babesiosis on Nantucket Island. Evidence for self-limited and subclinical infections. N. Engl. J. Med. 1977, 297, 825–827. [Google Scholar] [CrossRef]
- Krause, P.J.; Auwaerter, P.G.; Bannuru, R.R.; Branda, J.A.; Falck-Ytter, Y.T.; Lantos, P.M.; Lavergne, V.; Meissner, H.C.; Osani, M.C.; Rips, J.G.; et al. Clinical Practice Guidelines by the Infectious Diseases Society of America (IDSA): 2020 Guideline on Diagnosis and Management of Babesiosis. Clin. Infect. Dis. 2021, 72, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Grabias, B.; Clement, J.; Krause, P.J.; Lepore, T.; Kumar, S. Superior real-time polymerase chain reaction detection of Babesia microti parasites in whole blood utilizing high-copy BMN antigens as amplification targets. Transfusion 2018, 58, 1924–1932. [Google Scholar] [CrossRef]
- Krause, P.J.; Telford, S., 3rd; Spielman, A.; Ryan, R.; Magera, J.; Rajan, T.V.; Christianson, D.; Alberghini, T.V.; Bow, L.; Persing, D. Comparison of PCR with blood smear and inoculation of small animals for diagnosis of Babesia microti parasitemia. J. Clin. Microbiol. 1996, 34, 2791–2794. [Google Scholar] [CrossRef]
- Smith, R.P.; Hunfeld, K.P.; Krause, P.J. Management strategies for human babesiosis. Expert Rev. Anti-Infect. Ther. 2020, 18, 625–636. [Google Scholar] [CrossRef]
- Krause, P.J.; Gewurz, B.E.; Hill, D.; Marty, F.M.; Vannier, E.; Foppa, I.M.; Furman, R.R.; Neuhaus, E.; Skowron, G.; Gupta, S.; et al. Persistent and relapsing babesiosis in immunocompromised patients. Clin. Infect. Dis. 2008, 46, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Osorno, B.M.; Vega, C.; Ristic, M.; Robles, C.; Ibarra, S. Isolation of Babesia spp. from asymptomatic human beings. Vet. Parasitol. 1976, 2, 111–120. [Google Scholar] [CrossRef]
- Krause, P.J.; McKay, K.; Gadbaw, J.; Christianson, D.; Closter, L.; Lepore, T.; Telford, S.R., III; Sikand, V.; Ryan, R.; Persing, D.; et al. Increasing health burden of human babesiosis in endemic sites. Am. J. Trop. Med. Hyg. 2003, 68, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Hai, V.V.; Almeras, L.; Socolovschi, C.; Raoult, D.; Parola, P.; Pagès, F. Monitoring human tick-borne disease risk and tick bite exposure in Europe: Available tools and promising future methods. Ticks Tick-Borne Dis. 2014, 5, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Lesko, C.R.; Keil, A.P.; Edwards, J.K. The Epidemiologic Toolbox: Identifying, Honing, and Using the Right Tools for the Job. Am. J. Epidemiol. 2020, 189, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N.C.; Porter, W.T.; Wachara, J.C.; Lowrey, T.J.; Martin, L.; Motyka, P.J.; Salkeld, D.J. Using citizen science to describe the prevalence and distribution of tick bite and exposure to tick-borne diseases in the United States. PLoS ONE 2018, 13, e0199644. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, J.P. In defense of case reports and case series. Ann. Intern. Med. 2001, 134, 330–334. [Google Scholar] [CrossRef]
- Ruebush, T.K., 2nd; Juranek, D.D.; Spielman, A.; Piesman, J.; Healy, G.R. Epidemiology of human babesiosis on Nantucket Island. Am. J. Trop. Med. Hyg. 1981, 30, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Kavathas, P.B.; Ruddle, N.H. Immunoepidemiology; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar]
- Lantos, P.M.; Rumbaugh, J.; Bockenstedt, L.K.; Falck-Ytter, Y.T.; Aguero-Rosenfeld, M.E.; Auwaerter, P.G.; Baldwin, K.; Bannuru, R.R.; Belani, K.K.; Bowie, W.R.; et al. Clinical Practice Guidelines by the Infectious Diseases Society of America, American Academy of Neurology, and American College of Rheumatology: 2020 Guidelines for the Prevention, Diagnosis, and Treatment of Lyme Disease. Neurology 2021, 96, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.E.; Williamson, P.C.; Bloch, E.M.; Clifford, J.; Cyrus, S.; Shaz, B.H.; Kessler, D.; Gorlin, J.; Erwin, J.L.; Krueger, N.X.; et al. Serologic screening of United States blood donors for Babesia microti using an investigational enzyme immunoassay. Transfusion 2016, 56, 1866–1874. [Google Scholar] [CrossRef]
