
Strain Characterization of Streptococcus suis Serotypes 28 and 31, Which Harbor the Resistance Genes optrA and ant(6)-Ia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statements
2.2. Bacterial Strains and Culture Conditions
2.3. Whole-Genome Sequencing Analysis
2.4. Phylogenetic Analysis
2.5. Antibiotic Susceptibility Test
2.6. Experimental Animal Infection
2.7. Statistical Analyses
3. Results
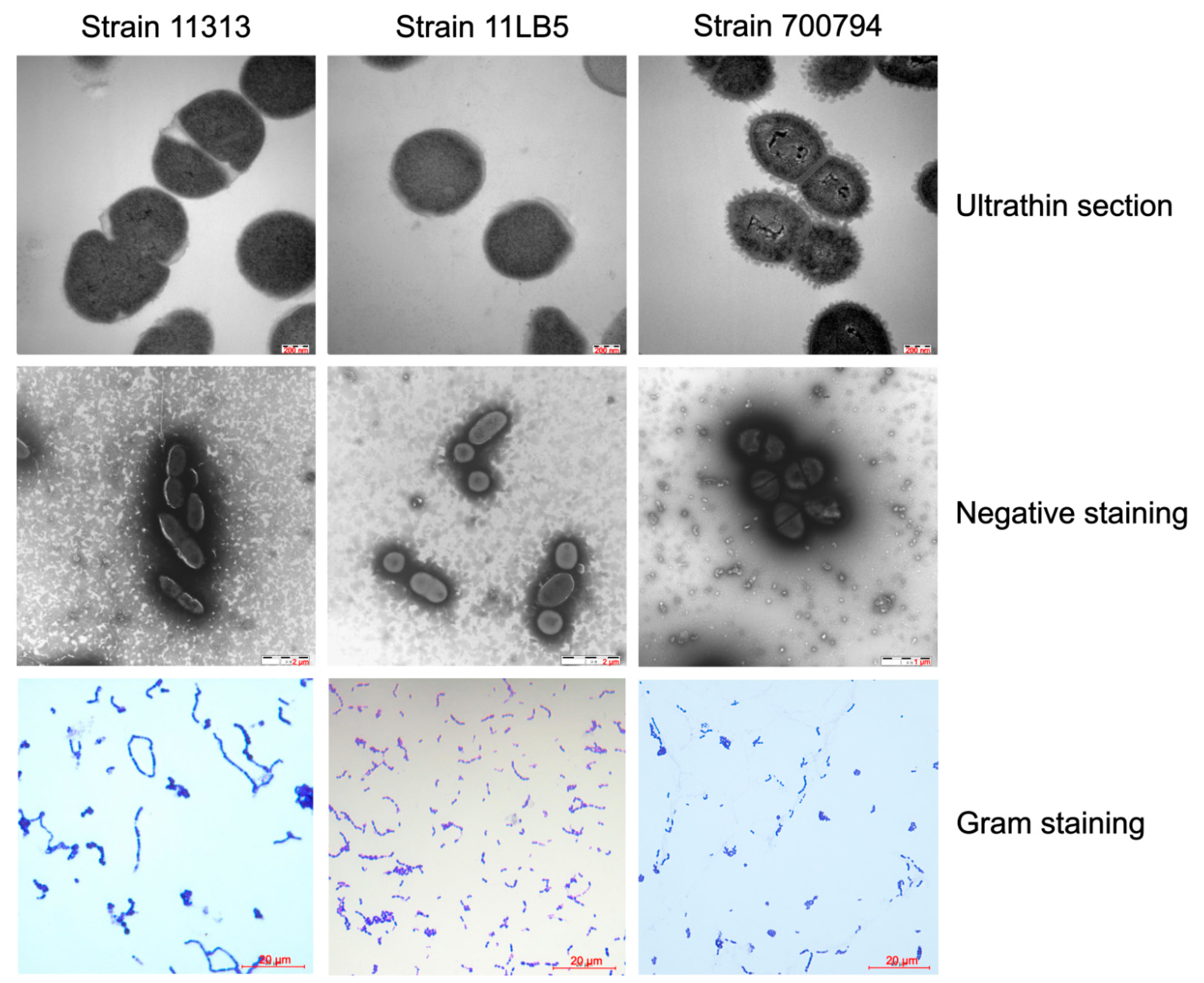
3.1. Characteristics of S. suis Strains 11313 and 11LB5
3.2. General Genomic Features of Strains 11313 and 11LB5
3.3. Structure of the Cps Gene Cluster in Strains 11313 and 11LB5
3.4. Phylogenetic Analysis of Serotype-Specific Genes of Strains 11313 and 11LB5
3.5. Antimicrobial Susceptibility Profiles
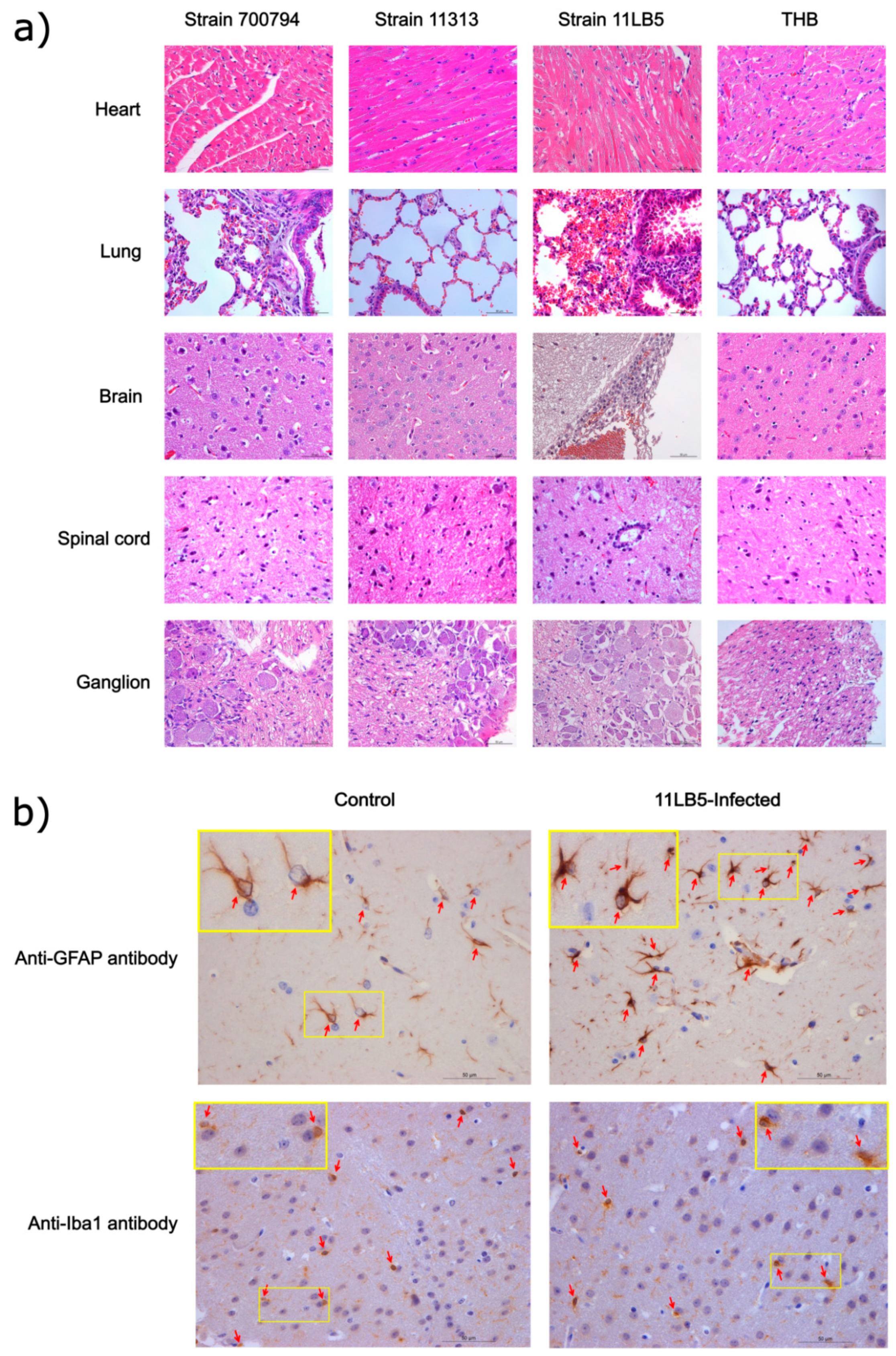
3.6. Virulence Evaluation of Strains 11313 and 11LB5 Isolates in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oh, S.I.; Jeon, A.B.; Jung, B.Y.; Byun, J.W.; Gottschalk, M.; Kim, A.; Kim, J.W.; Kim, H.Y. Capsular serotypes, virulence-associated genes and antimicrobial susceptibility of Streptococcus suis isolates from pigs in Korea. J. Vet. Med. Sci. 2017, 79, 780–787. [Google Scholar] [CrossRef] [Green Version]
- Athey, T.B.; Teatero, S.; Lacouture, S.; Takamatsu, D.; Gottschalk, M.; Fittipaldi, N. Determining Streptococcus suis serotype from short-read whole-genome sequencing data. BMC Microbiol. 2016, 16, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tien, L.H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA-DNA homology and sodA and recN phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, C.; Varaldo, P.E.; Facinelli, B. Streptococcus suis, an Emerging Drug-Resistant Animal and Human Pathogen. Front. Microbiol. 2011, 2, 235. [Google Scholar] [CrossRef] [Green Version]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Wang, S.; Gao, M.; An, T.; Liu, Y.; Jin, J.; Wang, G.; Jiang, C.; Tu, Y.; Hu, S.; Li, J.; et al. Genetic diversity and virulence of novel sequence types of Streptococcus suis from diseased and healthy pigs in China. Front. Microbiol. 2015, 6, 173. [Google Scholar] [CrossRef] [Green Version]
