
Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
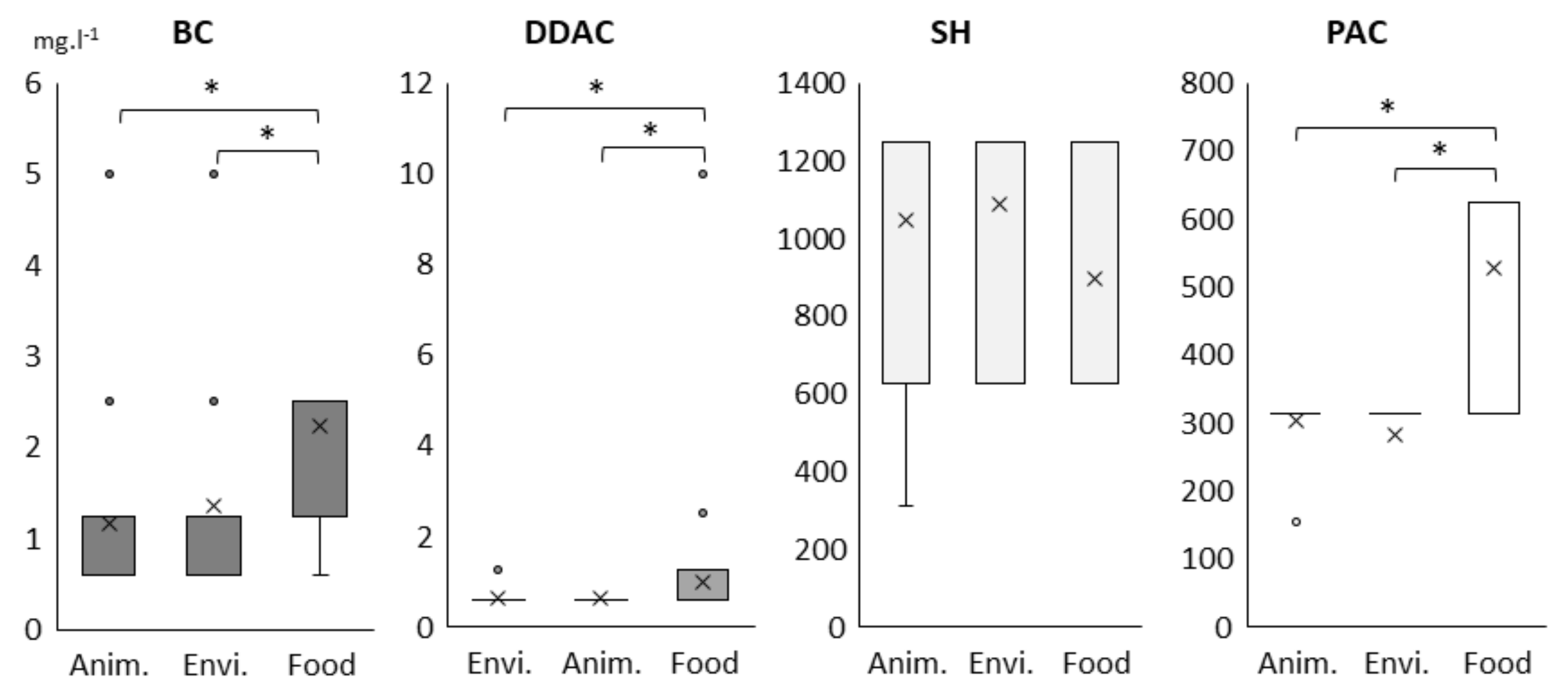
2.1. Biocides and Antibiotic Susceptibility Profiles of the 205 Lm Strains
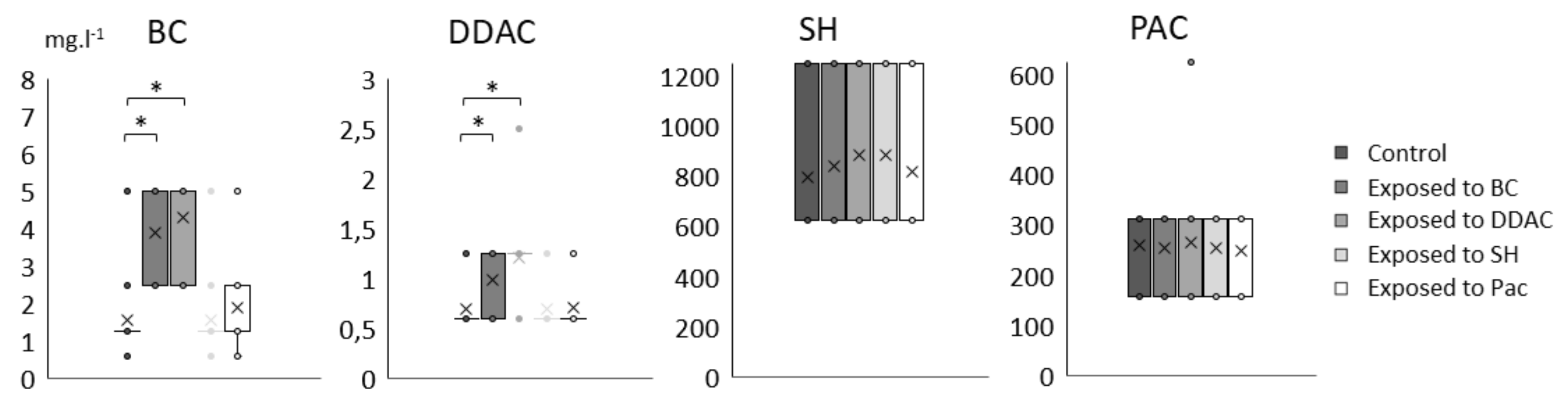
2.2. Adaptation to Biocide after Repeated Exposures to BC, DDAC, SH and PAC
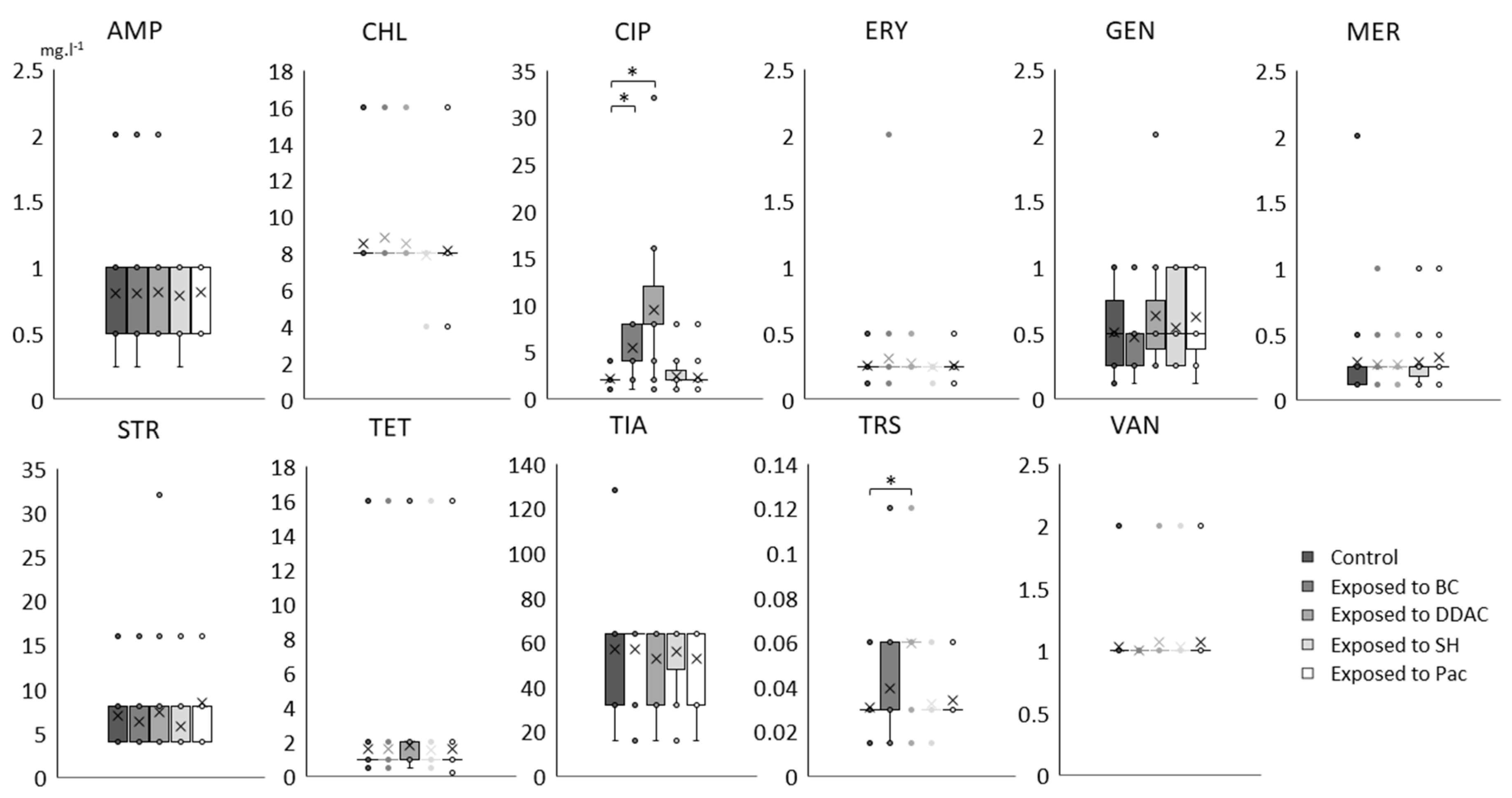
2.3. Effects of Adaptation to Biocides on Antibiotic MIC Values
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Minimal Inhibitory Concentrations (MIC) of Antibiotics and Biocides
4.3. Adaptation Experiments to Four Biocides
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. Available online: https://efsa.onlinelibrary.wiley.com/doi/epdf/10.2903/j.efsa.2019.5926 (accessed on 19 June 2020).