- Niccolai, L.M.; Ruddle, N.H.; Krause, P.J. Introduction to immunology, epidemiology, and immunoepidemiology. In Immunoepidemiology; Krause, P.J., Kavathas, P.B., Ruddle, N.H., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–17. [Google Scholar]
- Diuk-Wasser, M.A.; Liu, Y.; Steeves, T.K.; Folsom-O’Keefe, C.; Dardick, K.R.; Lepore, T.; Bent, S.J.; Usmani-Brown, S.; Telford, S.R., III; Fish, D.; et al. Monitoring human babesiosis emergence through vector surveillance New England, USA. Emerg. Infect. Dis. 2014, 20, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.P., Jr.; Elias, S.P.; Borelli, T.J.; Missaghi, B.; York, B.J.; Kessler, R.A.; Lubelczyk, C.B.; Lacombe, E.H.; Hayes, C.M.; Coulter, M.S.; et al. Human babesiosis, Maine, USA, 1995–2011. Emerg. Infect. Dis. 2014, 20, 1727–1730. [Google Scholar] [CrossRef]
- Goethert, H.K.; Mather, T.N.; Buchthal, J.; Telford, S.R., III. Retrotransposon-based blood meal analysis of nymphal deer ticks demonstrates spatiotemporal diversity of Borrelia burgdorferi and Babesia microti reservoirs. Appl. Environ. Microbiol. 2021, 87, e02370-20. [Google Scholar] [CrossRef]
- Barbour, A.G.; Bunikis, J.; Travinsky, B.; Hoen, A.G.; Diuk-Wasser, M.A.; Fish, D.; Tsao, J.I. Niche partitioning of Borrelia burgdorferi and Borrelia miyamotoi in the same tick vector and mammalian reservoir species. Am. J. Trop. Med. Hyg. 2009, 81, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Wagemakers, A.; Jahfari, S.; de Wever, B.; Spanjaard, L.; Starink, M.V.; de Vries, H.J.C.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi in vectors and hosts in The Netherlands. Ticks Tick-Borne Dis. 2017, 8, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.B.; Herwaldt, B.L. Babesiosis Surveillance-United States, 2011–2015. MMWR Surveill. Summ. 2019, 68, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.Y.; Jiang, B.G.; Yuan, T.T.; von Fricken, M.; Jia, N.; Jiang, R.R.; Zhang, Y.; Li, X.L.; Sun, Y.; Ruan, X.D.; et al. Genetic Diversity and Coexistence of Babesia in Ticks (Acari: Ixodidae) from Northeastern China. Vector Borne Zoonotic Dis. 2020, 20, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Hickling, G.J.; Burke, R.L.; Ogden, N.H.; Beati, L.; LeBrun, R.A.; Arsnoe, I.M.; Gerhold, R.; Han, S.; Jackson, K.; et al. Why Lyme disease is common in the northern US, but rare in the south: The roles of host choice, host-seeking behavior, and tick density. PLoS Biol. 2021, 19, e3001066. [Google Scholar] [CrossRef]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B.; Augagneur, Y.; Bres, V.; Duclos, A.; Randazzo, S.; et al. Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 9102–9114. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Cornillot, E.; McCracken, C.; Usmani-Brown, S.; Dwivedi, A.; Ifeonu, O.O.; Crabtree, J.; Gotia, H.T.; Virji, A.Z.; Reynes, C.; et al. Genome-wide diversity and gene expression profiling of Babesia microti isolates identify polymorphic genes that mediate host-pathogen interactions. Sci. Rep. 2016, 6, 35284. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, J.E.; Tran, A.D.; Freimark, L.; Schaffner, S.F.; Goethert, H.; Andersen, K.G.; Bazner, S.; Li, A.; McGrath, G.; Sloan, L.; et al. A global map of genetic diversity in Babesia microti reveals strong population structure and identifies variants associated with clinical relapse. Nat. Microbiol. 2016, 1, 16079. [Google Scholar] [CrossRef] [PubMed]
- Goethert, H.K.; Telford, S.R., III. Not “out of Nantucket”: Babesia microti in southern New England comprises at least two major populations. Parasit. Vectors 2014, 7, 546. [Google Scholar] [CrossRef] [PubMed]
- Skrip, L.A.; Townsend, J.P. Modeling Approaches Toward Understanding Infectious Disease Transmission. In Immunoepidemiology; Krause, P.J., Kavathas, P.B., Ruddle, N.H., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 227–243. [Google Scholar] [CrossRef]