- Huan, H.; Jiang, L.; Tang, L.; Wang, Y.; Guo, S. Isolation and characterization of Streptococcus suis strains from swine in Jiangsu province, China. J. Appl. Microbiol. 2020, 128, 1606–1612. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, W.; Li, X.; Lu, C.; Chen, J.; Fan, W.; Huang, B. Characterization of Streptococcus suis isolates from slaughter swine. Curr. Microbiol. 2013, 66, 344–349. [Google Scholar] [CrossRef]
- Wang, S.; Liu, P.; Li, C.; Tan, Y.; Cai, X.; Zhou, D.; Jiang, Y. Isolation and characterization of 89K pathogenicity island-positive ST-7 strains of Streptococcus suis serotype 2 from healthy pigs, Northeast China. Sci. World J. 2012, 2012, 302386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Zhang, Y.; Ma, J.; Dong, W.; Zhong, X.; Pan, Z.; Yao, H. ICESsuHN105, a Novel Multiple Antibiotic Resistant ICE in Streptococcus suis Serotype 5 Strain HN105. Front. Microbiol. 2019, 10, 274. [Google Scholar] [CrossRef]
- Bojarska, A.; Janas, K.; Pejsak, Z.; Otulak-Koziel, K.; Garbaczewska, G.; Hryniewicz, W.; Sadowy, E. Diversity of serotypes and new cps loci variants among Streptococcus suis isolates from pigs in Poland and Belarus. Vet. Microbiol. 2020, 240, 108534. [Google Scholar] [CrossRef]
- Hatrongjit, R.; Kerdsin, A.; Gottschalk, M.; Takeuchi, D.; Hamada, S.; Oishi, K.; Akeda, Y. First human case report of sepsis due to infection with Streptococcus suis serotype 31 in Thailand. BMC Infect. Dis. 2015, 15, 392. [Google Scholar] [CrossRef]
- Yao, X.; Li, M.; Wang, J.; Wang, C.; Hu, D.; Zheng, F.; Pan, X.; Tan, Y.; Zhao, Y.; Hu, L.; et al. Isolation and characterization of a native avirulent strain of Streptococcus suis serotype 2: A perspective for vaccine development. Sci. Rep. 2015, 5, 9835. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okura, M.; Lachance, C.; Osaki, M.; Sekizaki, T.; Maruyama, F.; Nozawa, T.; Nakagawa, I.; Hamada, S.; Rossignol, C.; Gottschalk, M.; et al. Development of a two-step multiplex PCR assay for typing of capsular polysaccharide synthesis gene clusters of Streptococcus suis. J. Clin. Microbiol. 