- De Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.A.; Kirk, M.D.; Havelaar, A.H.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; Cabo, M.L. Current Knowledge on Listeria monocytogenes Biofilms in Food-Related Environments: Incidence, Resistance to Biocides, Ecology and Biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide-Tolerant Listeria monocytogenes Isolates from German Food Production Plants Do Not Show Cross-Resistance to Clinically Relevant Antibiotics. Appl. Environ. Microbiol. 2019, 85, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivant, A.-L.; Garmyn, D.; Piveteau, P. Listeria monocytogenes, a down-to-earth pathogen. Front. Cell. Infect. Microbiol. 2013, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Félix, B.; Sevellec, Y.; Palma, F.; Felten, A.; Radomski, N.; Mallet, L.; Blanchard, Y.; Leroux, A.; Soumet, C.; Bridier, A.; et al. A European-Wide Dataset to Decipher Adaptation Mechanisms of Listeria Monocytogenes to Diverse Ecological Niches. Sci. Data 2020, in press. [Google Scholar]
- European Commission Regulation (EC) No 852/2004 of the European Parliament and of the Council of 29 April 2004 on the Hygiene of Foodstuffs. Off. J. Eur. Comm. 2004. L139/1. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:32004R0852 (accessed on 17 February 2021).
- European Commission Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 Concerning the Making Available on the Market and Use of Biocidal Products. Off. J. Eur. Comm. 2012. L 167/1. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32012R0528 (accessed on 17 February 2021).