- Garner, M.G.; Hamilton, S.A. Principles of epidemiological modelling. Rev. Sci. Tech. 2011, 30, 407–416. [Google Scholar] [CrossRef]
- Hersh, M.H.; Tibbetts, M.; Strauss, M.; Ostfeld, R.S.; Keesing, F. Reservoir competence of wildlife host species for Babesia microti. Emerg. Infect. Dis. 2012, 18, 1951–1957. [Google Scholar] [CrossRef]
- Tufts, D.M.; Diuk-Wasser, M.A. Vertical Transmission: A Vector-Independent Transmission Pathway of Babesia microti in the Natural Reservoir Host Peromyscus leucopus. J. Infect. Dis. 2021, 223, 1787–1795. [Google Scholar] [CrossRef]
- Herwaldt, B.L.; Linden, J.V.; Bosserman, E.; Young, C.; Olkowska, D.; Wilson, M. Transfusion-associated babesiosis in the United States: A description of cases. Ann. Intern. Med. 2011, 155, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Cornett, J.K.; Malhotra, A.; Hart, D. Vertical Transmission of Babesiosis From a Pregnant, Splenectomized Mother to Her Neonate. Infect. Dis. Clin. Pract. 2012, 20, 408–410. [Google Scholar] [CrossRef]
- Brennan, M.B.; Herwaldt, B.L.; Kazmierczak, J.J.; Weiss, J.W.; Klein, C.L.; Leith, C.P.; He, R.; Oberley, M.J.; Tonnetti, L.; Wilkins, P.P.; et al. Transmission of Babesia microti Parasites by Solid Organ Transplantation. Emerg. Infect. Dis. 2016, 22, 1869–1876. [Google Scholar] [CrossRef]
- Bloch, E.M.; Krause, P.J.; Tonnetti, L. Preventing Transfusion-Transmitted Babesiosis. Pathogens 2021, 10, 1176. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Chawla, A.; Berardi, V.; Padbury, J.; Skowron, G.; Krause, P.J. Preventing transfusion-transmitted babesiosis: Preliminary experience of the first laboratory-based blood donor screening program. Transfusion 2012, 52, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.D.; Winton, C.S.; Tonnetti, L.; Townsend, R.L.; Berardi, V.P.; Hewins, M.E.; Weeks, K.E.; Dodd, R.Y.; Stramer, S.L. Screening for Babesia microti in the U.S. blood supply. N. Engl. J. Med. 2016, 375, 2236–2245. [Google Scholar] [CrossRef] [PubMed]
- Saetre, K.; Godhwani, N.; Maria, M.; Patel, D.; Wang, G.; Li, K.I.; Wormser, G.P.; Nolan, S.M. Congenital babesiosis after maternal infection with Borrelia burgdorferi and Babesia microti. J. Pediatric. Infect. Dis. Soc. 2018, 7, e1–e5. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.R., III; Pollack, R.J.; Ryan, R.; Brassard, P.; Zemel, L.; Spielman, A. Babesiosis: An underdiagnosed disease of children. Pediatrics 1992, 89, 1045–1048. [Google Scholar] [PubMed]
- Notifiable Diseases and Mortality Tables. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, Nd-38.
- Joseph, J.T.; Roy, S.S.; Shams, N.; Visintainer, P.; Nadelman, R.B.; Hosur, S.; Nelson, J.; Wormser, G.P. Babesiosis in Lower Hudson Valley, New York, USA. Emerg. Infect. Dis. 2011, 17, 843–847. [Google Scholar] [CrossRef]
- Rodgers, S.E.; Mather, T.N. Human Babesia microti incidence and Ixodes scapularis distribution, Rhode Island, 1998–2004. Emerg. Infect. Dis. 2007, 13, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Fida, M.; Challener, D.; Hamdi, A.; O’Horo, J.; Abu Saleh, O. Babesiosis: A Retrospective Review of 38 Cases in the Upper Midwest. Open Forum Infect. Dis. 2019, 6, ofz311. [Google Scholar] [CrossRef] [PubMed]
- Mareedu, N.; Schotthoefer, A.M.; Tompkins, J.; Hall, M.C.; Fritsche, T.R.; Frost, H.M. Risk Factors for Severe Infection, Hospitalization, and Prolonged Antimicrobial Therapy in Patients with Babesiosis. Am. J. Trop. Med. Hyg. 2017, 97, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Talarico, J.; Chang, H.G.; Birkhead, G.S.; Heimberger, T.; Morse, D.L. Human babesiosis in New York State: Review of 139 hospitalized cases and analysis of prognostic factors. Arch. Intern. Med. 1998, 158, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.R., III; Ryan, R.; Hurta, A.B.; Kwasnik, I.; Luger, S.; Niederman, J.; Gerber, M.; Spielman, A. Geographical and temporal distribution of babesial infection in Connecticut. J. Clin. Microbiol. 