2014, 52, 1714–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.E.; Damman, M.; van der Velde, J.; Wagenaar, F.; Wisselink, H.J.; Stockhofe-Zurwieden, N.; Smits, M.A. Identification and characterization of the cps locus of Streptococcus suis serotype 2: The capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 1999, 67, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Fan, W.; Cai, L.; Huang, B.; Lu, C. Genetic analysis of the capsular polysaccharide synthesis locus in 15 Streptococcus suis serotypes. FEMS Microbiol. Lett. 2011, 324, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Takamatsu, D.; Maruyama, F.; Nozawa, T.; Nakagawa, I.; Osaki, M.; Sekizaki, T.; Gottschalk, M.; Kumagai, Y.; Hamada, S. Genetic analysis of capsular polysaccharide synthesis gene clusters from all serotypes of Streptococcus suis: Potential mechanisms for generation of capsular variation. Appl. Environ. Microbiol. 2013, 79, 2796–2806. [Google Scholar] [CrossRef] [Green Version]
- King, S.J.; Leigh, J.A.; Heath, P.J.; Luque, I.; Tarradas, C.; Dowson, C.G.; Whatmore, A.M. Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis: Identification of virulent clones and potential capsular serotype exchange. J. Clin. Microbiol. 2002, 40, 3671–3680. [Google Scholar] [CrossRef] [Green Version]
- Seele, J.; Nau, R.; Prajeeth, C.K.; Stangel, M.; Valentin-Weigand, P.; Seitz, M. Astrocytes Enhance Streptococcus suis-Glial Cell Interaction in Primary Astrocyte-Microglial Cell Co-Cultures. Pathogens 2016, 5, 43. [Google Scholar] [CrossRef]
- Farina, C.; Aloisi, F.; Meinl, E. Astrocytes are active players in cerebral innate immunity. Trends Immunol. 2007, 28, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Osek, J. Antimicrobial resistance mechanisms among Campylobacter. Biomed. Res. Int. 2013, 2013, 340605. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, J.; Wei, Q.; Hu, Q.; Lin, X.; Chen, M.; Ye, R.; Lv, H. Characterization of aminoglycoside resistance and virulence genes among Enterococcus spp. isolated from a hospital in China. Int. J. Environ. Res. Public Health 2015, 12, 3014–3025. [Google Scholar] [CrossRef] [PubMed]