- Davin-Regli, A.; Pagès, J.M. Cross-Resistance between Biocides and Antimicrobials: An Emerging Question. Rev. Sci. Tech. Int. Off. Epizoot. 2012, 31, 89–104. [Google Scholar] [CrossRef]
- Forbes, S.; Knight, C.G.; Cowley, N.L.; Amézquita, A.; McClure, P.; Humphreys, G.; McBain, A.J. Variable Effects of Exposure to Formulated Microbicides on Antibiotic Susceptibility in Firmicutes and Proteobacteria. Appl. Environ. Microbiol. 2016, 82, 3591–3598. [Google Scholar] [CrossRef] [Green Version]
- Maillard, J.-Y. Bacterial Target Sites for Biocide Action. J. Appl. Microbiol. 2002, 92, 16S–27S. [Google Scholar] [CrossRef] [PubMed]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial Resistance in Listeria Species. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 6, 237–259. [Google Scholar] [CrossRef]
- Gerba, C.P. Quaternary Ammonium Biocides: Efficacy in Application. Appl. Environ. Microbiol. 2014, 81, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes Strains Selected on Ciprofloxacin or the Disinfectant Benzalkonium Chloride Exhibit Reduced Susceptibility to Ciprofloxacin, Gentamicin, Benzalkonium Chloride, and Other Toxic Compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Jiang, X.; Zhang, Y.; Ji, S.; Gao, W.; Shi, L. Effect of Benzalkonium Chloride Adaptation on Sensitivity to Antimicrobial Agents and Tolerance to Environmental Stresses in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mereghetti, L.; Quentin, R.; Der Mee, N.M.-V.; Audurier, A. Low Sensitivity of Listeria monocytogenes to Quaternary Ammonium Compounds. Appl. Environ. Microbiol. 2000, 66, 5083–5086. [Google Scholar] [CrossRef] [Green Version]
- Soumet, C.; Méheust, D.; Pissavin, C.; Legrandois, P.; Frémaux, B.; Feurer, C.; Le Roux, A.; Denis, M.; Maris, P. Reduced susceptibilities to biocides and resistance to antibiotics in food-associated bacteria following exposure to quaternary ammonium compounds. J. Appl. Microbiol. 2016, 121, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Bansal, M.; Nannapaneni, R.; Sharma, C.S.; Kiess, A. Listeria monocytogenes Response to Sublethal Chlorine Induced Oxidative Stress on Homologous and Heterologous Stress Adaptation. Front. Microbiol. 2018, 9, 2050. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; García-Fernández, C.; Alonso-Calleja, C.; Capita, R. Susceptibility of Listeria monocytogenes planktonic cultures and biofilms to sodium hypochlorite and benzalkonium chloride. Food Microbiol. 2019, 82, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.-J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Morrissey, I.; Oggioni, M.R.; Knight, D.R.; Curiao, T.; Coque, T.; Kalkanci, A.; Martinez, J.L.; The BIOHYPO Consortium. Evaluation of Epidemiological Cut-Off Values Indicates that Biocide Resistant Subpopulations Are Uncommon in Natural Isolates of Clinically-Relevant Microorganisms. PLoS ONE 2014, 9, e86669. [Google Scholar] [CrossRef] [Green Version]
- Gadea, R.; Fuentes, M.; Ángel, F.; Pulido, R.P.; Gálvez, A.; Ortega, E. Effects of exposure to quaternary-ammonium-based biocides on antimicrobial susceptibility and tolerance to physical stresses in bacteria from organic foods. Food Microbiol. 2017, 63, 58–71. [Google Scholar] [CrossRef]
- Kampf, G. Antibiotic Resistance Can Be Enhanced in Gram-Positive Species by Some Biocidal Agents Used for Disinfection. Antibiotics 2019, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of Antibiotic Resistance inListeria monocytogenesIsolated from Food Products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [Green Version]
- Maertens, H.; Demeyere, K.; De Reu, K.; Dewulf, J.; Vanhauteghem, D.; Van Coillie, E.; Meyer, E. Effect of subinhibitory exposure to quaternary ammonium compounds on the ciprofloxacin susceptibility of Escherichia coli strains in animal husbandry. BMC Microbiol. 2020, 20, 155. [Google Scholar] [CrossRef]