1991, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rand, P.W.; Lubelczyk, C.; Holman, M.S.; Lacombe, E.H.; Smith, R.P., Jr. Abundance of Ixodes scapularis (Acari: Ixodidae) after the complete removal of deer from an isolated offshore island, endemic for Lyme Disease. J. Med. Entomol. 2004, 41, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, H.J.; LaBonte, A.M.; Stafford, K.C. The relationship between deer density, tick abundance, and human cases of Lyme disease in a residential community. J. Med. Entomol. 2014, 51, 777–784. [Google Scholar] [CrossRef]
- Wilson, M.L.; Telford, S.R., III; Piesman, J.; Spielman, A. Reduced abundance of immature Ixodes dammini (Acari: Ixodidae) following elimination of deer. J. Med. Entomol. 1988, 25, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.A.; Berrang-Ford, L.; Buck, P.A.; Drebot, M.A.; Lindsay, L.R.; Ogden, N.H. Major emerging vector-borne zoonotic diseases of public health importance in Canada. Emerg. Microbes Infect. 2015, 4, e33. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; AbdelMalik, P.; Pulliam, J. Emerging infectious diseases: Prediction and detection. Can. Commun. Dis. Rep. 2017, 43, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, J.C.; Greenberg, P.D.; Antique, J.; Jimenez-Lucho, V.E. Severe babesiosis in Long Island: Review of 34 cases and their complications. Clin. Infect. Dis. 2001, 32, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.G.; Diuk-Wasser, M.A.; Ben Mamoun, C.; Krause, P.J. Babesiosis. Infect. Dis. Clin. N. Am. 2015, 29, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.M.; Kumar, S.; Krause, P.J. Persistence of Babesia microti Infection in Humans. Pathogens 2019, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Raffalli, J.; Wormser, G.P. Persistence of babesiosis for >2 years in a patient on rituximab for rheumatoid arthritis. Diagn. Microbiol. Infect. Dis. 2016, 85, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.S.; Westblade, L.F.; Dziedziech, A.; Visone, J.E.; Furman, R.R.; Jenkins, S.G.; Schuetz, A.N.; Kirkman, L.A. Clinical and Molecular Evidence of Atovaquone and Azithromycin Resistance in Relapsed Babesia microti Infection Associated with Rituximab and Chronic Lymphocytic Leukemia. Clin. Infect. Dis. 2017, 65, 1222–1225. [Google Scholar] [CrossRef] [PubMed]
- Wormser, G.P.; Prasad, A.; Neuhaus, E.; Joshi, S.; Nowakowski, J.; Nelson, J.; Mittleman, A.; Aguero-Rosenfeld, M.; Topal, J.; Krause, P.J. Emergence of resistance to azithromycin-atovaquone in immunocompromised patients with Babesia microti infection. Clin. Infect. Dis. 2010, 50, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B.; et al. Evidence for transmission of the zoonotic apicomplexan parasite Babesia duncani by the tick Dermacentor albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, E.J.; Lowenstine, L.J.; Hemmer, R.; Robinson, T.; Conrad, P.A. Comparative pathogenesis of human WA1 and Babesia microti isolates in a Syrian hamster model. Lab. Anim. Sci. 1996, 46, 507–515. [Google Scholar]
- Hemmer, R.M.; Wozniak, E.J.; Lowenstine, L.J.; Plopper, C.G.; Wong, V.; Conrad, P.A. Endothelial cell changes are associated with pulmonary edema and respiratory distress in mice infected with the WA1 human Babesia parasite. J. Parasitol. 1999, 85, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Caulfield, A.J.; Pritt, B.S. Lyme Disease Coinfections in the United States. Clin. Lab. Med. 2015, 35, 827–846. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.C.; Kain, K.C. Two tick-borne diseases in one: A case report of concurrent babesiosis and Lyme disease in Ontario. Cmaj 1999, 160, 1851–1853. [Google Scholar]
- Scott, J.D.; Sajid, M.S.; Pascoe, E.L.; Foley, J.E. Detection of Babesia odocoilei in humans with babesiosis symptoms. Diagnostics 2021, 11, 947. [Google Scholar] [CrossRef]