- El-Mahdy, R.; Mostafa, A.; El-Kannishy, G. High level aminoglycoside resistant enterococci in hospital-acquired urinary tract infections in Mansoura, Egypt. Germs 2018, 8, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Seo, K.W.; Shim, J.B.; Son, S.H.; Noh, E.B.; Lee, Y.J. Molecular characterization of antimicrobial-resistant Enterococcus faecalis and Enterococcus faecium isolated from layer parent stock. Poult. Sci. 2019, 98, 5892–5899. [Google Scholar] [CrossRef]
- Dec, M.; Urban-Chmiel, R.; Stępień-Pyśniak, D.; Wernicki, A. Assessment of antibiotic susceptibility in Lactobacillus isolates from chickens. Gut Pathog. 2017, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Berbel, D.; Càmara, J.; García, E.; Tubau, F.; Guérin, F.; Giard, J.C.; Domínguez, M.; Cattoir, V.; Ardanuy, C. A novel genomic island harbouring lsa(E) and lnu(B) genes and a defective prophage in a Streptococcus pyogenes isolate resistant to lincosamide, streptogramin A and pleuromutilin antibiotics. Int. J. Antimicrob. Agents 2019, 54, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, C.; Magi, G.; Mingoia, M.; Bagnarelli, P.; Ripa, S.; Varaldo, P.E.; Facinelli, B. Characterization of a Streptococcus suis tet(O/W/32/O)-carrying element transferable to major streptococcal pathogens. Antimicrob. Agents Chemother. 2012, 56, 4697–4702. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Sun, J.; Wu, Y.; Chen, L.; Duan, D.; Lv, X.; Wang, L. Identification and pathogenicity of an XDR Streptococcus suis isolate that harbours the phenicol-oxazolidinone resistance genes optrA and cfr, and the bacitracin resistance locus bcrABDR. Int. J. Antimicrob. Agents 2019, 54, 43–48. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, R.; Li, D.; Feßler, A.T.; Wu, C.; Schwarz, S.; Wang, Y. Distribution of optrA and cfr in florfenicol-resistant Staphylococcus sciuri of pig origin. Vet. Microbiol. 2017, 210, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Seo, K.W.; Son, S.H.; Noh, E.B.; Lee, Y.J. Genetic characterization of high-level aminoglycoside-resistant Enterococcus faecalis and Enterococcus faecium isolated from retail chicken meat. Poult. Sci. 2019, 98, 5981–5988. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Frimodt-Møller, N.; Aarestrup, F.M. Presence of erm gene classes in gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef]