- Romanova, N.A.; Wolffs, P.F.G.; Brovko, L.Y.; Griffiths, M.W. Role of Efflux Pumps in Adaptation and Resistance of Listeria monocytogenes to Benzalkonium Chloride. Appl. Environ. Microbiol. 2006, 72, 3498–3503. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Yu, T.; Xu, Y.; Wang, H.; Korkeala, H.; Shi, L. MdrL, a major facilitator superfamily efflux pump of Listeria monocytogenes involved in tolerance to benzalkonium chloride. Appl. Microbiol. Biotechnol. 2018, 103, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Godreuil, S.; Galimand, M.; Gerbaud, G.; Jacquet, C.; Courvalin, P. Efflux Pump Lde Is Associated with Fluoroquinolone Resistance in Listeria monocytogenes. Antimicrob. Agents Chemother. 2003, 47, 704–708. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Zhou, L.; Gao, N.; Wang, Y.; Wang, D.; Zhang, Z.; Chen, M.; Su, Y.; Li, L.; Yan, H.; et al. Expression of efflux pump geneldein ciprofloxacin-resistant foodborne isolates of Listeria monocytogenes. Microbiol. Immunol. 2012, 56, 843–846. [Google Scholar] [CrossRef] [PubMed]
- To, M.S.; Favrin, S.; Romanova, N.; Griffiths, M.W. Postadaptational Resistance to Benzalkonium Chloride and Subsequent Physicochemical Modifications of Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 5258–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacevic, J.; Ziegler, J.; Wałecka-Zacharska, E.; Reimer, A.; Kitts, D.D.; Gilmour, M.W. Tolerance of Listeria monocytogenes to Quaternary Ammonium Sanitizers Is Mediated by a Novel Efflux Pump Encoded by emrE. Appl. Environ. Microbiol. 2015, 82, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Komora, N.; Bruschi, C.; Magalhães, R.; Ferreira, V.B.; Teixeira, P. Survival of Listeria monocytogenes with different antibiotic resistance patterns to food-associated stresses. Int. J. Food Microbiol. 2017, 245, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Suárez, J.V.; Ortiz, S.; López-Alonso, V. Potential Impact of the Resistance to Quaternary Ammonium Disinfectants on the Persistence of Listeria monocytogenes in Food Processing Environments. Front. Microbiol. 2016, 7, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0.008 | 0.016 | 0.032 | 0.064 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | |
| AMP | 25 | 128 | 52 | ||||||||||||
| CHL | 1 | 192 | 12 | ||||||||||||
| CIP | 4 | 58 | 122 | 19 | 2 | ||||||||||
| ERY | 22 | 178 | 5 | ||||||||||||
| GEN | 1 | 12 | 71 | 76 | 45 | ||||||||||
| MER | 134 | 70 | 1 | ||||||||||||
| STR | 4 | 98 | 80 | 23 | |||||||||||
| TET | 1 | 8 | 161 | 34 | 1 | ||||||||||
| TIA | 1 | 5 | 53 | 144 | 2 | ||||||||||
| TRS | 20 | 176 | 9 | ||||||||||||
| VAN | 199 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guérin, A.; Bridier, A.; Le Grandois, P.; Sévellec, Y.; Palma, F.; Félix, B.; LISTADAPT Study Group; Roussel, S.; Soumet, C. Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes. Pathogens 2021, 10, 220. https://doi.org/10.3390/pathogens10020220
Guérin A, Bridier A, Le Grandois P, Sévellec Y, Palma F, Félix B, LISTADAPT Study Group, Roussel S, Soumet C. Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes. Pathogens. 2021; 10(2):220. https://doi.org/10.3390/pathogens10020220
Chicago/Turabian StyleGuérin, Alizée, Arnaud Bridier, Patricia Le Grandois, Yann Sévellec, Federica Palma, Benjamin Félix, LISTADAPT Study Group, Sophie Roussel, and Christophe Soumet. 2021. "Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes" Pathogens 10, no. 2: 220. https://doi.org/10.3390/pathogens10020220
APA StyleGuérin, A., Bridier, A., Le Grandois, P., Sévellec, Y., Palma, F., Félix, B., LISTADAPT Study Group, Roussel, S., & Soumet, C. (2021). Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes. Pathogens, 10(2), 220. https://doi.org/10.3390/pathogens10020220