- Gonzalez, J.; Echaide, I.; Pabón, A.; Gabriel Piñeros, J.J.; Blair, S.; Tobón-Castaño, A. Babesiosis prevalence in malaria-endemic regions of Colombia. J. Vector Borne Dis. 2018, 55, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Garnham, P.C. Human babesiosis: European aspects. Trans. R. Soci. Trop. Med. Hyg. 1980, 74, 153–155. [Google Scholar] [CrossRef]
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick-Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Gray, J.S.; Hunfeld, K.P. Human babesiosis in Europe: What clinicians need to know. Infection 2013, 41, 1057–1072. [Google Scholar] [CrossRef] [PubMed]
- Martinot, M.; Zadeh, M.M.; Hansmann, Y.; Grawey, I.; Christmann, D.; Aguillon, S.; Jouglin, M.; Chauvin, A.; De Briel, D. Babesiosis in immunocompetent patients, Europe. Emerg. Infect. Dis. 2011, 17, 114–116. [Google Scholar] [CrossRef]
- Häselbarth, K.; Tenter, A.M.; Brade, V.; Krieger, G.; Hunfeld, K.P. First case of human babesiosis in Germany-Clinical presentation and molecular characterisation of the pathogen. Int. J. Med. Microbiol. 2007, 297, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Hunfeld, K.P.; Baier, M.; Krumbholz, A.; Sachse, S.; Lorenzen, T.; Kiehntopf, M.; Fricke, H.J.; Straube, E. First confirmed autochthonous case of human Babesia microti infection in Europe. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Tomassone, L.; Berriatua, E.; De Sousa, R.; Duscher, G.G.; Mihalca, A.D.; Silaghi, C.; Sprong, H.; Zintl, A. Neglected vector-borne zoonoses in Europe: Into the wild. Vet. Parasitol. 2018, 251, 17–26. [Google Scholar] [CrossRef] [PubMed]
- González, L.M.; Estrada, K.; Grande, R.; Jiménez-Jacinto, V.; Vega-Alvarado, L.; Sevilla, E.; Barrera, J.; Cuesta, I.; Zaballos, Á.; Bautista, J.M.; et al. Comparative and functional genomics of the protozoan parasite Babesia divergens highlighting the invasion and egress processes. PLoS Negl. Trop. Dis. 2019, 13, e0007680. [Google Scholar] [CrossRef]
- Gray, J.S. Identity of the causal agents of human babesiosis in Europe. Int. J. Med. Microbiol. 2006, 296 (Suppl. 40), 131–136. [Google Scholar] [CrossRef]
- Haapasalo, K.; Suomalainen, P.; Sukura, A.; Siikamaki, H.; Jokiranta, T.S. Fatal babesiosis in man, Finland, 2004. Emerg. Infect. Dis. 2010, 16, 1116–1118. [Google Scholar] [CrossRef] [PubMed]
- Mørch, K.; Holmaas, G.; Frolander, P.S.; Kristoffersen, E.K. Severe human Babesia divergens infection in Norway. Int. J. Infect. Dis. 2015, 33, 37–38. [Google Scholar] [CrossRef]
- Centeno-Lima, S.; do Rosário, V.; Parreira, R.; Maia, A.J.; Freudenthal, A.M.; Nijhof, A.M.; Jongejan, F. A fatal case of human babesiosis in Portugal: Molecular and phylogenetic analysis. Trop. Med. Int. Health 2003, 8, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Kukina, I.V.; Zelya, O.P.; Guzeeva, T.M.; Karan, L.S.; Perkovskaya, I.A.; Tymoshenko, N.I.; Guzeeva, M.V. Severe babesiosis caused by Babesia divergens in a host with intact spleen, Russia, 2018. Ticks Tick-Borne Dis. 2019, 10, 101262. [Google Scholar] [CrossRef]
- Mysterud, A.; Jore, S.; Østerås, O.; Viljugrein, H. Emergence of tick-borne diseases at northern latitudes in Europe: A comparative approach. Sci. Rep. 2017, 7, 16316. [Google Scholar] [CrossRef]
- Onyiche, T.E.; Răileanu, C.; Fischer, S.; Silaghi, C. Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature. Pathogens 2021, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, A. A historical review of Babesia spp. associated with deer in Europe: Babesia divergens/Babesia divergens-like, Babesia capreoli, Babesia venatorum, Babesia cf. odocoilei. Vet. Parasitol. 2021, 294, 109433. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Krause, P.J. Update on babesiosis. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 984568. [Google Scholar] [CrossRef]
- Gonzalez, L.M.; Rojo, S.; Gonzalez-Camacho, F.; Luque, D.; Lobo, C.A.; Montero, E. Severe babesiosis in immunocompetent man, Spain, 2011. Emerg. Infect. Dis. 2014, 20, 724–726. [Google Scholar] [CrossRef]