- Tan, M.F.; Tan, J.; Zeng, Y.B.; Li, H.Q.; Yang, Q.; Zhou, R. Antimicrobial resistance phenotypes and genotypes of Streptococcus suis isolated from clinically healthy pigs from 2017 to 2019 in Jiangxi Province, China. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Serotype | wzy Genes | Accession No. |
|---|---|---|---|
| 11313 | 28 | cps28L | NA647439 |
| 11LB5 | 31 | cps31L | NA647982 |
| 2651 | 1/2 | cps1/2I | KC537384 |
| 5428 | 1 | cps1I | JX986790 |
| R735 | 2 | cps2I | KC537364 |
| 4961 | 3 | cps3L | KC537365 |
| 6407 | 4 | cps4K | KC537366 |
| 11538 | 5 | cps5L | KC537367 |
| 2524 | 6 | cps6I | KC537368 |
| 8074 | 7 | cps7L | KC537369 |
| 14636 | 8 | cps8K | JX986797 |
| 22083 | 9 | cps9J | KC537370 |
| 4417 | 10 | cps10M | JX986799 |
| 12814 | 11 | cps11N | KC537371 |
| 8830 | 12 | cps12J | KC537372 |
| 10581 | 13 | cps13L | JX961643 |
| 13730 | 14 | cps14H | JX986804 |
| NCTC 10446 | 15 | cps15K | JX961644 |
| 2726 | 16 | cps16I | KC537373 |
| 93A | 17 | cps17O | KC537374 |
| NT77 | 18 | cps18N | KC537375 |
| 42A | 19 | cps19L | KC537376 |
| 14A | 21 | cps21P | KC537378 |
| 89-2479 | 23 | cps23J | JX986802 |
| 88-5299A | 24 | cps24M | KC537380 |
| 89-3576-3 | 25 | cps25M | JX986803 |
| 89-5259 | 27 | cps27K | JX961652 |
| 89-590 | 28 | cps28L | JX961653 |
| 92-1191 | 29 | cps29L | JX961654 |
| 92-1400 | 30 | cps30I | KC537382 |
| 92-4172 | 31 | cps31L | JX961656 |
| Strain | Serotype | MLST Type | Accession No. | Size (bp) | GC (%) | Genes | Proteins | Antibiotic Resistance Genes | Virulence Phenotype |
|---|---|---|---|---|---|---|---|---|---|
| 11313 | 28 | ST422 | NA647439 | 2,261,105 | 41.45 | 1860 | 1802 | erm(B),optrA, tet(l), tet(o), ant(6)-Ia | gdh+/mrp-/sly-/epf- |
| 11LB5 | 31 | ST421 | NA647982 | 2,202,712 | 41.42 | 2078 | 1998 | aac(6′)-aph(2′’), ant(6)-Ia, erm(B), tet(40), tet(o/w/32/o) | gdh+/mrp-/sly-/epf- |
| Category | Antibiotic | K-B(mm)/Antimicrobial Susceptibility * | MIC (mg/L) /Antimicrobial Susceptibility * | ||||
|---|---|---|---|---|---|---|---|
| Strain 11313 | Strain 11LB5 | Strain 700794 | Strain 11313 | Strain 11LB5 | Strain 700794 | ||
| Penicillin | Ampicillin | 15/S | 24/S | 20/S | <8/S | <4/S | <4/S |