- Hunfeld, K.P.; Lambert, A.; Kampen, H.; Albert, S.; Epe, C.; Brade, V.; Tenter, A.M. Seroprevalence of Babesia infections in humans exposed to ticks in midwestern Germany. J. Clin. Microbiol. 2002, 40, 2431–2436. [Google Scholar] [CrossRef] [PubMed]
- Langton, C.; Gray, J.S.; Waters, P.F.; Holman, P.J. Naturally acquired babesiosis in a reindeer (Rangifer tarandus tarandus) herd in Great Britain. Parasitol. Res. 2003, 89, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Malandrin, L.; Jouglin, M.; Sun, Y.; Brisseau, N.; Chauvin, A. Redescription of Babesia capreoli (Enigk and Friedhoff, 1962) from roe deer (Capreolus capreolus): Isolation, cultivation, host specificity, molecular characterisation and differentiation from Babesia divergens. Int. J. Parasitol. 2010, 40, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Wiegmann, L.; Silaghi, C.; Obiegala, A.; Karnath, C.; Langer, S.; Ternes, K.; Kämmerling, J.; Osmann, C.; Pfeffer, M. Occurrence of Babesia species in captive reindeer (Rangifer tarandus) in Germany. Vet. Parasitol. 2015, 211, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Welc-Falęciak, R.; Pawełczyk, A.; Radkowski, M.; Pancewicz, S.A.; Zajkowska, J.; Siński, E. First report of two asymptomatic cases of human infection with Babesia microti (Franca, 1910) in Poland. Ann. Agric. Environ. Med. 2015, 22, 51–54. [Google Scholar] [CrossRef]
- Moniuszko-Malinowska, A.; Swiecicka, I.; Dunaj, J.; Zajkowska, J.; Czupryna, P.; Zambrowski, G.; Chmielewska-Badora, J.; Żukiewicz-Sobczak, W.; Swierzbinska, R.; Rutkowski, K.; et al. Infection with Babesia microti in humans with non-specific symptoms in North East Poland. Infect. Dis. 2016, 48, 537–543. [Google Scholar] [CrossRef]
- Arsuaga, M.; Gonzalez, L.M.; Lobo, C.A.; de la Calle, F.; Bautista, J.M.; Azcárate, I.G.; Puente, S.; Montero, E. First Report of Babesia microti-Caused Babesiosis in Spain. Vector Borne Zoonotic Dis. 2016, 16, 677–679. [Google Scholar] [CrossRef] [PubMed]
- Meer-Scherrer, L.; Adelson, M.; Mordechai, E.; Lottaz, B.; Tilton, R. Babesia microti infection in Europe. Curr. Microbiol. 2004, 48, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Foppa, I.M.; Krause, P.J.; Spielman, A.; Goethert, H.; Gern, L.; Brand, B.; Telford, S.R., III. Entomologic and serologic evidence of zoonotic transmission of Babesia microti, eastern Switzerland. Emerg. Infect. Dis. 2002, 8, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Pancewicz, S.; Moniuszko, A.; Bieniarz, E.; Puciło, K.; Grygorczuk, S.; Zajkowska, J.; Czupryna, P.; Kondrusik, M.; Swierzbińska-Pijanowska, R. Anti-Babesia microti antibodies in foresters highly exposed to tick bites in Poland. Scand. J. Infect. Dis. 2011, 43, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, S.; Calderini, P.; Cassini, R.; Galuppi, R.; Tampieri, M.P.; Pietrobelli, M.; Cancrini, G. Human exposure to piroplasms in central and northern Italy. Vet. Ital. 2014, 50, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, E.; Jaulhac, B.; Garcia-Bonnet, N.; Hunfeld, K.P.; Féménia, F.; Huet, D.; Goulvestre, C.; Vaillant, V.; Deffontaines, G.; Abadia-Benoist, G. Seroprevalence of seven pathogens transmitted by the Ixodes ricinus tick in forestry workers in France. Clin. Microbiol. Infect. 2016, 22, 735.e1–735.e9. [Google Scholar] [CrossRef] [PubMed]
- Svensson, J.; Hunfeld, K.P.; Persson, K.E.M. High seroprevalence of Babesia antibodies among Borrelia burgdorferi-infected humans in Sweden. Ticks Tick-Borne Dis. 2019, 10, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Lempereur, L.; Shiels, B.; Heyman, P.; Moreau, E.; Saegerman, C.; Losson, B.; Malandrin, L. A retrospective serological survey on human babesiosis in Belgium. Clin. Microbiol. Infect. 2015, 21, 96.e1–96.e7. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmsson, P.; Lövmar, M.; Krogfelt, K.A.; Nielsen, H.V.; Forsberg, P.; Lindgren, P.E. Clinical/serological outcome in humans bitten by Babesia species positive Ixodes ricinus ticks in Sweden and on the Åland Islands. Ticks Tick-Borne Dis. 2020, 11, 101455. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, L.; Yin, H.; Gubbels, M.J.; Beyer, D.; Niemann, S.; Jongejan, F.; Ahmed, J.S. Phylogeny of sheep and goat Theileria and Babesia parasites. Parasitol. Res. 2003, 91, 398–406. [Google Scholar] [CrossRef]
- Strasek-Smrdel, K.; Korva, M.; Pal, E.; Rajter, M.; Skvarc, M.; Avsic-Zupanc, T. Case of Babesia crassa-like infection, Slovenia, 2014. Emerg. Infect. Dis. 2020, 26, 1038–1040. [Google Scholar] [CrossRef] [PubMed]
- Man, S.Q.; Qiao, K.; Cui, J.; Feng, M.; Fu, Y.F.; Cheng, X.J. A case of human infection with a novel Babesia species in China. Infect. Dis. Poverty 2016, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Krause, P.J. Babesiosis in China, an emerging threat. Lancet Infect. Dis. 2015, 15, 137–139. [Google Scholar] [CrossRef]
- Chen, Z.; Li, H.; Gao, X.; Bian, A.; Yan, H.; Kong, D.; Liu, X. Human babesiosis in China: A systematic review. Parasitol. Res. 2019, 118, 1103–1112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xia, S.; Yin, S.Q.; Zhou, X.N. Emergence of babesiosis in China-Myanmar border areas. Parasit. Vectors 2015, 8, 390. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhang, L.; Yao, L.; Li, J.; Chen, H.; Ni, Q.; Pan, C.; Jin, L. Human babesiosis in southeast China: A case report. Int. J. Infect. Dis. 2018, 68, 36–38. [Google Scholar] [CrossRef]
- Bloch, E.M.; Kasubi, M.; Levin, A.; Mrango, Z.; Weaver, J.; Munoz, B.; West, S.K. Babesia microti and malaria infection in Africa: A pilot serosurvey in Kilosa district, Tanzania. Am. J. Trop. Med. Hyg. 2018, 99, 51–56. [Google Scholar] [CrossRef]
- Qi, C.; Zhou, D.; Liu, J.; Cheng, Z.; Zhang, L.; Wang, L.; Wang, Z.; Yang, D.; Wang, S.; Chai, T. Detection of Babesia divergens using molecular methods in anemic patients in Shandong Province, China. Parasitol. Res. 2011, 109, 241–245. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, S.; Yang, J.; Liu, J.; Zhang, D.; Li, Y.; Luo, J.; Guan, G.; Yin, H. Babesia divergens in human in Gansu province, China. Emerg. Microb. Infect. 2019, 8, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, S.G.; Jiang, J.F.; Wang, X.; Zhang, Y.; Wang, H.; Cao, W.C. Babesia venatorum infection in child, China. Emerg. Infect. Dis. 2014, 20, 896–897. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, S.G.; Chen, S.B.; Wang, J.Z.; Xu, B.; Zhou, H.J.; Ge, H.X.; Chen, J.H.; Hu, W. Co-infections with Babesia microti and Plasmodium parasites along the China-Myanmar border. Infect. Dis. Poverty 2013, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Piccaluga, P.P.; Poletti, G.; Martinelli, G.; Gherlinzoni, F. Babesia infection in Italy. Lancet Infect. Dis. 2004, 4, 212. [Google Scholar] [CrossRef]
- Blum, S.; Gattringer, R.; Haschke, E.; Walochnik, J.; Tschurtschenthaler, G.; Lang, F.; Oberbauer, R. The case: Hemolysis and acute renal failure. Babesiosis. Kidney Int. 2011, 80, 681–683. [Google Scholar] [CrossRef]
- Hashemi-Fesharki, R. Tick-borne diseases of sheep and goats and their related vectors in Iran. Parassitologia 1997, 39, 115–117. [Google Scholar] [PubMed]
- Chen, Y.; Yan, D.; Zhang, Y.C. Transfusion-associated babesiosis in China: A case report. Transfus. Apher. Sci. 2020, 59, 102902. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.T.; Cable, R.G.; Tonnetti, L.; Spencer, B.; Rios, J.; Leiby, D.A. Seroprevalence of Babesia microti in blood donors from Babesia-endemic areas of the northeastern United States: 2000 through 2007. Transfusion 2009, 49, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.R.; Ye, L.I.; Fan, J.W.; Li, C.; Tang, F.; Liu, W.; Ren, L.Z.; Bai, J.Y. Detection of Kobe-type and Otsu-type Babesia microti in wild rodents in China’s Yunnan province. Epidemiol. Infect. 2017, 145, 2704–2710. [Google Scholar] [CrossRef] [PubMed]