| Cephalosporin | Ceftazidime | 26/S | 21/S | 26/S | <4/S | <4/S | <4/S |
| Carbapenem | Meropenem | 38/S | 50/S | 40/S | <1/S | <1/S | <1/S |
| Cephamycin | Cefoxitin | 25/S | 34/S | 30/S | <8/S | <4/S | <4/S |
| Aminoglycoside | Streptomycin | 10/R | 6/R | 10/R | >512/R | >512/R | >512/R |
| Tetracycline | Tetracycline | 6/R | 6/R | 20/S | >512/R | >512/R | <8/S |
| Phenicol | Chloramphenicol | 16/I | 40/S | 21/S | 16/I | <4/S | <8/S |
| Folate pathway | Trimethoprim | 10/R | 28/S | 20/S | >32/R | <0.5/S | <1/S |
| Nitrofuran | Nitrofurantoin | 28/S | 32/S | 30/S | <16/S | 8/S | 8/S |
| Fluoroquinolone | Ciprofloxacin | 6/R | 22/S | 30/S | >216/R | <1/S | <0.5/S |
| Macrolide | Spiramycin | 6/R | 6/R | 23/S | >512/R | >512/R | 8/S |
| Fosfomycin | Fosfomycin | 25/S | 32/S | 26/S | <64/S | <32/S | <64/S |
| Peptide antibiotics | Vancomycin | 15/S | 25/S | 20/S | 8/S | 4/S | 4/S |
| oxazolidinones | linezolid | 32/S | 34/S | 44/S | <0.5/S | <0.5/S | 0.25/S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhang, D.; Jiang, C.; He, H.; Cui, C.; Duan, W.; Hu, S.; Wang, J.; Cai, X. Strain Characterization of Streptococcus suis Serotypes 28 and 31, Which Harbor the Resistance Genes optrA and ant(6)-Ia. Pathogens 2021, 10, 213. https://doi.org/10.3390/pathogens10020213
Wang S, Zhang D, Jiang C, He H, Cui C, Duan W, Hu S, Wang J, Cai X. Strain Characterization of Streptococcus suis Serotypes 28 and 31, Which Harbor the Resistance Genes optrA and ant(6)-Ia. Pathogens. 2021; 10(2):213. https://doi.org/10.3390/pathogens10020213
Chicago/Turabian StyleWang, Shujie, Defu Zhang, Chenggang Jiang, Haijuan He, Chenchen Cui, Weitong Duan, Shouping Hu, Jun Wang, and Xuehui Cai. 2021. "Strain Characterization of Streptococcus suis Serotypes 28 and 31, Which Harbor the Resistance Genes optrA and ant(6)-Ia" Pathogens 10, no. 2: 213. https://doi.org/10.3390/pathogens10020213
APA StyleWang, S., Zhang, D., Jiang, C., He, H., Cui, C., Duan, W., Hu, S., Wang, J., & Cai, X. (2021). Strain Characterization of Streptococcus suis Serotypes 28 and 31, Which Harbor the Resistance Genes optrA and ant(6)-Ia. Pathogens, 10(2), 213. https://doi.org/10.3390/pathogens10020213