- Shiota, T.; Kurimoto, H.; Haguma, N.; Yoshida, Y. Studies on babesia first found in murine in Japan: Epidemiology, morphology and experimental infection. Zentralblatt Bakteriol. Mikrobiol. Hyg. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1984, 256, 347–355. [Google Scholar] [CrossRef]
- Wei, Q.; Tsuji, M.; Zamoto, A.; Kohsaki, M.; Matsui, T.; Shiota, T.; Telford, S.R., III; Ishihara, C. Human babesiosis in Japan: Isolation of Babesia microti-like parasites from an asymptomatic transfusion donor and from a rodent from an area where babesiosis is endemic. J. Clin. Microbiol. 2001, 39, 2178–2183. [Google Scholar] [CrossRef]
- Sayama, Y.; Zamoto-Niikura, A.; Matsumoto, C.; Saijo, M.; Ishihara, C.; Matsubayashi, K.; Nagai, T.; Satake, M. Analysis of antigen-antibody cross-reactivity among lineages and sublineages of Babesia microti parasites using human babesiosis specimens. Transfusion 2018, 58, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Zamoto-Niikura, A.; Tsuji, M.; Imaoka, K.; Kimura, M.; Morikawa, S.; Holman, P.J.; Hirata, H.; Ishihara, C. Sika deer carrying Babesia parasites closely related to B. divergens, Japan. Emerg. Infect. Dis. 2014, 20, 1398–1400. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, E.; Moustafa, M.A.M.; Lee, K.; Ching, A.L.C.; Shimozuru, M.; Sashika, M.; Nakao, R.; El-Khodery, S.A.; Tsubota, T. Utilizing attached hard ticks as pointers to the risk of infection by Babesia and Theileria species in sika deer (Cervus nippon yesoensis), in Japan. Exp. Appl. Acarol. 2020, 82, 411–429. [Google Scholar] [CrossRef]
- Hong, S.H.; Lee, S.E.; Jeong, Y.I.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Song, J.W.; Gu, S.H.; Cho, S.H.; Lee, W.J. Prevalence and molecular characterizations of Toxoplasma gondii and Babesia microti from small mammals captured in Gyeonggi and Gangwon Provinces, Republic of Korea. Vet. Parasitol. 2014, 205, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Tuvshintulga, B.; Sivakumar, T.; Battsetseg, B.; Narantsatsaral, S.O.; Enkhtaivan, B.; Battur, B.; Hayashida, K.; Okubo, K.; Ishizaki, T.; Inoue, N.; et al. The PCR detection and phylogenetic characterization of Babesia microti in questing ticks in Mongolia. Parasitol. Int. 2015, 64, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Arsuaga, M.; González, L.M.; Padial, E.S.; Dinkessa, A.W.; Sevilla, E.; Trigo, E.; Puente, S.; Gray, J.; Montero, E. Misdiagnosis of babesiosis as malaria, Equatorial Guinea, 2014. Emerg. Infect. Dis. 2018, 24, 1588–1589. [Google Scholar] [CrossRef] [PubMed]
- Fuller, A.; Manitta, J.; Marks, R.; Tencic, S.; Gordon, C.L. First reported case of imported human Babesia microti infection in Australia. Pathology 2012, 44, 580–582. [Google Scholar] [CrossRef]
- Faddy, H.M.; Rooks, K.M.; Irwin, P.J.; Viennet, E.; Paparini, A.; Seed, C.R.; Stramer, S.L.; Harley, R.J.; Chan, H.T.; Dennington, P.M.; et al. No evidence for widespread Babesia microti transmission in Australia. Transfusion 2019, 59, 2368–2374. [Google Scholar] [CrossRef]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef]
- Jefferies, R.; Ryan, U.M.; Irwin, P.J. PCR-RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet. Parasitol. 2007, 144, 20–27. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; O’Bryan, J.; Krause, P.J. The Global Emergence of Human Babesiosis. Pathogens 2021, 10, 1447. https://doi.org/10.3390/pathogens10111447
Kumar A, O’Bryan J, Krause PJ. The Global Emergence of Human Babesiosis. Pathogens. 2021; 10(11):1447. https://doi.org/10.3390/pathogens10111447
Chicago/Turabian StyleKumar, Abhinav, Jane O’Bryan, and Peter J. Krause. 2021. "The Global Emergence of Human Babesiosis" Pathogens 10, no. 11: 1447. https://doi.org/10.3390/pathogens10111447
APA StyleKumar, A., O’Bryan, J., & Krause, P. J. (2021). The Global Emergence of Human Babesiosis. Pathogens, 10(11), 1447. https://doi.org/10.3390/pathogens10111447